Article
소나무재선충병 피해지에 식재된 편백의 낙엽 · 낙지에 의한 탄소 및 질소 유입량
강현철
1, 백경원
1, 최병길
1, 하지석
1, 김춘식
1,*
Carbon and Nitrogen Inputs by Litterfall of Chamaecyparis obtusa Planted in Pine Wilt Disease-disturbed Forests
Hyeon Cheol Kang
1, Gyeongwon Baek
1, Byeonggil Choi
1, Jiseok Ha
1, Choonsig Kim
1,*
1Department of Forest Resources, Gyeongsang National University, Jinju 52725, Korea
© Copyright 2021 Korean Society of Forest Science. This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received: Jan 13, 2021; Revised: Mar 01, 2021; Accepted: Mar 02, 2021
Published Online: Mar 31, 2021
요약
소나무재선충병 피해목을 모두베기하거나 미피해 곰솔을 잔존하고 편백을 수하식재 한 15년생 편백 조림지를 대상으로 1년 동안(2018년 12월~2019년 12월) 낙엽·낙지에 의한 탄소 및 질소 유입량을 조사하였다. 편백의 평균 흉고직경은 모두베기구가 12.10 cm로 수하식재구의 9.42 cm에 비해 유의적으로 크게 나타났다. 편백 낙엽의 유기탄소 및 질소 농도는 모두베기구나 수하식재구 사이에 유의적인 차이(P > 0.05)가 없었으나, 낙엽의 C/N비는 8월~10월에 유입된 낙엽이 93으로 12월~3월 사이에 유입된 낙엽의 143보다 유의적으로 낮았다. 낙엽·낙지 구성 성분의 탄소 농도는 곰솔 가지, 편백 낙엽, 곰솔 낙엽 등의 탄소 농도가 활엽수나 기타 물질에 비해 유의적으로 높았으며, 질소 농도는 활엽수 잎, 편백 녹엽(green leaf), 기타 물질 등이 편백 낙엽이나 곰솔 가지 등에 비해 높은 농도를 보였다. C/N비는 곰솔 낙엽과 편백 낙엽이 기타 물질, 편백 녹엽, 활엽수 낙엽 등에 비해 유의적으로 높았다. 편백 낙엽에 의한 유기탄소 및 질소 유입량은 모두베기구가 773 kg C ha−1 yr−1와 6.95 kg N ha−1 yr−1로 수하식재구 78 kg C ha−1 yr−1와 0.70 kg N ha−1 yr−1에 비해 유의적으로 많은 양이 유입되었다. 그러나 총 유기탄소 및 질소 유입량은 수하식재구 3,765 kg C ha−1 yr−1와 47.6 kg N ha−1 yr−1, 모두베기구 1,290 kg C ha−1 yr−1와 17.2 kg N ha−1 yr−1로 수하식재구가 상층 곰솔의 낙엽·낙지 유입으로 인하여 3배 정도 많았다. 본 연구결과에 따르면 모두베기구에 조성된 편백의 흉고직경 생장은 수하식재구에 비해 양호하였으나, 낙엽·낙지에 의한 탄소 및 질소 유입량은 상당히 감소하는 것으로 나타났다.
Abstract
In this study, carbon (C) and nitrogen (N) inputs by the litterfall of Japanese cypress (Chamaecyparis obtusa Endlicher) planted in pine wilt disease-disturbed forests were determined. The study sites were located in Sacheon-si, Gyeongsangnam-do. Eight plots under two regeneration sites (DR: four plots of C. obtusa planted under slightly disturbed Pinus thunbergii stands; CR: four plots of C. obtusa planted following the clear-cutting of severely disturbed pine stands) were established to collect litterfall from December 2018 to December 2019. The growth of diameter at breast height (DBH) was significantly higher in the CR treatment (12.10 cm) than that in the DR treatment (9.42 cm). C and N concentrations and the C/N ratio in C. obtusa leaf litter did not differ significantly between the two regeneration treatments, but the C/N ratio was significantly lower in the leaf litter collected in October (93) relative to that collected in December (143). The C concentration of litterfall components was significantly higher in C. obtusa leaf litter and in P. thunbergii needle litter than in broadleaved and miscellaneous litter, whereas the N concentration in broadleaved and miscellaneous litter was significantly higher than that in the leaf litter of C. obtusa and in branch litter. Thus, the C/N ratio was significantly higher in C. obtusa leaf litter and branch litter compared with that in miscellaneous and broadleaved litter. Respective C and N inputs by leaf litter were 773 kg C ha−1 yr−1 and 6.95 kg N ha−1 yr−1 for the CR treatments, and 78 kg C ha−1 yr−1 and 0.70 kg N ha−1 yr−1 for the DR treatment. Total C and N inputs were higher for the DR treatment (3,765 kg C ha−1 yr−1 and 47.6 kg N ha−1 yr−1, respectively) than for the CR treatment (1,290 kg C ha−1 yr−1 and 17.2 kg N ha−1 yr−1, respectively). These results indicate that, for C. obtusa, the DBH growth in the CR treatment was superior to that in the DR treatment, but the C and N inputs by litterfall were considerably reduced in CR treatments.
Keywords: carbon; double-layer forest regeneration; Japanese cypress; litterfall; nitrogen; pine wilt disease
서 론
산림생태계의 낙엽·낙지를 통한 탄소 및 양분 유입은 토양생물의 에너지 및 영양원으로 제공되는 기능뿐만 아니라, 임상(forest floor) 및 토양 내 탄소 및 양분 축적량의 증가와 양분순환에 가장 중요한 요인이다. 그러나 낙엽·낙지량은 임령, 임분 종류, 임분 밀도, 위도, 기후요인(Berg and Laskowski, 2006)뿐만 아니라 간벌(Inagaki et al., 2008)이나 시비(Kim et al., 2013a) 같은 산림경영 활동 등에 의해서도 상당한 차이를 보이는 것으로 알려져 있다.
산림갱신을 위한 후계림 조성에는 모두베기, 산벌, 택벌, 패치 벌채 등을 통하여 목적 수종을 동령림이나 이단 이령림, 또는 복층림으로 조성하는 다양한 방법이 있다(Paquette et al., 2006; Diaci et al., 2017; Nolet et al., 2017). 그러나 모두베기에 의한 산림조성은 토양 답압, 낙엽층 유실, 토양 침식, 토양 양분량 감소 등 산림생태계의 다양한 기능에 부정적인 영향을 끼치는 것으로 알려져 있어, 최근에는 생물다양성 증진과 생태계서비스 기능을 향상할 수 있는 산림갱신의 중요성이 강조되고 있다(Paquette et al., 2006; O’Hara and Ramage, 2013; Nolet et al., 2017).
남부지역 소나무재선충병 피해지의 경우 피해목 벌채 위주로 작업이 실시되어(Jeon et al., 2011), 피해 극심지는 모두베기, 경미지역은 상층 임목이 잔존된 임분이 증가하고 있다. 이들 지역은 산림생태계 복원을 위해 모두베기나 수하식재를 통하여 소나무재선충병에 저항성을 보이는 편백을 조림하고 있다(Baek, 2020). 한편, 편백은 내음성이 강하여 낙엽활엽수 임분의 중층 및 하층에 우점하기도 하며(Kim et al., 2013c), 국내 편백 경영림의 경우 임지안정, 수원함양, 풍치 및 용재 공급적인 면에서 복층림 경영을 권장하고 있다(Kim et al., 2004). 복층림 조성지는 식재목의 경쟁식생 감소나 상층 잔존목을 통한 낙엽·낙지의 지속적인 공급으로 임목 벌채지의 급격한 산림환경 변화를 완화할 수 있는 것으로 알려져 있다(Kim et al., 1996). 한편, 국외에서 조사된 결과에 따르면 일본 교토지역 소나무재선충병 발생 소나무 임분은 편백의 천연갱신에 의한 순림 조성이 가능한 것으로 알려져 있다(Morimoto and Yoshida, 2005; Nakanishi et al., 2009).
