Article
솜대 조림지와 확산지의 바이오매스, 임상, 토양의 양분 특성
곽유식1, 백경원1, 최병길1, 하지석1, 배은지2, 김춘식1,*
Nutrient Characteristics of Biomass, Forest Floor, and Soil between Plantation and Expansion Sites of Phyllostachys nigra var. henonis
You Sig Kwak1, Gyeongwon Baek1, Byeonggil Choi1, Jiseok Ha1, Eun Ji Bae2, Choonsig Kim1,*
1Department of Forest Resources, Gyeongsang National University, Jinju 52725, Korea
2Forest Biomaterials Research Center, National Institute of Forest Science, Jinju 52817, Korea
© Copyright 2021 Korean Society of Forest Science. This is an Open-Access article distributed under the terms of the
Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits
unrestricted non-commercial use, distribution, and reproduction in any
medium, provided the original work is properly cited.
Received: Jan 05, 2021; Revised: Mar 01, 2021; Accepted: Mar 02, 2021
Published Online: Mar 31, 2021
요약
본 연구는 대나무 확산에 있어서 토양 성질이 미치는 영향을 조사하기 위해 매년 시비가 실시된 솜대 조림지와 미시비 확산지를 대상으로 바이오매스 부위별 양분 농도와 임상 및 토양 30 cm 깊이의 양분 특성을 비교하였다. 솜대 조림지의 잎, 가지, 줄기 내 질소와 인 농도는 확산지에 비해 유의적으로(P < 0.05) 높았으나, 임상의 양분 농도는 조림지와 확산지 사이에 유의적인 차이가 없었다. 토양 0~30 cm 깊이의 평균 유기탄소는 조림지가 30.80 mg g−1으로 확산지 15.64 mg g−1에 비해 유의적으로 높았다. 평균 전질소 농도의 경우 조림지 2.47 mg g−1, 확산지 1.24 mg g−1, 인은 조림지 10.27 mg kg−1, 확산지 5.61 mg kg−1, 포타슘은 조림지 0.27 cmolc kg−1, 확산지 0.16 cmolc kg−1로 조림지가 확산지에 비해 유의적으로 높았다. 본 연구 결과에 따르면 토양 양분 상태는 솜대의 인접 산림지역으로 확산에 큰 영향을 끼치지 않는 것으로 나타났다.
Abstract
In this study, the relationships between bamboo expansion and the nutrient characteristics of bamboo biomass, the forest floor, and mineral soil (at 30-cm depth) were determined in unfertilized expansion sites and fertilized plantations of Phyllostachysnigra var. henonis in Jinju, Gyeongsangnam-do. Nitrogen and phosphorus concentrations in bamboo components (culm, branches, and foliage) were significantly higher in the plantation site than those in the expansion site (P < 0.05). However, the nutrient concentration of the forest floor did not differ significantly between the plantation and expansion sites. Mean organic carbon concentration at 0–30-cm soil depth was significantly higher in the plantation site (30.80 g kg−1) than that in the expansion site (15.64 g kg−1). In addition, total nitrogen, phosphorus, and exchangeable K+ at 0–30-cm soil depth were significantly higher in the plantation site than those in the expansion site. These results indicate that bamboo can spread to areas with low-nutrient concentrations in adjacent forests.
Keywords: bamboo; bamboo spreading; bamboo nutrient; forest soils; nutrition distribution
서 론
대나무는 탄소흡수, 연료, 가구 재료, 식용, 황폐지의 토양 개량 등 다양한 기능이 제공되는 중요한 목본성 초본으로 알려져 있다(Christanty et al., 1996; Nath et al., 2015; Yuen et al., 2017). 그러나 최근 방치된 대나무 임분의 산림지역 확산은 일본, 중국 등 세계 여러 지역에서 산림자원 관리적인 면에서 문제로 대두되고 있다(Okutomi et al., 1996; Griscom and Ashton, 2003; Suzuki and Nakagoshi, 2008; Inoue et al., 2012; Bai et al., 2106a, b; Wang et al., 2016; Shiau and Chiu, 2017; Song et al., 2017). 대나무 확산지의 산림생태계는 생물 다양성 감소, 토양 수분 및 성질 변화, 토양 미생물 다양성 감소 등이 보고되고 있으며, 대나무로부터 방출되는 타감물질은 하층 식생의 생육을 억제하거나 고사를 초래하기도 한다(Suzuki and Naka goshi, 2008; Inoue et al., 2012; Wang et al., 2016).
