Article
시비처리에 따른 상동나무 용기묘의 생장 및 생리특성
어현지1, 손용환1, 박성혁1, 박광훈1, 이경철2, 손호준1,*
Growth and Physiological Characteristics of Containerized Seedlings of Sageretia thea at Different Fertilization Treatments
Hyun Ji Eo1, Yong Hwan Son1, Sung Hyuk Park1, Gwang Hun Park1, Kyeong Cheol Lee2, Ho Jun Son1,*
1Forest Medicinal Resources Research Center, National Institute of Forest Science, Yeongju 36040, Korea
2Department of Forestry, Korea National College of Agriculture and Fisheries, Jeonju 54874, Korea
© Copyright 2021 Korean Society of Forest Science. This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received: Apr 05, 2021; Revised: May 28, 2021; Accepted: May 28, 2021
Published Online: Jun 30, 2021
요약
본 연구는 다양한 효능·효과가 최근에 입증된 상동나무를 대상으로 다목적 수용성 복합비료를 농도(0.5 g·L−1, 1.0 g·L−1, 1.5 g·L−1, 2.0 g·L−1)처리에 따른 생장특성(간장, 근원경 및 H/D율)과 생리특성(광합성, 엽록소 함량 및 형광반응)을 정량화하여 적정 시비 수준을 알아보고자 실시하였다. 간장생장은 시비농도가 1.5 g·L−1까지 높아짐에 따라 생장량이 증가했으나 2.0g·L−1 시비처리에서는 낮아졌다. 근원경 생장은 시비농도가 높아짐에 따라 생장량은 유의하게 낮아졌다. 광합성 반응은 1.5 g·L−1 시비처리에서 엽육세포 내 CO2 반응곡선, 최대광합성효율, 최대 카르복실화 속도가 가장 높았다. 엽록소 형광반응과 엽록소 지수는 무처리구에 비해 시비처리구에서 유의적으로 광합성 기구의 효율을 높이고 있음을 확인하였다. 따라서 우량한 묘목품질의 상동나무를 생산하기 위한 시비 수준은 1.0~1.5 g·L−1 농도라고 판단된다.
Abstract
This study aims to optimize the appropriate concentration of fertilizers for Sageretia thea by analyzing growth performances (height and root collar diameter) and physiological characteristics (photosynthesis, chlorophyll contents, and chlorophyll fluorescence reaction). As fertilizer concentration was increased to 1.5 g·L−1, growth increased, but it decreased at 2.0 g·L−1 treatment. Root collar diameter growth was reduced because of higher fertilizer concentrations. Photosynthesis reactions showed the highest CO2 reaction curves, maximum photosynthesis rate, and maximum carboxylation rate in the 1.5 g·L−1 fertilizer treatment. The chlorophyll fluorescence reaction and SPAD values revealed that fertilizer treatment improves photosynthesis efficiency and robustness compared with untreated control. Therefore, the appropriate fertilizer concentration for producing good seedling quality of Sageretia thea is 1.0~1.5 g·L−1.
Keywords: Sageratia thea; physiological characteristics; growth performances; medicinal plants
서 론
우리나라에서 산림자원은 예로부터 향촌의 생활을 영위하는 용도나 구황작물, 한약재로서 다양하게 이용되어 왔는데, 최근 건강에 대한 지대한 관심과 함께 약효와 특유의 맛, 식생활 습관, 계절감 등을 이유로 수요가 크게 증가하고 있다(Cho, 2000). 이러한 식⋅약용식물은 최근 생명공학기술이 발달함에 따라 경험적으로 판단해왔던 효능이 과학적으로 가치평가 되고 있으며, 이를 활용해 과거 단순 식용으로서의 기능은 이제 천연물신약, 기능성식품 및 화장품의 원료로서 광범위하게 활용되고 있는 상황이다(Song et al., 2020). 동시에 1993년 생물다양성협약(Convention on Biological Diversity, CBD)이 발효됨에 따라 국제적으로 자생식물의 주권에 대한 관심이 여느 때보다 높아지면서 국가 간 생물자원의 주권 확보 경쟁으로 이어지고 있다. 따라서 향후 있을 특허권 경쟁에서 자생 생물자원에 대한 명백한 근거를 확보하는 것이 중요하다(Kim et al., 2004).