임목의 생장 반응은 광합성 부위인 엽량과 밀접한 관계를 가지기 때문에(Lehtonen et al., 2008), 소나무재선충병 피해지에 식재된 편백의 질적, 양적 생장에 영향을 미칠 수 있는 엽량의 정량적인 평가는 편백의 생장 해석 및 양분순환에 중요한 의미를 갖는다. 그러나 임목이나 임분 내 엽량의 정량적 평가는 임목 벌채 같은 파괴적인 방법을 포함하기 때문에 낙엽·낙지 수집을 통한 엽량의 정량화와 양분 동태 조사는 비 파괴적 방법으로 생장 및 임분 변화를 구명할 수 있는 수단이 될 수 있다. 본 연구는 소나무재선충병 피해지에 모두베기나 수하식재된 편백을 대상으로 식생복원 방법이 편백의 생장과 낙엽·낙지에 의한 탄소 및 질소 유입량에 미치는 영향을 조사하기 위한 목적으로 수행하였다.
재료 및 방법
1. 일반현황
조사지는 경상남도 사천시 곤양면 검정리 산 34번지(35º04’N, 128º02’E)로 소나무재선충병 피해가 극심하였던 지역이다(Kim et al., 2010). 이 지역의 산림토양은 퇴적암인 사암이나 혈암을 모재로 생성된 적색계갈색약건산림토양형(rB2)이 분포하고 있으며, 조사지의 해발고는 30 m~40 m, 사면방위는 남서 방향이었다. 조사구는 2000년대 초반 소나무재선충병이 발병한 후 피해목에 대한 선택적 벌채가 이루어지고, 2008년 편백 4년생을 상층 곰솔 아래 수하식재한 지역과 소나무재선충병 극심지를 모두베기 후 편백을 식재한 모두베기 지역을 대상으로 하였다(Figure 1). 2018년 12월 조사구 설치 시 편백의 임령은 15년생이었다.
Figure 1.
Location of the study site in pine wilt disease-disturbed forests (a: under-planted site; b: collection of green needle; c: DR: under-planted site, CR: clearcut site; d: clearcut site in 2007; e: clearcut site in 2019).
Download Original Figure
2. 낙엽·낙지량과 탄소 및 질소 농도 분석
낙엽·낙지량은 각 처리구 내 10 m × 10 m 조사구를 4개 설치하고 2018년 12월 8일 조사구 중심 부분에 수집면적 0.25 m2의 원형 낙엽수집기를 지상으로부터 60 cm 높이에 각 처리구 당 1개씩(총 8개) 설치하였다. 낙엽·낙지는 1년 동안 총 5회(2019년 3월 9일, 6월 6일, 8월 5일, 10월 23일, 12월 13일) 수집하였다. 수집된 낙엽은 지퍼 비닐주머니에 넣어 실험실로 운반하고 65ºC 건조기에서 항량에 도달할 때까지 건조하였으며, 건조된 시료는 잎, 가지, 생식기관을 포함하는 기타 물질 등으로 분류하고 무게를 측정하였다. 건조된 시료는 분쇄기로 분말을 만든 후, 0.420 mm체(ASTM No. 40)를 이용하여 분석 시료를 조제하고 원소분석기(Thermo Scientific, Flash 2000, Italy)를 이용하여 탄소 및 질소 농도를 분석하였다. 한편, 일부 유입량이 적었던 낙엽·낙지 시료와 8월 5일 채취 시료는 분석 시료 양이 적어 10월 시료와 혼합하여 분석하였다. 낙엽·낙지의 부위별 탄소 및 질소 농도는 낙엽·낙지 건중량과 곱하여 처리구의 탄소 및 질소 유입량으로 환산하였다.
3. 임목 및 토양 특성 조사
조사구 내 임목의 흉고직경 생장량은 2018년과 2019년 12월에 직경테이프를 이용하여 조사하였으며, 수고는 편백을 대상으로 12 m 수고측정봉(Senshin, Japan)으로 측정하였다. 조사구의 곰솔 바이오매스는 인근 진주에서 조사 된 곰솔 IV 영급의 흉고직경을 독립변수로 하는 바이오매스 상대생장식 -0.5346+2.0426×Log10(DBH)(Kim et al., 2013b)과 편백은 우리나라 남부지역을 대상으로 개발된 지위지수 12의 흉고직경을 독립변수로 하는 상대생장식 -0.9483+2.3257×Log10(DBH)을(Kim et al., 2020) 이용하였다. 토양 성질은 깊이 0~5 cm를 대상으로 토양 입경분포는 비중계법, 유기탄소와 전질소는 대용량 원소분석기(Vario Macro cube, Germany), 유효인산은 NH4F와 HCl에 의해 추출된 용액을 UV 분광분석기(Jenway 6505, UK)로 측정하였다. 치환성 포타슘, 칼슘, 마그네슘의 경우 2.5 g의 건조 토양에 55 ml 암모늄 아세테이트 용액을 첨가한 후 16시간 동안 진공 추출기(Vacuum Extractor, Model 24VE, USA)로 용액을 추출하고, ICP-OES(Perkin Elmer Optima 5300 DV, USA)를 이용하여 분석하였다(Kalra and Maynard, 1991).
4. 토양 환경요인
토양 수분함량, 토양 온도, 토양 pH, 토양 전기전도도(EC) 같은 환경요인은 2019년 1월에서 7월까지 매월 1회씩 조사하였다. 토양 온도는 디지털 토양 온도 측정기(K-Type, Summit, Korea)를 이용하여 측정하였으며, 토양 수분, 토양 pH, 토양 EC는 코어 샘플러를 이용하여 채취한 0~5 cm 깊이의 토양 시료를 분석하였다. 토양 pH는 1:5(토양:증류수) 혼합액을 대상으로 이온전극 pH측정기(Model 735P, istek, Korea), 토양 EC는 EC측정기(Orion 3-Star, Thermo Scientific, Singapore)로 측정하였다. 토양 수분함량은 105ºC 건조기에서 24시간 이상 건조 후 계산하였다.
5. 자료 분석
수집된 자료는 처리 및 수집 시기나 낙엽·낙지 부위 및 수집 시기 등을 2원 분산분석(two-way ANOVA)을 이용하여 주요인(갱신방법, 낙엽·낙지 구성요인, 낙엽·낙지 수집 시기)과 상호작용 요인에 대한 유의성을 P < 0.05 수준에서 검정하였으며, 처리 간에 유의성이 인정되는 경우 평균 간의 비교는 Tukey 방법을 이용하였다(SAS Institute, 2003).