대나무 확산에 대한 일련의 과정은, 1) 인접 산림지역으로 지하경(rhizome)의 생장 및 확산, 2) 산림지역에 지하경으로부터 죽순 발생, 3) 발순된 죽순은 임목 생장 기간 동안 양호한 광 조건을 보이는 상층 임관에 도달, 4) 새롭게 생육한 대나무가 수관부(canopy layer)를 점유하여 장기간 생존하는 전략을 가지고 있다(Yoo et al., 2017). 최근 조사된 우리나라 대나무 분포면적은 22,067 ha로 알려져 있으며, 방치된 대나무 임분의 증가에 따라 확산이 상당히 진행되고 있는 것으로 보고되고 있다(Yoo et al., 2017). 국내 대나무 확산 유형은 확산형, 관리형, 혼합형, 쇠퇴형, 분리형으로 구분되며, 경상남도 사천시와 거제시 대나무 혼효임분이나 맹종죽 임분의 30년(1980년~2010년) 동안 확산 속도는 0.06~0.08 ha yr−1, 확산 거리는 0.9~1.1 m yr−1 정도였다(Yoo et al., 2017). 국외 사례로 일본 가고시마현 대나무의 임분은 15년 동안 2.09 m yr−1 확산하였으며(Okutomi et al., 1996), 중국의 맹종죽 임분은 인접한 전나무 임분으로 1.28 m yr−1, 상록활엽수 임분으로 1.04 m yr−1 확산하였다. 대나무 확산은 산림 토양환경에 상당한 영향을 끼치는 것으로 알려져 있다(Umemura and Takenaka, 2015; Shiau and Chiu, 2017; Zaninovich et al., 2017). 예를 들면, 아르헨티나 북부 산림토양은 대나무 확산으로 인한 낙엽·낙지량 및 낙엽분해율 감소로 인하여 표토 5 cm 깊이의 토양 유기탄소량이 50% 정도 감소하였고(Zaninovich et al., 2017), 대만 삼나무 조림지의 경우도 맹종죽의 확산으로 인하여 유기탄소 및 질소 총량이 감소하여 토양의 질(soil quality)이 저하된 것으로 보고되고 있다(Shiau and Chiu, 2017). 한편, 일본에서 조사된 대나무 확산 임분지의 토양 pH는 4.3~5.1로 인접한 편백 임분의 토양 pH 3.4~4.5보다 유의적으로 높아 토양 산성화를 완화하는 효과가 있었다(Umemura and Takenaka, 2015).
대나무의 산림지역 확산은 방치되거나 관리되지 않는 임분에서 주로 발생하는 것으로 알려져 있으나(Okutomi et al., 1996; Umemura and Takenaka, 2015; Yoo et al., 2017), 산림관리가 실시되는 지역에서도 대나무 확산이 관찰되고 있으며(Yoo et al., 2017), 이들 지역의 대나무 확산에 대한 정보는 거의 없는 편이다. 한편, 대나무의 산림 내 확산 기작을 이해하기 위해서는 지상부와 지하부의 양분 분포를 조사하는 것이 중요한 것으로 알려져 있으나(Umemura and Takenaka, 2015), 국내에서는 아직까지 이들에 관한 연구가 수행된 바 없다. 본 연구는 국내 대나무 분포 면적 22,968 ha 중 6,175 ha가 분포하는 것으로 알려진(National Institute of Forest Science, 2016) 솜대를 대상으로 지속적인 산림 관리가 실시된 조림지와 솜대 확산지의 바이오매스 및 지하부 토양 내 양분 특성을 비교하여 대나무 확산과 토양 성질과의 관련성을 이해하기 위한 목적으로 수행하였다. 본 연구에서는 솜대가 확산되고 있는 지역의 바이오매스, 임상, 토양의 양분 함량은 지속적으로 산림관리가 실시된 조림지와 차이가 있을 것이라는 가설을 검정하고자 하였다.
재료 및 방법
1. 조사지 개황
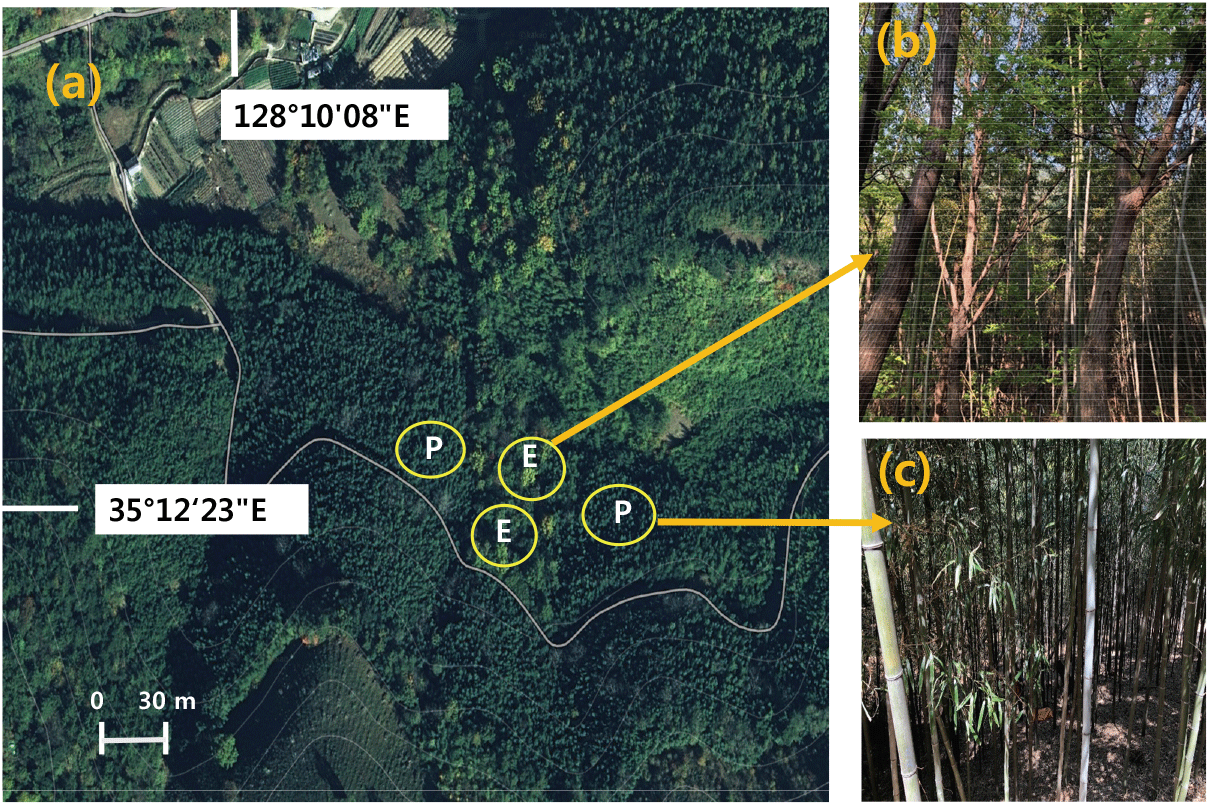
본 연구 조사지는 경상남도 진주시 금산면 장사리 국립산림과학원 산림바이오소재연구소의 월아연구시험림 내 식재된 솜대 조림지를 대상으로 하였다(Figure 1). 이 지역은 1994년에서 1995년 사이에 왕대와 솜대를 각 5 ha씩 총 10 ha를 조림한 지역으로(Jung et al., 2017), 조사 당시는 조성 후 약 25년이 경과한 임분이다. 조림지 관리면에서 이 지역은 2000년 이후 지속적인 산림관리가 실시되어 매년 2차에 걸쳐 년 580 kg ha−1의 복합비료(N-P-K:21-17-17)를 3~4월에 290 kg ha−1(N: 60 kg, P2O5: 49 kg, K2O: 49 kg)과 7~8월에 290 kg ha−1을 시비하고 있다. 조사지의 해발고는 110 m~120 m 범위였고 산림토양형은 퇴적암인 사암이나 혈암에서 유래된 암적갈색건조산림토양형(DRb1)과 암적갈색약건산림토양형(DRb2)이 분포하였다. 