결과 및 고찰
1. 임분 및 토양 특성
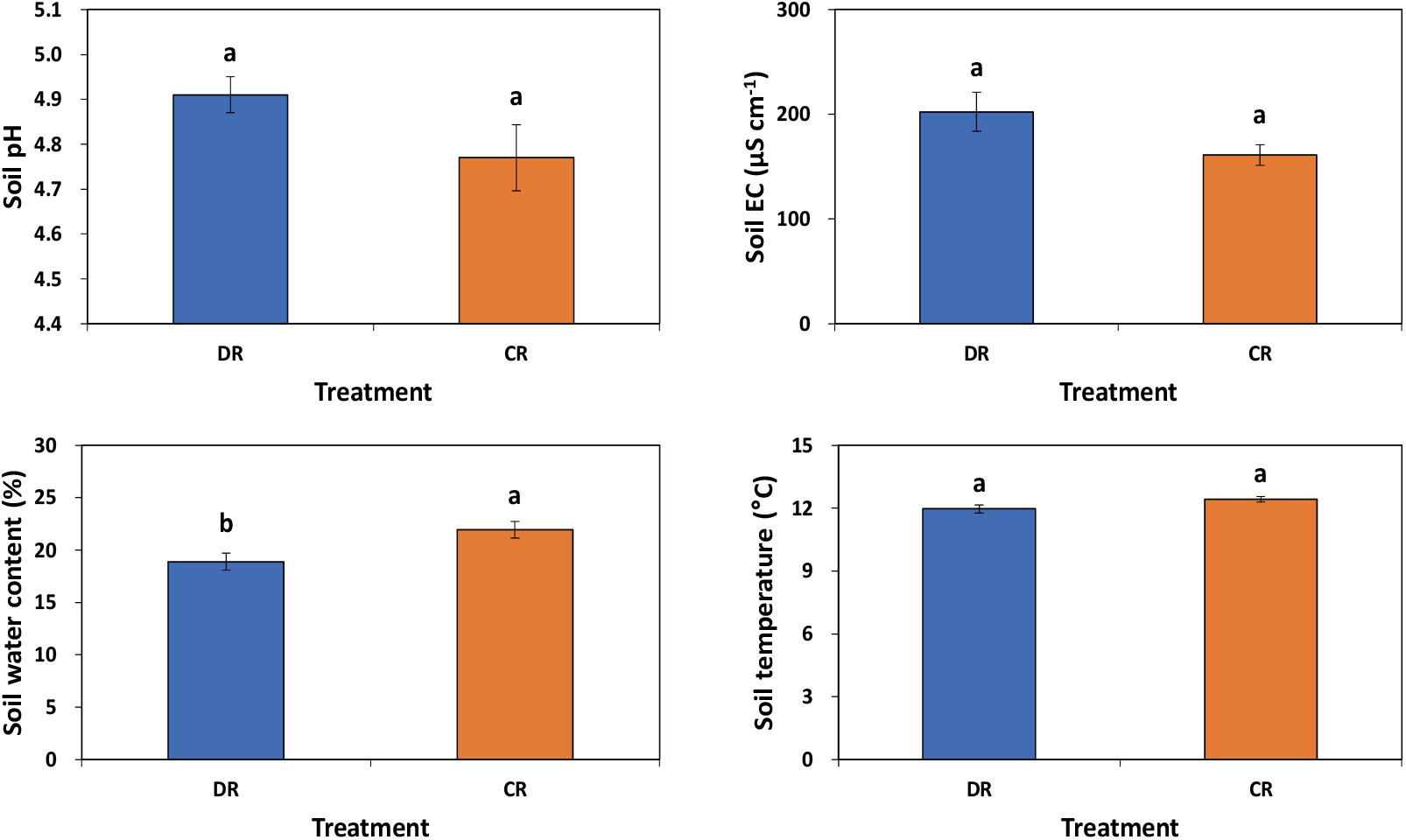
수하 식재구의 임분 밀도는 편백 900본 ha−1으로 모두베기구 1,050본 ha−1에 비해 150본 ha−1 정도가 낮은 값이나 상층에 잔존된 곰솔이 275본 ha−1 정도 분포하여, 총 임분 밀도는 1,175본 ha−1였다. 흉고직경 생장량은 2018년 모두베기구가 10.95 cm, 2019년 12.10 cm로 수하식재구의 8.45 cm와 9.42 cm에 비해 각 2.50 cm와 2.68 cm 정도 유의적으로 크게 나타났다(Table 1). 이는 모두베기구의 경우 광이나 토양 수분환경이 수하식재구에 비해 양호하기 때문으로 평균 토양 수분함량의 경우 모두베기구가 21.9%로 수하식재구 18.9%에 비해 유의적으로 높았다(Figure 2). 한편, 일본 큐슈지역에서 조사된 결과에 따르면 상층 잔존 임목에 의한 하층의 낮은 광 조건은 편백 생장에 부정적인 요인으로 제시된 바 있다(Hirata et al., 2015). 직경 생장과 유사하게 수고 생장도 모두베기구가 수하식재구에 비해 약 70 cm 정도 크나 유의적인 차이는 없었다. 그러나 수하식재구 편백의 지상부 임목 바이오매스는 22,523 kg ha−1로 모두베기구 40,935 kg ha−1와 상당한 차이가 있었다(Table 1). 모두베기구의 편백 바이오매스는 평균 흉고직경 10.9 cm, 밀도 2,300본 ha−1를 가지는 순천지역 14년생 편백 조림지 임목 바이오매스 76,095 kg ha−1(Park et al., 2000)에 비해 낮았으며, 이는 두 지역 편백의 흉고직경 생장에 큰 차이가 없음을 고려할 때 임분 밀도 차가 원인으로 사료된다. 한편, 수하식재구에 잔존된 곰솔의 지상부 임목 바이오매스는 95,867 kg ha−1로 인접지역인 진주에서 조사된 35년생 85,170 kg ha−1과 51년생 121,600 kg ha−1 (Kim et al., 2013b)의 범위에 분포하였다.
Table 1.
General stand characteristics in under-planted (DR) and clearcut (CR) regeneration treatments of Chamaecyparis obtusa.
|
Treatment |
Species |
Stand density (tree ha−1) |
DBH (cm) |
Height (cm) |
Aboveground biomass (kg ha−1) |
|
2018 |
2019 |
2018 |
2019 |
2018 |
2019 |
|
DR |
Chamaecyparis obtusa (C) |
900 (168) |
8.45 (0.94)b |
9.42 (0.99)b |
- |
665 (50)a |
17,979 (7,179) |
22,523 (8,610) |
|
Pinus thunbergii (P) |
275 (85) |
30.2 (0.57) |
31.42 (0.74) |
- |
- |
90,743 (33,467) |
95,867 (33,611) |
|
C + P |
1,175 (137) |
- |
- |
- |
- |
108,722 (299,48) |
118,396 (29,074) |
|
CR |
Chamaecyparis obtusa
|
1,050 (104) |
10.95 (0.38)a |
12.10 (0.41)a |
- |
736 (29)a |
32,551 (3,497) |
40,935 (4,604) |
Download Excel Table
Figure 2.
Mean soil pH, soil EC, soil water content, and soil temperature in under-planted (DR) and clearcut (CR) regeneration treatments of Chamaecyparis obtusa. Vertical bars indicate standard error. Different letters on the bars represent a significant difference at P < 0.05.
Download Original Figure
토양 입경 분포 중 모래 함량은 모두베기구가 33.5%로 수하식재구 17.8%에 비해 높게 나타났으나 점토 함량은 수하식재구가 25.2%로 모두베기구 17.0%에 비해 높았다(Table 2). 모두베기구의 점토 함량이 낮게 나타난 것은 상층 임관의 제거와 낙엽·낙지 유입량 감소에 따른 광물질 토층의 노출과 강우 발생 시 광물질 표토층으로부터 점토 같은 세립질 입자의 세탈이나 용탈이 발생하였거나, 인접한 소규모 임분 단위에서도 공간적 변동이 크게 나타났기 때문으로 사료된다. 전질소, 유기탄소, 인 등은 모두베기구가 수하식재구에 비해 낮은 농도를 보이나 처리 간 유의적인 차이는 없었다. 그러나 교환성 마그네슘 함량은 수하식재구에 비해 모두베기구가 유의적으로 낮았으며, 이는 양이온의 흡착 능력이 높은 점토나 유기탄소 농도가 낮게 나타났기 때문이다. 모두베기구의 토양 유기탄소와 전질소 농도는 3.10%와 0.24%로 우리나라 남부지역 편백림의 평균 유기탄소 농도 7.2%나 전질소 농도 0.41%에 비해 낮았다(Kim et al., 2020).
Table 2.