조사구는 2020년 4월 입지환경 특성 및 밀도를 고려하여 조림지와 확산지를 대표하는 각 2개의 10 m × 10 m 조사구를 설치하였다(Figure 1). 조사지로 선정된 확산지는 1995년 솜대 조림지 조성 시 사유지로서 솜대가 전혀 식재되지 않았던 지역으로 조림지 조성 약 25년 경과 후 인접 조림지로부터 유입된 대나무 확산 정도를 추정할 수 있는 지역이었다. 조림지에는 목본성 수종이 전혀 분포하지 않았지만 확산지에는 밤나무(Castanea crenata S. et Z.), 아까시나무(Robinia pseudoacacia L.), 은사시나무(Populus tomentiglandulosa T.B. Lee), 상수리나무(Quercus acutissima Carruth), 졸참나무(Q. serrata Murray) 등이 생육하고 있었으며, 수종식별이 어려운 고사목은 평균 400본 ha−1이었다(Table 1).
Figure 1.
Location (a) of expansion sites (E, b) and plantation (P, c) in Phyllostachys nigra var. henonis.
Download Original Figure
Table 1.
General characteristics of the study site.
|
Treatment |
Latitude (N) |
Longitude (E) |
Aspect |
Elevation (m a.s.l.) |
Density (culm ha−1) |
DBH (cm) |
Basal area (m2 ha−1) |
|
Plantation 1 |
35°12′24″ |
128°10′14″ |
E |
110 |
12,000 |
4.58 |
|
|
Plantation 2 |
35°12′22″ |
128°10′16″ |
NW |
120 |
11,400 |
5.10 |
|
|
Expansion 1 |
35°12′23″ |
128°10′15″ |
NE |
110 |
-
– bamboo: 6,600
-
– living tree: 500
-
– dead tree: 600
|
-
– bamboo: 5.69
-
– living tree: 20.7
-
– dead tree: 15.3
|
-
– living tree: 22.6
-
– dead tree: 11.4
|
|
Expansion 2 |
35°12′23″ |
128°10′15″ |
NE |
120 |
-
– bamboo: 6,440
-
– living tree: 800
-
– dead tree: 200
|
-
– bamboo: 5.67
-
– living tree: 17.6
-
– dead tree: 9.8
|
-
– living tree: 22.8
-
– dead tree: 1.7
|
Download Excel Table
2. 시료채취 및 분석방법
1) 바이오매스 및 임상 유기물 양분 분석
조림지와 확산지의 바이오매스 양분 분석 시료는 각 조사구에 분포하는 솜대를 직경급이 고르게 분포하도록 하여 2020년 5월 1일 조림지 6본, 확산지 6본 등 총 12본을 톱을 이용하여 지상부 10 cm 높이에서 벌채하였다. 벌채된 솜대는 수관 폭을 줄자를 이용하여 측정한 후, 줄기, 가지, 잎을 분리하고 바이오매스 시료 일부를 비닐주머니에 밀봉하여 실험실로 운반하고 65°C 온풍건조기에서 항량에 도달할 때까지 건조하였다. 또한 지하부 뿌리 양분 농도는 각 조사구 내 가로 50 cm, 세로 50 cm, 깊이 50 cm의 토양 단면을 2개씩 제작하고 400 cm3 원형 강철 토양채취기(내직경: 7.04 cm; 높이: 10.4 cm)를 이용하여 0~10 cm, 10~20 cm, 20~30 cm 간격으로 토양을 채취하여 실험실로 운반하였다. 채취된 토양 내 뿌리는 육안으로 식별하고 분리하여 수돗물로 세척 후 65°C 건조기에서 항량에 도달할 때까지 건조하였다.
건조된 바이오매스 부위별 시료는 분쇄 후 0.425 mm 체(US standard No. 40)로 양분 분석용 시료를 조제하였다. 탄소와 질소는 원소분석기(Thermo Fisher Scientific Flash 2000, Italy), 인, 포타슘, 칼슘, 마그네슘은 분쇄된 시료 0.5 g을 470°C 전기로에서 4시간 이상 회화한 후 염산과 질산으로 추출된 용액을(Kalra and Maynard, 1991) 대상으로 ICP (Perkin Elmer Optima 5300, USA)를 이용하여 분석하였다. 임상(forest floor)의 시료는 30 cm × 30 cm 강철프레임을 이용하여 토양층 상부까지 채취하고 실험실로 운반한 후 65°C 온풍건조기에 넣고 항량에 도달할 때까지 건조하였다. 건조된 임상시료는 분쇄하여 바이오매스 분석과 동일한 방법으로 양분 분석을 실시하였다.
2) 토양 물리·화학적 성질 분석
각 조사구 내 토양 분석 시료는 임상 시료가 채취된 지점의 토양층을 대상으로 400 cm3 원형 강철 토양 채취기를 이용하여 0~30 cm 깊이의 시료를 10 cm 간격으로 각 2개씩 채취하고 지퍼 백에 넣고 밀봉하였다. 채취한 토양시료는 실험실로 운반하여 생중량을 측정하고, 온풍 건조기에서 105°C로 48시간 이상 건조 후 토양 용적밀도와 건조된 토양 시료를 2 mm 체로 선별한 후 석력 함량비를 계산하였다. 또 하나의 토양시료는 2주 이상 음건 후 2 mm 체(US standard No. 10)로 토양 성질 분석용 시료를 조제하였다. 토양 pH와 전기전도도(Electrical conductivity, EC)는 토양 pH 측정기(Model-735, ISTEC, Korea)와 EC 측정기(Thermo Fisher Scientific Orion 3-star, Singapore)로 측정하였다. 토양 탄소 및 질소 농도 분석은 40 mesh 체를 이용하여 시료를 조제하고 대용량 원소분석기(Vario Macro cube, Germany)로 분석하였다. 인, 포타슘, 칼슘, 마그네슘 등은 2.5 g의 건조 토양을 암모늄 아세테이트 용액 55 ml를 첨가하고 16시간 동안 진공 추출한 용액을 ICP(Perkin Elmer Optima 5300, USA)를 이용하여 분석하였다(Kalra and Maynard, 1991).
3. 자료 분석
바이오매스 부위별 양분과 임상 및 토양 분석으로부터 얻어진 자료는 조림지와 확산지의 비교를 위해 SAS의 Proc T-test를 이용하여 P < 0.05 수준에서 검정하였다(SAS Institute Inc., 2003).
결과 및 고찰
1. 조사구 현황
솜대 확산지의 솜대 밀도는 조림지의 약 57% 정도로(Table 1), 일본에서 조사된 맹종죽의 편백 임분 확산지 38%에 비해서는 높은 편이었다(Umemura and Takenaka, 2015). 일본에서 조사된 맹종죽 확산지는 편백 고사목 밀도가 높을 경우 맹종죽 밀도가 높은 것으로 보고되고 있으나(Umemura and Takenaka, 2015), 본 조사구의 고사목 밀도와 솜대 밀도 사이에 뚜렷한 관계는 없었다. 본 연구 조사구의 솜대 조림지 밀도는 2009년 동일지역에서 조사된 12,950 주 ha−1(Jung et al., 2017), 전라남도 담양의 11,700 주 ha−1(Park et al., 2017)와 유사하였다. 평균 흉고직경은 조림지가 4.84 cm로 2009년 동일지역에서 조사된 흉고직경 4.2 cm보다는 약간 크게 나타났으나(Jung et al., 2017), 죽순 생산을 위해 퇴비가 시비된 담양지역의 흉고직경 6.53 cm보다는 작았다(Park et al., 2017). 평균 흉고직경은 확산지가 5.68 cm로 조림지 4.84 cm에 비해 크게 나타나 솜대 밀도가 흉고직경 크기에 어느 정도 기여하고 있는 것으로 사료되며, 전라남도 순천에서 조사된 솜대 밀도 8,600 주 ha−1의 평균 흉고직경 5.8 cm와는 큰 차이가 없었다(Park and Ryu, 1996).
2. 솜대 표본의 현황
벌채된 솜대 바이오매스 표본의 평균 흉고직경은 조림지 5.73 cm에 비해 확산지가 5.93 cm로 약간 크게 나타났으나 유의적인 차이는 없었다(Table 2). 간고나 지하고 등도 유의적인 차이는 없었으나 수관 폭의 경우 확산지가 2.38 m, 조림지 1.35 m로(P < 0.05) 솜대 밀도가 높았던 조림지의 수관 폭이 작았다. 대나무 부위별 건중량은 확산지가 조림지에 비해 약간씩 크게 나타났으며, 이는 벌채된 확산지 표본의 평균 흉고직경이 조림지에 비해 0.20 cm 정도 크기 때문으로 사료된다.
Table 2.
General characteristics of sampled bamboos in plantation and expansion treatments of Phyllostachysnigra var. henonis.