Soil physical and chemical properties in under-planted (DR) and clearcut (CR) regeneration treatments of Chamaecyparis obtusa.
|
Treatment |
Sand (%) |
Silt (%) |
Clay (%) |
N (%) |
C (%) |
C/N |
P (mg kg−1) |
K+
|
Ca2+
|
Mg2+
|
|
(cmolc kg−1) |
|
DR |
17.8 (4.3)b |
57.0 (2.7)a |
25.2 (1.6)a |
0.28 (0.02)a |
3.52 (0.28)a |
12.5 (0.32)a |
2.56 (0.83)a |
0.35 (0.03)a |
1.00 (0.50)a |
1.57 (0.21)a |
|
CR |
33.5 (1.9)a |
49.5 (1.7)a |
17.0 (0.4)b |
0.24 (0.02)a |
3.10 (0.55)a |
12.3 (1.24)a |
1.67 (0.62)a |
0.37 (0.02)a |
1.03 (0.43)a |
0.80 (0.08)b |
Download Excel Table
2. 낙엽의 탄소 및 질소 농도
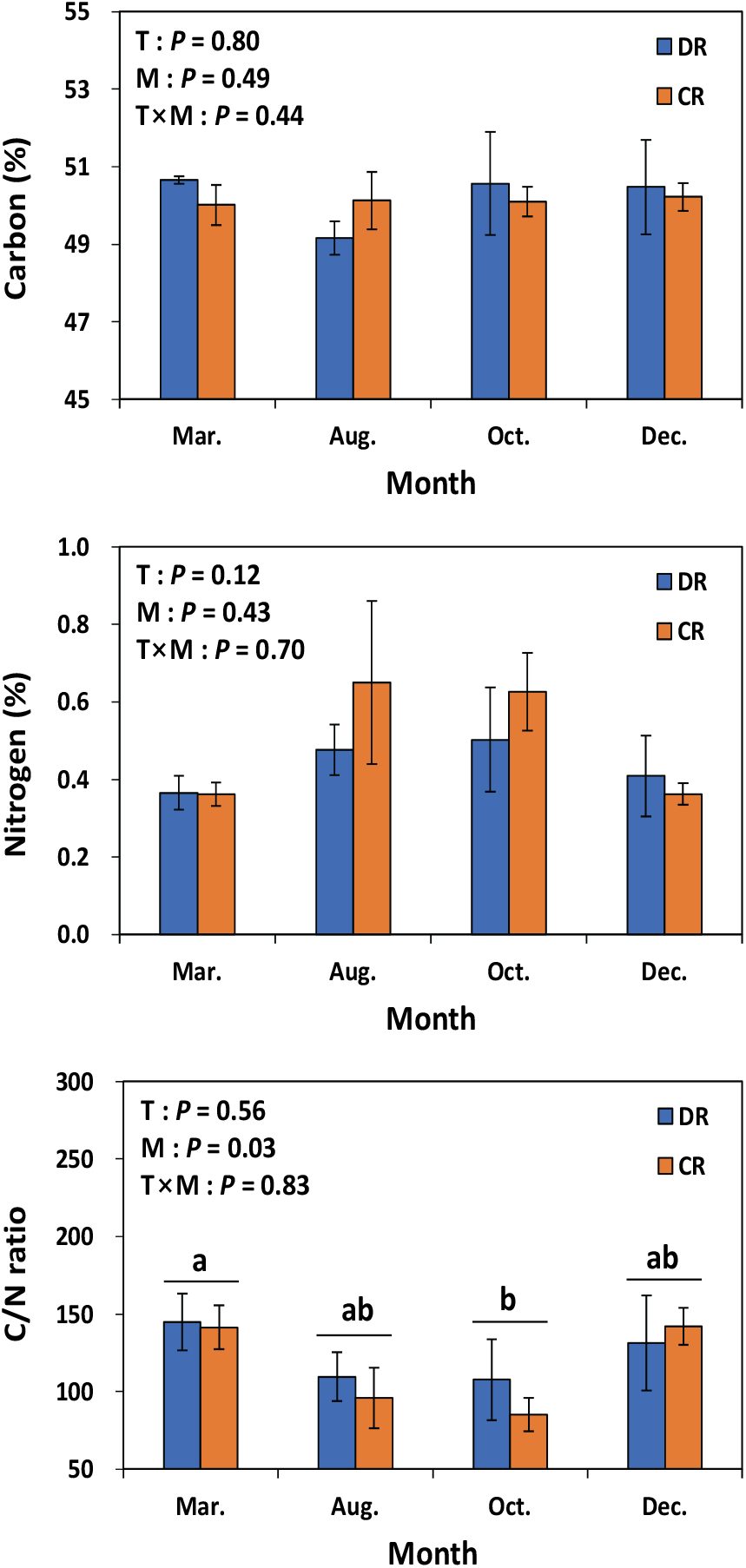
편백 낙엽의 평균 탄소 농도는 모두베기구와 수하식재구 모두 50.1%로 유의적인 차이는 없었다(Figure 3). 이는 수집된 편백 낙엽의 탄소 농도가 광, 양분환경, 낙엽 유입 시기 같은 시·공간적 요인의 변화로부터 영향이 크지 않았음을 시사한다. 일본에서 조사된 카나가와현 편백 40년생의 11월에 수집된 낙엽의 탄소 농도는 50.2%(Nakajima and Kaneko, 2012)로 본 조사구 결과와 차이는 없었다. 그러나 8월~10월에 유입된 낙엽의 C/N비는 93으로 12월~3월의 C/N비 143보다 유의적으로 낮았다. 편백 낙엽의 탄소 농도가 수집 시기별로 유의적인 차이가 없음을 고려할 때, 8월~10월의 낮은 C/N비는 이 시기 동안 높은 양분 농도를 보이는 녹엽(green leaves) 유입이 원인으로 사료된다. 유사한 결과로 일본 카나가와현의 편백 조림지 낙엽의 C/N비는 11월에 63으로 8월의 39에 비해 높은 값을 보였으며, 11월의 높은 C/N비는 낙엽 내 질소 농도 감소가 원인으로 제시된 바 있다(Nakajima and Kaneko, 2012).
Figure 3.
Carbon concentration, nitrogen concentration and C/N ratio of leaf litter of Chamaecyparis obtusa in under-planted (DR) and clearcut (CR) regeneration treatments. T: treatment method; M: month. Vertical bars indicate standard error. Different letters on the bars represent a significant difference at P < 0.05.
Download Original Figure
편백 낙엽 내 질소 농도의 경우 모두베기구나 수하식재구 사이에 유의적인 차이는 없었지만, 임목 생육 기간 동안(8월과 10월) 채취한 낙엽의 경우 모두베기구에서 채취한 낙엽의 질소 농도가 높은 경향을 보였다. 유사한 결과로 Nakanishi et al. (2009)는 모두베기 후 토양 질소유효도의 증가에 따라 편백 낙엽 내 질소 농도가 증가하는 것을 관찰한 바 있다. 본 연구에서 12월에 채취한 녹엽의 질소 농도는 모두베기구 1.10%, 수하식재구 1.15%, 낙엽의 질소 농도는 모두베기구 0.35%, 수하식재구 0.40%로서, 일본 교토지역 편백 녹엽의 질소 농도 0.86%~1.23%의 범위에 분포하였으나, 낙엽의 질소 농도 0.47%~0.71%에 비해서는(Nakanishi et al., 2009) 낮아 토양 양분환경이 열악한 이 지역의 편백 조림지는 양분 재전류(nutrient retranslocation)가 큰 것으로 나타났다.