|
Treatment |
DBH (cm) |
Height (m) |
Clear culm length (cm) |
Crown width (cm) |
Culm (g bamboo−1) |
Branches (g bamboo−1) |
Foliage (g bamboo−1) |
Total (g bamboo−1) |
|
Plantation |
5.73 (0.74)a |
12.0 (1.3)a |
6.2 (1.4)a |
1.35 (0.29)b |
5,865 (1,908)a |
580 (171)a |
494 (175)a |
6,939 (2,211)a |
|
Expansion |
5.93 (0.63)a |
12.3 (1.0)a |
5.8 (0.9)a |
2.38 (0.07)a |
5,901 (1,340)a |
682 (98)a |
485 (86)a |
7,068 (1,430)a |
Download Excel Table
3. 바이오매스 부위별 양분 농도
바이오매스 부위별 양분 농도 중 줄기, 가지, 잎의 질소 농도는 조림지가 확산지에 비해 높았다(Table 3). 이는 매년 2차에 걸친 복합비료 시비가 원인으로 사료되며 유사한 결과가 진주지역 맹종죽 조림지의 시비구와 비시비구에서도 관찰된 바 있다(Kim et al., 2018). 인의 경우 질소 농도와 유사한 경향을 보여 줄기, 가지, 뿌리의 인 농도는 조림지가 확산지에 비해 높아 시비의 영향이 뚜렷하였으나, 포타슘과 칼슘 농도는 조림지와 확산지 사이에 유의적인 차이가 없었다. 마그네슘 농도는 뿌리를 제외하고 줄기, 가지, 잎 모두 조림지가 확산지에 비해 양분 농도가 높게 나타나 질소나 인 농도 등에서 나타난 결과와 유사하였다. 양분 농도의 크기는 잎의 경우 조림지는 질소 > 칼슘 > 포타슘 > 인 > 마그네슘 순이었으나, 확산지는 질소 > 포타슘 > 칼슘 > 인 > 마그네슘 순으로 다르게 나타났다(Table 3). 확산지는 경상남도 진주 맹종죽의 잎 양분 농도 크기 질소 > 포타슘 > 칼슘 > 인 > 마그네슘과 동일하였으나(Kim et al., 2018), 전라남도 담양에서 7월에 조사된 솜대 잎의 양분 포타슘 > 질소 > 인 > 칼슘 > 마그네슘과는 차이가 있었다(Park et al., 2017). 이는 두 지역 사이에 5월 과 7월의 조사 시기 차로 인한 양분 농도 크기에 차이가 발생한 것으로 사료된다. 대나무의 부위별 양분 농도는 양분 재흡수나 토양 양분 흡수 기작의 차이로 인하여 계절적 변동이 매우 큰 것으로 알려져 있으며(Wu et al., 2009; Umemura and Takenaka, 2015), 흉고직경(Park et al., 2017), 밀도(Wu et al., 2009), 관리방법(Song et al., 2017) 등에 따라 다르게 나타나는 것으로 알려져 있다.
Table 3.
Nutrient concentration of bamboo components in plantation and expansion treatments of Phyllostachys nigra var. henonis.
|
Component |
Treatment |
Nutrient (mg g−1) |
|
C |
N |
P |
K |
Ca |
Mg |
|
Culm |
Plantation |
450.87 (0.68)a |
2.85 (0.37)a |
0.83 (0.08)a |
4.62 (1.46)a |
0.37 (0.11)a |
0.41 (0.10)a |
|
Expansion |
449.13 (0.84)a |
0.95 (0.11)b |
0.28 (0.08)b |
2.73 (0.30)a |
0.43 (0.04)a |
0.16 (0.03)b |
|
Branches |
Plantation |
444.21 (1.47)a |
7.01 (0.97)a |
0.99 (0.18)a |
6.53 (1.99)a |
0.39 (0.04)a |
0.30 (0.02)a |
|
Expansion |
451.34 (9.71)a |
3.09 (0.17)b |
0.30 (0.04)b |
3.17 (0.32)a |
0.39 (0.02)a |
0.12 (0.02)b |
|
Foliage |
Plantation |
422.52 (3.80)a |
25.64 (1.11)a |
1.38 (0.13)a |
5.41 (0.73)a |
6.94 (0.67)a |
1.33 (0.15)a |
|
Expansion |
413.76 (8.56)a |
21.25 (0.58)b |
1.08 (0.02)a |
7.09 (0.36)a |
5.52 (0.49)a |
0.71 (0.05)b |
|
Root |
Plantation |
349.40 (17.92)a |
4.43 (0.16)a |
0.78 (0.03)a |
2.49 (0.18)a |
1.10 (0.27)a |
1.11 (0.14)a |
|
Expansion |
342.25 (18.24)a |
3.74 (0.34)a |
0.61 (0.02)b |
2.12 (0.17)a |
3.23 (1.50)a |
1.15 (0.03)a |
Download Excel Table
4. 임상의 양분 농도
임상의 질소와 인 농도는 시비가 실시된 조림지가 확산지보다 약간 높았으나 유의적인 차이는 없었다(Table 4). 이는 확산지의 경우 일부 잔존 임목으로부터 유입되는 임목의 낙엽이 양분 농도에 영향을 끼칠 수 있으나, 확산지의 임상은 주로 솜대 낙엽으로 구성되어 있었기 때문이다. 임상의 양분 농도 크기는 탄소 > 질소 > 칼슘 > 마그네슘 > 포타슘 > 인 순으로 조림지와 확산지 모두 동일하였다.
Table 4.
Nutrient concentration of the forest floor in plantation and expansion treatments of Phyllostachys nigra var. henonis.