3. 낙엽·낙지 부위별 탄소 및 질소 농도
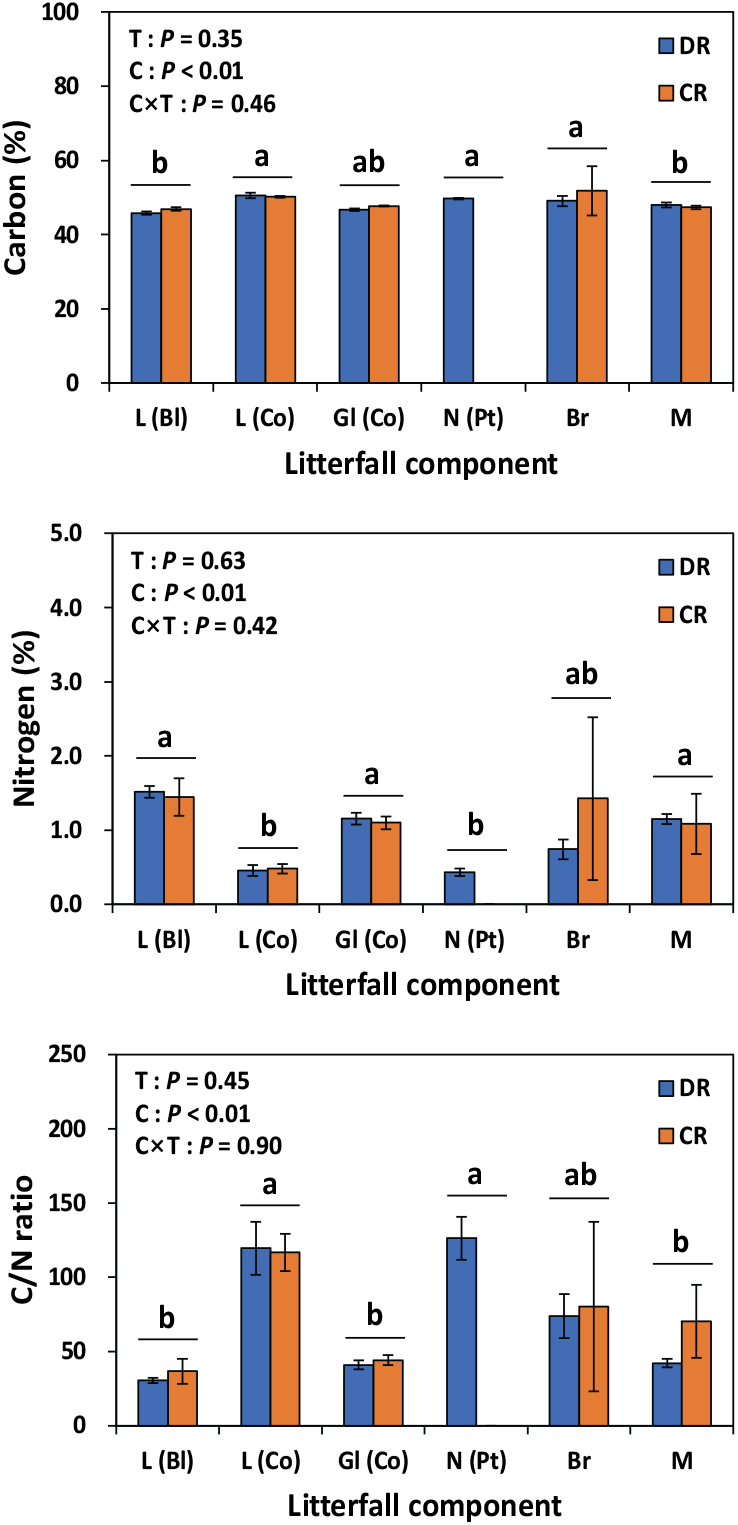
낙엽·낙지의 탄소 농도는 가지, 편백 낙엽, 소나무 낙엽 등의 탄소 농도가 활엽수나 기타 물질에 비해 유의적으로 높았다(Figure 4). 임목의 부위별 탄소 농도는 질소 농도와 부의 상관이 있는 것으로 알려져 있으며(Kim et al., 2017), 본 연구에서도 질소 농도가 낮은 곰솔 잎이나 가지 등의 탄소 농도가 잎과 같은 타 부위에 높은 경향을 보였다. 낙엽·낙지의 질소 농도는 활엽수 잎, 편백 녹엽, 기타 물질 등이 편백 낙엽이나 가지 등에 비해 높은 농도를 보였다. 그 결과 낙엽·낙지 부위별 C/N비는 곰솔 낙엽이나 편백 낙엽 등이 기타 물질, 편백 녹엽, 활엽수 낙엽 등에 비해 유의적으로 낮은 C/N비를 보였다.
Figure 4.
Carbon concentration, nitrogen concentration and C/N ratio of litterfall components [L (Bl): broadleaved leaf litter; L (Co): leaf litter of Chamaecyparis obtusa; Gl (Co): green leave of Chamaecyparis obtusa; N (Pt): needle litter of Pinus thunbergii; Br: branch litter; M: miscellaneous litter] litter of Chamaecyparis obtusa in under-planted (DR) and clearcut (CR) regeneration treatments. T: treatment method; C: litterfall component. Vertical bars indicate standard error. Different letters on the bars represent a significant difference at P < 0.05.
Download Original Figure
4. 낙엽에 의한 유기탄소 및 질소 유입량
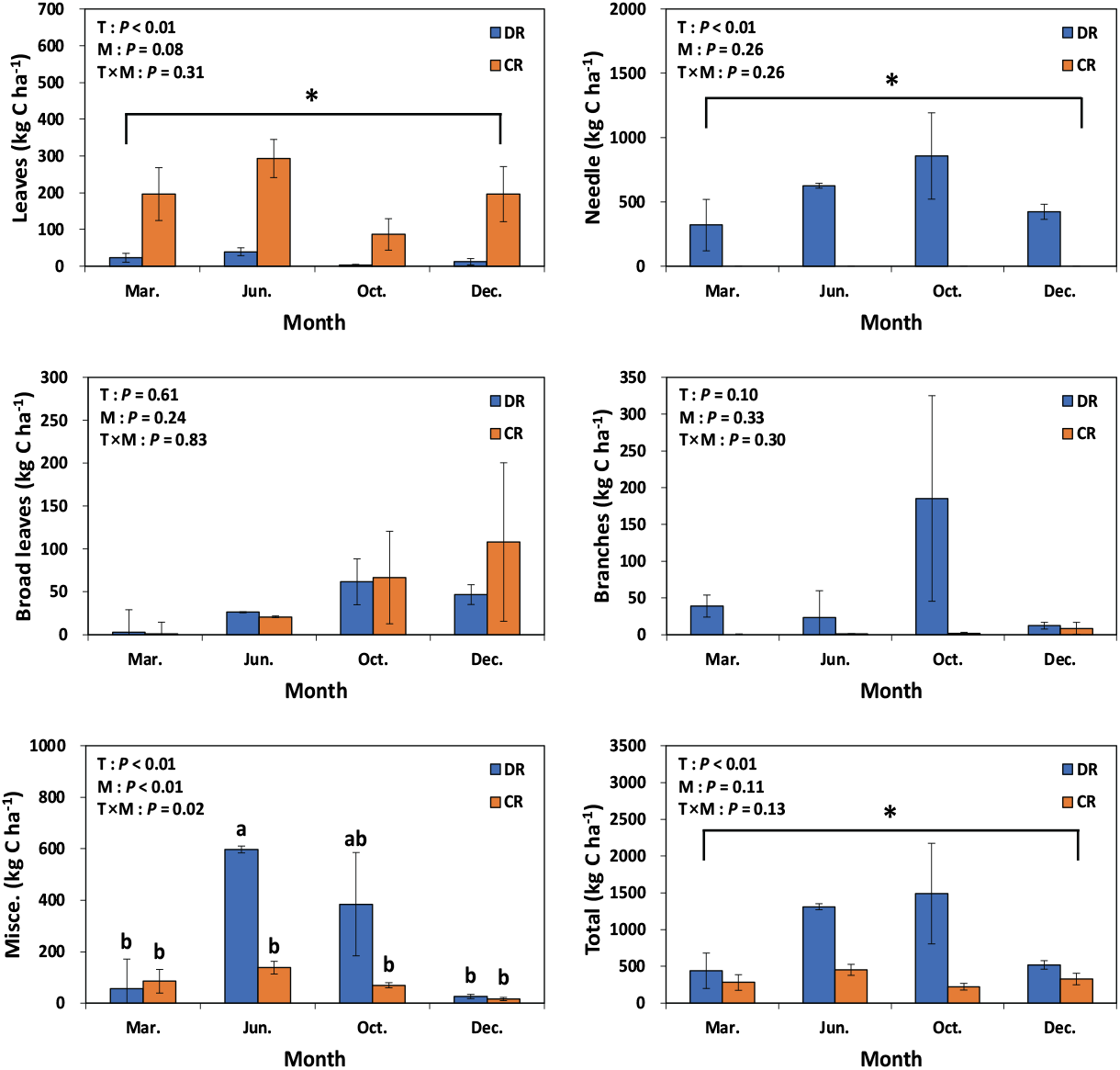
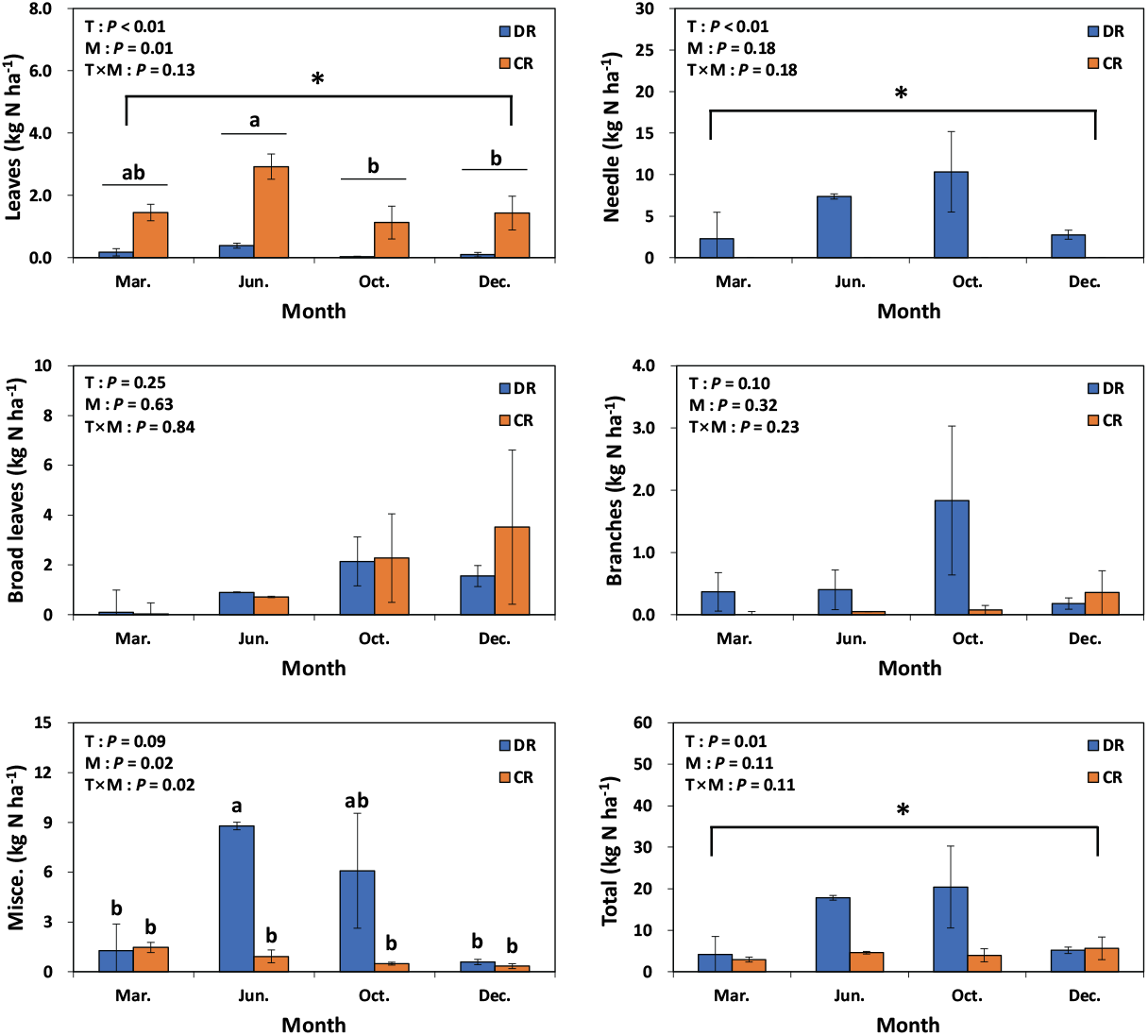
편백 낙엽에 의한 탄소 및 질소 유입량은 모두베기구나 수하식재구 모두 3월~6월이 가장 많은 유입량을 보인 반면에 6월~10월은 가장 적게 유입되었다(Figure 5, 6). 본 연구결과는 일본 아이치현에서 조사된 25년생 편백 조림지의 낙엽이 가을과 2월에 가장 많은 양이 유입된 결과(Hagihara et al., 1978; Inagaki et al., 2008)와는 상이하며, 이는 일본 편백 임분의 경우 강설량이 낙엽 유입량에 영향을 끼친 반면에, 본 조사구는 겨울 동안 강설이 발생하지 않는 온난한 지역이기 때문으로 사료된다. 