|
Treatment |
Nutrient (mg g−1) |
|
C |
N |
P |
K |
Ca |
Mg |
|
Plantation |
395.38 (12.15)a |
14.96 (1.60)a |
1.09 (0.16)a |
1.20 (0.15)a |
8.19 (1.54)a |
1.32 (0.24)a |
|
Expansion |
402.60 (15.77)a |
12.26 (1.20)a |
0.69 (0.07)a |
1.31 (0.20)a |
8.62 (0.98)a |
1.68 (0.36)a |
Download Excel Table
5. 토양의 물리·화학적 성질
1) 토양 용적밀도, 석력함량, 토양 pH, 전기전도도
토양 20~30 cm 깊이의 전기전도도(EC)를 제외하고 토양 깊이별 석력함량, 토양 pH, 토양 용적밀도는 조림지와 확산지 사이에 유의적인 차이가 없었다. 토양 전기전도도는 모든 깊이에서 조림지가 확산지에 비해 높은 값을 보이고 있어 시비의 영향이 어느 정도 나타난 것으로 사료된다. 토양 용적밀도는 조림지가 확산지에 비해 낮은 경향을 보였으며, 평균 용적밀도는 조림지가 확산지에 비해 유의적으로 낮았다(Table 5). 이는 솜대 밀도가 높은 조림지의 경우 지하경 및 뿌리의 발달로 인한 토양 내 공간 증가로 토양 용적밀도가 낮게 나타났거나, 토양 용적밀도와 밀접한 관계를 가지는 조림지의 토양 유기탄소 농도가 확산지에 비해 2배 정도 높았기 때문으로 사료된다(Figure 2). 석력 함량은 조림지 0.41~0.51 g g−1, 확산지 0.32~0.37 g g−1 정도 분포하여 조림지가 높은 비율을 보였으며, 소규모 임분 단위에서도 석력 함량은 조림지와 확산지 사이에 공간적 변동이 크게 나타났다. 토양 pH는 유의적인 차이는 없었으나 조사한 0~30 cm 깊이 모두 확산지가 조림지에 비해 높은 경향을 보였으며, 이는 대나무 확산지는 토양 표면에서 교환성 양이온의 활성이 높아 토양 pH가 증가하는 것으로 보고된 결과와 유사하였다(Umemura and Takenaka, 2015).
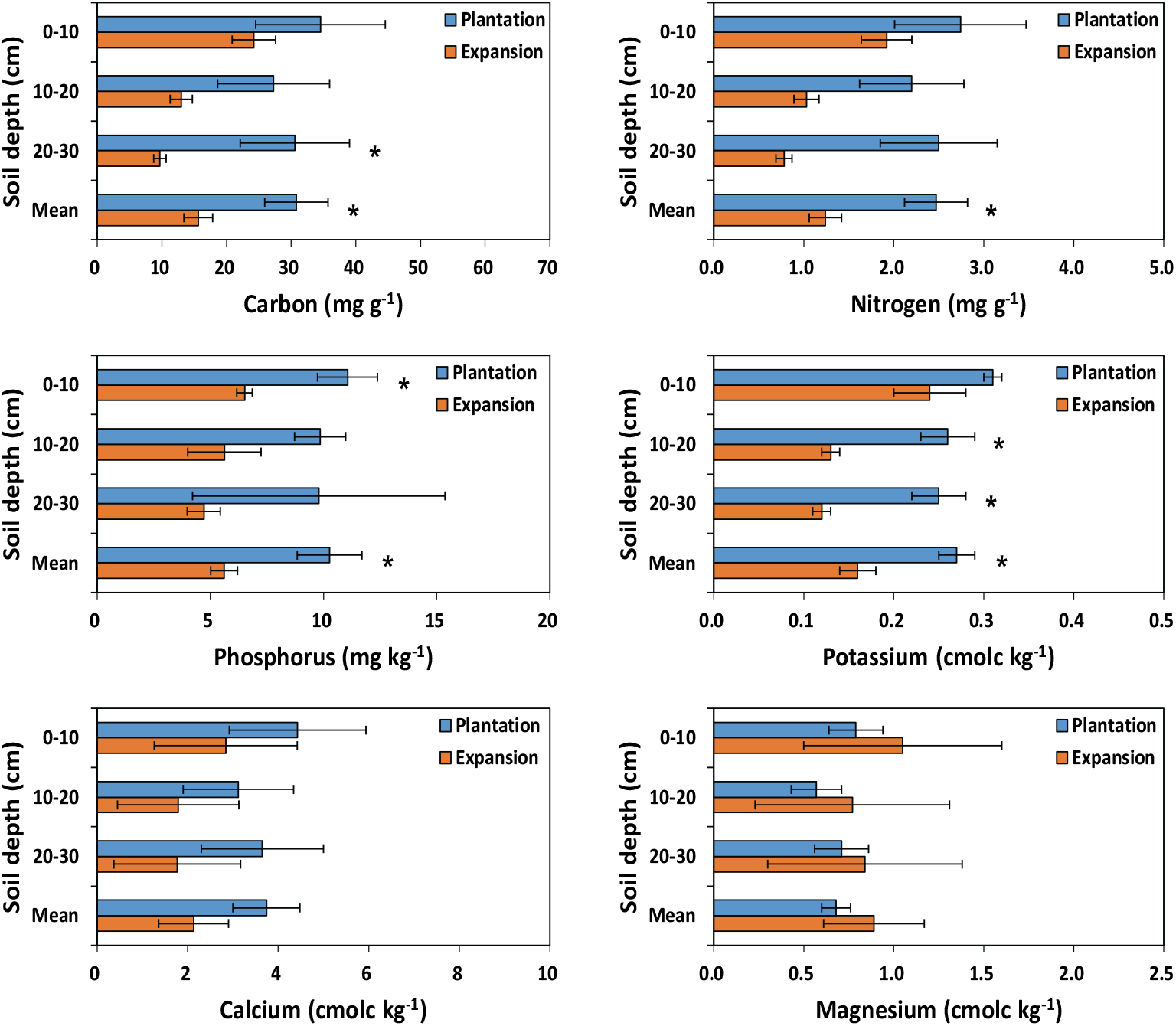
Figure 2.
Chemical properties at three soil depth in plantation and expansion treatments of Phyllostachys nigra var. henonis. Horizontal bars are standard error (n=6). Asterisks between plantation and expansion treatments represent a significant difference at P < 0.05.
Download Original Figure
Table 5.