주로 중층과 하층에 분포하는 낙엽활엽수의 낙엽 유입량은 모두베기구와 수하식재구 사이에 유의적인 차이가 없었으며, 10월~12월 사이에 가장 많은 양이 유입되어, 국내에서 알려진 남부지역 낙엽활엽수의 heavy litterfall 시기(Kim et al., 2013a)와 일치하였다. 가지에 의한 탄소 및 질소 유입량은 수하식재구의 경우 장마 기간이나 태풍이 주로 많이 발생하는 시기인 6월~10월에 가장 많이 유입되었고, 모두베기구는 12월에 유입량이 많았다. 주로 생식기관으로 구성되는 기타물질에 의한 탄소 및 질소 유입량은 모두베기구나 수하식재구 모두 6월과 10월에 가장 많이 유입되었으며, 6월의 경우 곰솔 숫꽃이나 편백 숫꽃 같은 생식기관이 많이 유입되었고 10월은 구과(cone)가 많았다. 본 연구 결과와 대조적으로 일본 편백 조림지에 기타물질의 최대 유입 시기는 곤충배설물의 유입이 주로 발생하는 여름이었다(Miyaura and Hozumi, 1985). 수하식재구의 곰솔 낙엽·낙지에 의한 탄소 및 질소 유입량은 침엽수 임분의 heavy litterfall 시기인 10월이 가장 많았고(Berg and Laskowski, 2006), 6월, 12월, 3월 순으로 편백 낙엽 유입의 계절적 변동과는 다른 경향을 보였다.
Figure 5.
Seasonal carbon inputs of litterfall components in under-planted (DR) and clearcut (CR) regeneration treatments. T: treatment method; M: month. Leaf litter of Chamaecyparis obtusa (leaves) and needle litter (needle) of Pinus thunbergii. Vertical bars indicate standard error. Different letters and asterisk on the bars represent a significant difference at P < 0.05.
Download Original Figure
Figure 6.
Seasonal nitrogen inputs of litterfall components in under-planted (DR) and clearcut (CR) regeneration treatments. T: treatment method; M: month. Leaf litter of Chamaecyparis obtusa (leaves) and needle litter (needle) of Pinus thunbergii. Vertical bars indicate standard error. Different letters and asterisk on the bars represent a significant difference at P < 0.05.