Coarse fragment, soil pH, EC, and bulk density at three soil depth in plantation and expansion treatments of Phyllostachys nigra var. henonis.
|
Depth (cm) |
Treatment |
Coarse fragment (g g−1) |
Soil pH |
EC (uS cm−1) |
Bulk density (g cm−3) |
|
0-10 |
Plantation |
0.51 (0.07)a |
4.97 (0.15)a |
427.45 (75.36)a |
0.76 (0.06)a |
|
Expansion |
0.34 (0.08)a |
5.06 (0.29)a |
241.43 (105.54)a |
0.92 (0.06)a |
|
10-20 |
Plantation |
0.49 (0.08)a |
4.79 (0.17)a |
532.38 (157.97)a |
0.88 (0.06)a |
|
Expansion |
0.37 (0.07)a |
5.29 (0.40)a |
160.83 (22.04)a |
0.98 (0.04)a |
|
20-30 |
Plantation |
0.41 (0.05)a |
5.08 (0.23)a |
369.87 (85.16)a |
0.79 (0.16)a |
|
Expansion |
0.32 (0.08)a |
5.20 (0.51)a |
123.50 (14.25)b |
1.05 (0.05)a |
|
Mean |
Plantation |
0.47 (0.04)a |
4.93 (0.10)a |
449.9 (64.6)a |
0.81 (0.04)b |
|
Expansion |
0.34 (0.04)b |
5.18 (0.21)a |
175.25 (35.99)b |
0.97 (0.03)a |
Download Excel Table
2) 토양의 화학적 성질
조림지와 확산지 토양 20~30 cm의 유기탄소, 0~10 cm 인, 10~30 cm 포타슘 농도를 제외하고 조림지와 확산지 사이 토양 양분 농도에 유의적인 차이가 없었다(Figure 2). 토양 유기탄소 농도는 모든 토양 깊이에서 조림지가 확산지에 비해 높았으며, 평균 유기탄소 농도는 조림지가 30.80 mg g−1로 확산지 15.64 mg g−1에 비해 유의적으로 높았다. 이는 리그닌 같은 분해 저항성 물질의 함량이 낮은 대나무 낙엽은 쉽게 부식화(humification)가 발생하고(Shiau and Chiu, 2017), 밀도가 높은 조림지의 경우 뿌리 등에 의한 유기물의 토양 환원이 크기 때문으로 사료된다. 전질소의 경우도 조림지 2.47 g kg−1로 확산지 1.24 g kg−1에 비해 유의적으로 높았다. 한편, 확산지의 낮은 전질소 농도는 유기탄소 농도와 전질소 농도가 강한 상관을 보이기 때문이며(Weil and Brady, 2017), Shiau and Chiu (2017)은 맹종죽이 확산된 삼나무 임분의 경우 미생물에 의해 쉽게 분해가 가능한 대나무 낙엽·낙지 유입으로 인하여 토양 내 탄소 및 질소 총량의 감소를 관찰한 바 있다. 유사한 결과로, Bai et al.(2016b)는 산림지역에 대나무가 확산되는 경우 급속한 생장 및 단기간에 높은 밀도로 인한 높은 질소 요구도로 표토 내 토양 유기탄소와 전질소 농도가 감소한다고 보고한 바 있다. 토양 내 잔류 특성이 강한 인의 경우(Weil and Brady, 2017), 조림지 10.27 mg kg−1로 확산지 5.61 mg kg−1에 비해 유의적으로 높게 나타났으며, 이는 매년 복합비료 시비 영향으로 사료된다.
포타슘은 조림지 0.27 cmolc kg−1, 확산지 0.16 cmolc kg−1로 조림지가 확산지에 비해 유의적으로 크게 나타나(Figure 2) 시비의 영향이 뚜렷하였다. 칼슘의 경우 조림지와 확산지 사이에 유의적인 차이는 없었지만 조림지가 확산지에 비해 높은 경향을 보이고 있다. 일본에서 조사된 왕대 조림지의 칼슘 농도는 인접한 졸참나무 임분에 비해 높은 농도를 보였으며 주로 수관통과우나 수간류의 높은 칼슘 농도가 원인으로 제시된 바 있다(Sakai and Tadaki, 1997).
결 론
대나무 임분 조성 약 25년이 경과하고 지속적으로 시비가 실시된 솜대 조림지의 바이오매스 부위별 질소, 인, 마그네슘 농도나 토양 0~30 cm 깊이의 유기탄소, 전질소, 인, 포타슘 농도 등은 조림지가 확산지에 비해 유의적으로 높았으며 이는 지속적인 시비의 영향으로 사료된다. 한편, 솜대 확산지의 경우 토양 양분 농도가 낮음에도 불구하고 솜대 확산이 발생하고 있는 것은 토양의 비옥도가 상대적으로 낮은 인접지역으로 솜대가 지속적으로 확산될 수 있음을 시사한다. 본 연구가 제한된 공간적 범위에서 실시되었지만, 토양 양분 상태는 대나무 확산에 큰 영향을 끼치지 않는 것으로 사료되며, 추후 대나무 확산 기작의 보다 나은 이해를 위해서는 조림지, 확산지 및 미확산 자연림에 대한 입지환경, 타 식생을 포함하는 임분 구조, 토양 미량원소 등과 같은 다양한 요인과의 관계에 대한 연구가 필요할 것으로 사료된다.
감사의 글
본 논문은 국립산림과학원 산림바이오소재연구소의 연구용역 “대나무 황폐지·확산 죽종 별 임지 양료순환 특성분석(I)”과 국토교통부/국토교통과학기술진흥원(과제번호 20UMRG-B158194-01)의 연구비 지원으로 수행되었습니다. 저자들은 본 논문의 체계적 구성에 도움을 주신 두 분의 심사자님께 감사드립니다.