Download Original Figure
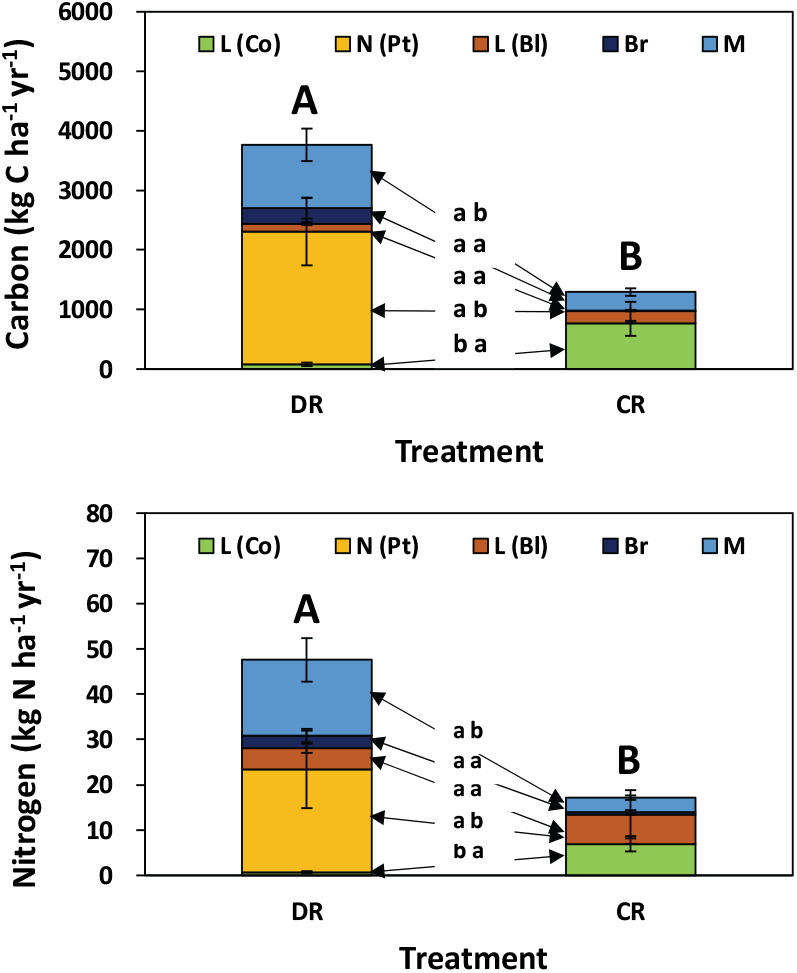
5. 총 탄소 및 질소 유입량
편백 낙엽·낙지에 의한 탄소 및 질소 유입량은 모두베기구가 773 kg C ha−1 yr−1, 6.95 kg N ha−1 yr−1로 수하식재구 78 kg C ha−1 yr−1, 0.70 kg N ha−1 yr−1에 비해 유의적으로 많은 양이 유입되었다(Figure 7). 이는 모두베기구의 경우 임분 밀도가 낮고 임목 사이에 충분한 공간으로 인하여 수관이 하부까지 잘 발달한 반면에 수하식재구는 광 및 토양 수분 조건 등의 차이가 원인으로 수관 발달이 빈약하였기 때문이다(Figure 1). 중층이나 하층에 분포하는 낙엽활엽수에 의한 낙엽량은 모두베기구가 많을 것으로 예측되었으나 수하식재구와 차이가 없었으며, 이는 모두베기구를 대상으로 전년도(2018)에 어린나무 가꾸기가 실시되어, 중·하층의 활엽수류가 대부분 제거되었기 때문이다. 가지의 탄소 및 질소 유입량은 수하식재구 260 kg C ha−1 yr−1과 2.8 kg N ha−1 yr−1로 모두베기구 11.6 kg C ha−1 yr−1과 0.48 kg N ha−1 yr−1에 비해 크게 나타났지만 유의적인 차이는 없었다. 기타 물질은 수하식재구 1,063 kg C ha−1 yr−1, 16.7 kg N ha−1 yr−1, 모두베기구 310 kg C ha−1 yr−1, 3.2 kg N ha−1 yr−1로 수하식재구가 높았으며, 이는 곰솔의 숫꽃 같은 생식기관의 유입이 많았기 때문이다. 총 탄소 및 질소 유입량은 수하식재구 3,765 kg C ha−1 yr−1, 47.6 kg N ha−1 yr−1, 모두베기구 1,290 kg C ha−1 yr−1, 17.2 kg N ha−1 yr−1로 수하식재구가 유의적으로 크게 나타났다. 타 연구에서 조사된 결과로 일본의 25년생 편백 임분은 낙엽 900 kg C ha−1 yr−1, 가지 100 kg C ha−1 yr−1, 기타 물질 250 kg C ha−1 yr−1, 총 낙엽량 1,100 kg C ha−1 yr−1(Miyaura and Hozumi, 1985.)로 본 조사구의 모두베기구와 큰 차이는 없었다. 일본 아이치현 18년생 편백 조림지의 경우도 총 낙엽·낙지량은 883 kg C ha−1 yr−1로, 이 중 편백 낙엽이 729 kg C ha−1 yr−1, 가지 49 kg C ha−1 yr−1, 수피, 생식기관, 곤충배설물들과 같은 기타 물질은 13 kg C ha−1 yr−1 정도(Hagihara et al., 1978)로, 모두베기구의 편백 낙엽이나 가지 유입 및 총 낙엽·낙지량과 유사하였다. 본 연구의 모두베기구 편백 낙엽의 질소 유입량은 6.95 kg N ha−1 yr−1로 일본 시코쿠지역 17년생 편백 조림지의 편백 잎에 의한 질소 유입량 10.2 kg N ha−1 yr−1~50 kg N ha−1 yr−1(Inagaki et al., 2004)에 비해 낮게 나타났다. 편백 낙엽의 질소 농도는 간벌(Inagaki et al., 2004), 연년 변화(Inagaki et al., 2008), 기상 요인(Inagaki et al., 2010), 해발고나 경사(Nakanishi et al., 2009) 등의 다양한 요인에 의해 영향을 받는 것으로 알려져 있으며, 본 조사구의 경우 토양 내 양분환경이 낮게 나타나는 것도 하나의 원인으로 사료된다. 한편, 모두베기구에서 가지량이 낮은 것은 유령림 단계에 임관 하부에 죽은 가지가 거의 나타나지 않고 자연낙지가 잘 발생하지 않았기 때문이다(Hagihara et al., 1978). 수하식재구의 상층 곰솔 낙엽은 2,227 kg C ha−1 yr−1과 78 kg N ha−1 yr−1로 편백 낙엽·낙지의 유기탄소 유입량에 비해 약 28배 정도 많았으며, 모두베기구에 비해 약 3배 이상 많이 유입되어, 편백 조림지의 상층 임관 제거로부터 발생할 수 있는 낙엽·낙지량의 급격한 감소에 완충 역할을 하고 있는 것으로 나타났다.
Figure 7.
Total carbon and nitrogen inputs of litterfall components [L (Bl): broadleaved leaf litter; L (Co): leaf litter of Chamaecyparis obtusa; Gl (Co): green leave of Chamaecyparis obtusa; N (Pt): needle litter of Pinus thunbergii; Br: branch litter; M: miscellaneous litter] in under-planted (DR) and clearcut (CR) regeneration treatments. Vertical bars indicate standard error. Different letters between treatments represent a significant difference at P < 0.05. T: treatment method; M: month.
Download Original Figure
결 론
소나무재선충병 피해지에 모두베기나 미피해목 일부를 잔존하고 편백을 수하식재 한 15년생 조림지의 평균 흉고직경 생장과 낙엽·낙지에 의한 탄소 및 질소 유입량은 갱신방법에 따라 차이가 있었다. 편백의 평균 흉고직경 생장은 모두베기구가 수하식재구에 비해 유의적으로 크게 나타나 양호한 직경생장 특성을 보였다. 그러나 편백 낙엽의 탄소 및 질소 농도는 갱신방법이나 수집 시기에 따라 유의적인 차이는 없었으나, 낙엽·낙지 부위별 탄소 및 질소 농도는 유의적인 차이가 있었다. 편백 낙엽에 의한 탄소 및 질소 유입량은 모두베기구가 수하식재구에 비해 유의적으로 높았으나, 총 낙엽·낙지에 의한 탄소 및 질소 유입량은 수하식재구에 비해 약 3배 정도 적은 양이 유입되어, 모두베기구의 임상 및 토양층의 유기물 감소가 상당 기간 지속 될 것으로 예측되었다.
감사의 글
본 논문은 정부(교육부, 과학기술정보통신부)의 재원으로 한국연구재단의 지원을 받아 수행된 연구(No. 2017R1 D1A3B03029826; No. 2020R1A2C1005791)입니다.
References
Baek, G. 2020. Soil CO
2 Efflux Dynamics in Fertilized
Pinus densiflora and
Querus variabilis Stands, and in Restored Pine Wilt Disease-disturbed Forests. A Ph.D. Dissertation of Gyeongnam National University of Science and Technology. pp. 96.

Berg, B. and Laskowski, R. 2006. Litter decomposition; A guide to carbon and nutrient turnover. Advance of Ecological Research 38: 20-71.
Diaci, J., Rozenbergar, D., Fidej, G. and Nagel, T.A. 2017. Challenges for uneven-aged silviculture in restoration of post-disturbance forests in central Europe: A synthesis. Forests 8(10): 378.

Hagihara, A. Suzuki, M. and Hozumi, K. 1978. Seasonal fluctuations of litter fall in a
Chamaecypais obtusa plantation. Journal of Japanese Forestry Society 60(11): 397-404.
Hirata, R., Ito, S., Eto, K., Sakuta, K., Mizoue, S. and Mitsuda, Y. 2015. Early growth of hinoki (
Chamaecyparis obtusa) trees under different and edge aspect at a stip-clearcut site in Kyushu, Southern Japan. Journal of Forest Research 20(6): 522-529.
Inagaki, Y, Miura, S. and Kohzu, A. 2004. Effects of forest type and stand age on litterfall quality and soil N dynamics in Shikoku district, southern Japan. Forest Ecology and Management 202(1-3): 107-117.
Inagaki, Y., Sakai, A., Kuramoto, S., Kodani, E., Yamada, T. and Kawasaki, T. 2008. Inter-annual variations of leaf-fall phenology and leaf-litter nitrogen concentration in a hinoki cypress (
Chamaecyparis obtusa Endlicher) stand. Ecological Research 23: 965-972.
Inagaki, Y., Okuda, S., Sakai, A., Nakanishi, A., Shibata, S. and Fukuta, H. 2010. Leaf-litter nitrogen concentration in hinoki cypress forests in relation to the time of leaf fall under different climatic conditions in Japan. Ecological Research 25(2): 429-438.
Jeon, K.S., Kim, C.S., Park, N.C., Hur, T.C. and Hong, S.C. 2011. Effects on control of pine wilt disease (
Bursaphelenchus xylophilus) by thinning methods in red pine (
Pinus densiflora) forest. Journal of Korean Forest Society 100(2): 165-171.
Kalra, Y.P. and Maynard, D.G. 1991. Methods Manual for Forest Soil and Plant Analysis. Forestry Canada, Northwest Region, Northern Forestry Centre, Edmonton, Alberta. Information Report NOR-X-319E. pp. 116.
Kim, C., Sharik, T.L. and Jurgensen, M.F. 1996. Litterfall, nitrogen, and phosphorus inputs at various levels of canopy removal in oak and pine stands. American Midland Naturalist 135(2): 195-204.
Kim, C., Jang, K.S., Kim, J.B., Byun, J.K., Lee, C.H. and Jeon, K.S. 2010. Relationship between soil properties and incidence of pine wilt disease at stand level. Landscape and Ecological Engineering 6: 119-124.
Kim, C., Byun, J.K., Park, J.H. and Ma, H.S. 2013a. Litter fall and nutrient status of green leaves and leaf litter at various compound ratios of fertilizer in saw-tooth oak stands. Annals of Forest Research 56(2): 339-350.
Kim, C., Lee, K.S., Son, Y.M. and Cho, H.S. 2013b. Allometric equations and biomass expansion factors in an age-sequence of black pine (
Pinus thunbergii) stands. Journal of Korean Forest Society 102(4): 543-549.
Kim, C., Lim, J.H., Lee, I.K., Park, B.B. and Chun, J.H. 2013c. Annual variation of litterfall production in a broadleaved deciduous forest at the Mt. Keumsan LTER site. Journal of Korean Forest Society 102(2): 210-215.
Kim, C., Yoo, B.O., Jung, S.Y. and Lee, K.S. 2017. Allometric equations to assess biomass, carbon and nitrogen content of black pine and red pine trees in southern Korea. iForest-Biogeoscience and Forestry 10(2): 483-490.
Kim, C., Baek, G., Choi, B., Ha, J., Bae, E.J., Lee, K.S. and Son, Y.M. 2020. Carbon stocks of tree, forest floor, and mineral soil in
Cryptomeria japonica and
Chamaecyparis obtusa stands. Journal of Korean Society of Forest Science 109(2): 169-178.
Kim, S.K., Bae, S.W., Lee, K.S. and Lee, K.J. 2004. Silvicultural strategy to establish
Chamaecyparis obtusa and
Cryptomeria japonica plantations. Annual Proceeding of Korean Forest Society 489-494.
Lehtonen, A., Lindholm, M., Hokkanen, T., Salminen, H. and Jalkanen, R. 2008. Testing dependence between growth and needle litterfall in Scots pine - a case study in northern Finland. Tree Physiology 28(11): 1741-1749.
Miyaura, T. and Hozumi, K. 1985. Measurement of litterfall in a hinoki (
Chamaecyparis obtusa S. et. Z.) plantation by the clothing-trap method. Journal of Japanese Forestry Society 67(7): 271-277.
Morimoto, J. and Yoshida, H. 2005. Dynamic changes of native Rhododendron colonies in the urban fringe of Kyoto city in Japan: detecting the long-term dynamism for conservation of secondary nature. Landscape and Urban Planning 70(3-4): 195-204.
Nakanishi, A., Inagaki, Y., Osawa, N., Shibata, S. and Hirata, K. 2009. Effects of patch cutting on leaf nitrogen nutrition in hinoki cypress (
Chamaecyparis obtusa Endlicher) at different elevations along a slope in Japan. Journal of Forest Research 14(6): 388-393.
Nakajima, G. and Kaneko, N. 2012. The effect of Japanese cypress plantation on soil biochemical characteristics converted from deciduous forests. Journal of Japanese Forest Society 94(3): 112-119.
Nolet, P., Kneeshaw, D., Messier, C. and Béland, M. 2018. Comparing the effects of even- and uneven-aged silviculture on ecological diversity and processes: A review. Ecology and Evolution 8(2): 1217-1226.
O’Hara, K.L. and Ramage, B.S. 2013. Silviculture in an uncertain world:utilizing multi-aged management systems to integrate disturbance. Forestry 86(4): 401-410.
Paquette, A., Bouchard, A. and Cogliastro, A. 2006. Survival and growth of under-planted trees: A meta-analysis across four biomes. Ecological Applications 16(4): 1575-1589.
Park, I.H., Lim, D.H. and Ryu, S.B. 2000. Biomass, net production and nutrient distribution related to age of young
Chamaecyparis obtusa plantations. Journal of Korean Forestry Society 89(1): 85-92.
SAS Institute Inc. 2003. SAS/STAT Statistical Software. Version 9.1 SAS Publishing Cary, NC.