References
Bai, S., Wang, Y., Conant, R.T., Zhou, G., Xu, Y., Wang, N., Fang, F. and Chen, J. 2016a. Can native clonal moso bamboo encroach on adjacent natural forest without human intervention? Scientific Reports 6: 31504.

Bai, S., Conant, R., Zhou, G., Wang, Y., Wang, N., Li, Y. and Zhang, K. 2016b. Effects of moso bamboo encroachment into native, broad-leaved forests on soil carbon and nitrogen pool. Scientific Reports 6: 31480.
Christanty, L., Mailly, D. and Kimmins, J.P. 1996. “Without bamboo, the land dies”: biomass, litterfall, and soil organic matter dynamics of a Javanese bamboo talun-kebun system. Forest Ecology and Management 87(1-3): 75-78.
Griscom, B.W. and Ashton, P.M.S. 2003. Bamboo control of forest succession:
Guadua sarcocarpa in Southeastern Peru. Forest Ecology and Management 175(1-3): 445-454.
Inoue, A. Tateishi, H., Sakuta, K., Yamamoto, K., Mizoue, N. and Kitahara, F. 2012. Relationship of light environment to stand attributes in a stand of bamboo,
Phyllostachys pubescens. Ecological Engineering 38(1): 135-139.

Jung, S.Y., Lee, K.S., Park, J.H., Park, Y.B., Yoo, B.O., Kim, C. and Cho, H.S. 2017. Biomass equations and accumulation of
Phyllostachys pubescens,
P. bambusoides, and
P. nigra var.
henonis stands. Journal of Agriculture and Life Science 51(5): 27-38.
Kalra, Y.P. and Maynard, D.G. 1991. Methods Manual for Forest Soil and Plant Analysis. Northern Forest Centre, Edmonton, Alberta. Information Report NOR-X-319. pp. 115.
Kim, C, Baek, G., Yoo, B.O., Jung, S.Y. and Lee, K.S. 2018. Regular fertilization effects on the nutrient distribution of bamboo components in a Moso bamboo (
Phyllostachys pubescens (Mazel) Ohwi) stand in south Korea. Forests 9(11): 671.
Nath, A.J., Lal, R. and Das, A.K. 2015. Managing woody bamboos for carbon farming and carbon trading. Global Ecology and Conservation 3: 654-663.
National Institute of Forest Science. 2016. Distribution Status of Bamboo Forest Resources in Korea, December 2016 (No. 16-27), Seoul, Korea.
Okutomi, K., Shinoda, S. and Fukuda, H. 1996. Causal analysis of the invasion of broad forest by bamboo in Japan. Journal of Vegetation Science 7(5): 723-728.
Park, I.H. and Ryu, S.B. 1996. Biomass, net production and nutrient distribution of bamboo
Phyllostachys stands in Korea. Journal of Korean Forestry Society 85(3): 453-461.
Park, S.W. Baek, G., Cho, H.S., Yoo, B.O., Jung, S.Y., Lee, K.S. and Kim, C. 2017. Nutrient distribution of culm, branches and leaf in
Phyllostachys bambusoides and
Phyllostachys nigra var.
nenosis. Journal of Korean Forest Society 106(4): 388-396.
Sakai, Y. and Tadaki, Y. 1997. Characteristics of stem flow and throughfall in a Madake and in a Mousouchiku stands. Bamboo Journal 14: 28-35.
SAS Institute Inc. 2003. SAS/STAT Statistical Software. Version 9.1 SAS Publishing Cary, NC.
Shiau, Y.J. and Chiu, C.Y. 2017. Changes in soil biochemical properties in a cedar plantation invaded by Moso bamboo. Forests 8(7): 222.
Song, Q. Lu, H., Yang, J. Yang, G. and Yang, Q. 2017. Accessing the impacts of bamboo expansion on NPP and N cycling in evergreen broadleaved forest in subtropical China. Scientific Reports 7: 40383.
Suzuki, S. and Nakagoshi, N. 2008. Expansion of bamboo forests caused by reduced bamboo-shoot harvest under different natural and artificial conditions. Ecological Research 23: 641-647.
Umemura, M. and Takenaka, C. 2015. Changes in chemical characteristics of surface soils in hinoki cypress (
Chamaecyparis obtusa) forests induced by the invasion of exotic Moso bamboo (
Phyllostachys pubescens) in central Japan. Plant Species Biology 30(1): 72-79.
Wang, Y., Bai, S., Binkley, D., Zhou, G. and Fang, F. 2016. The independence of clonal shoot's growth from light availability supports Moso bamboo invasion of closed- canopy forest. Forest Ecology and Management 368: 105-110.
Weil, R.R. and Brady, N.C. 2017. The Nature and Properties of Soils. 15th edition, Pearson, pp. 1104.
Wu, F.Z., Yang, W.Q., Wang, K.Y., Wu, N. and Lu, Y.J. 2009. Effect of stem density on leaf nutrient dynamics and nutrient use efficiency of dwarf bamboo. Pedosphere 19(4): 496-504.
Yoo, B.O., Park, J.H., Park, Y.B., Jung, S.Y., Lee, K.S. and Kim, C. 2017. Assessment of expansion characteristics and classification of distribution types for bamboo forests using GIS. Journal of the Korean Association of Geographic Information Studies 20(4): 55-64.
Yuen, J.Q., Fung, T. and Ziegler, A.D. 2017. Carbon stocks in bamboo ecosystems worldwide: Estimates and uncertainties. Forest Ecology and Management 393: 113-138.
Zaninovich, S.C., Montti, L.F., Alvarez, F. and Gatti, M.G. 2017. Replacing trees by bamboos: Changes from canopy to soil organic carbon storage. Forest Ecology and Management 400: 208-217.