상동나무는 갈매나무과(Rhamnaceae), 상동나무속(Sageretia)의 식물로 아시아의 동⋅남부 지역, 북아메리카 남부 지역에 분포하며, 우리나라에는 상동나무속 상동나무(Sageretia thea) 1속 1종이 제주도와 남해안의 해안가 및 산지에 분포하고 있다(Choo, 1992). 상동나무는 2m 정도의 반상록 성 활엽관목으로 가을에 꽃이 피고, 잎이 나는 봄에 열매가 흑색으로 익는다(Kim and Kim, 2011). 이러한 열매는 제주도, 전라도에서 식용하거나, 잎과 잔가지를 활용하면 옻독을 치료하는데 쓰인다고 하며(Chung et al., 2017), 감기, 발열, 간염 및 피부염 치료 등으로 사용되어 왔다(Kim and Hyun, 2015).
상동나무에 관한 기초적인 연구로는 종자발아 특성, 제주도 내 자생지 생육환경에 관한 특성 조사가 이루어졌으며(Song et al, 2014, 2015a, 2015b), 기능성 연구에서는 가지와 잎 추출물의 항염증 활성(Kim et al., 2019c)과 대장암, 폐암에 대한 세포사멸을 유도하는 효과 등이 보고되었다(Kim et al., 2019a, 2019b, 2020). 이 외에도 열매 추출물의 항산화 활성, 미백활성, 암세포 생장 억제 활성 등의 연구가 활발하게 진행되고 있다(Pyo et al., 2020). 하지만 상동나무의 대량생산에 관한 증식 및 재배학적 연구는 미흡한 실정이다.
본 연구는 식⋅약용 자원으로서 활용 가치가 높은 상동나무를 대량생산 및 산업화하기 위한 연구로, 시설양묘과정에서 미치는 다양한 생육환경 조건과 용기, 상토, 시비기술 등이 규명돼야한다(Grossniiclke, 2005). 그 중 시비는 용기묘의 특성상 반드시 실시되어야 하며(Broschat, 1995), 수종별 양분요구도에 맞는 시비과정을 통해 우량 묘목의 생산과 함께 시설양묘장 주변의 토양 및 계류수의 환경적인 문제를 저감할 수 있다(Bumgarner et al., 2008). 따라서 우량묘목 생산을 위한 상동나무의 시비별 생장 및 생리적 특성을 분석하여 시설양묘에 가장 적절한 시비 수준의 규격화를 통해 우량한 묘목의 생산은 물론 친환경이고 경제적인 개발에 도움을 줄 것으로 기대된다.
재료 및 방법
1. 공시재료 및 시비처리
본 연구에서 사용된 상동나무(Sageretia thea)는 2018년 5월 국립산림과학원 난대아열대산림연구소 연구시험림인 저지곶자왈에서 종자를 채취하여 육묘상자에 파종하였으며, 파종상은 발아시 까지 충분한 수분공급을 위해 일 1회 m2 당 20 L 관수를 실시하였다. 발아한 개체의 육묘는 온실에서 주 2~3회 m2당 20 L 관수를 실시했으며, 이후 2019년 3월 초 개체 중 형질이 고르고 우수한 1년생 실생묘를 240 mL 양묘용기에 이식하였다. 이식 후 활착을 확인하고 생육상태가 양호한 개체를 선별하여 처리구당 30본씩 총 150본을 사용하였다.
실험은 국립산림과학원 산림약용자원연구소의 시설온실에서 실시하였으며, 설정온도를 최고 25 ± 5℃, 최저 18 ± 5℃로 유지하고 고온 시 환기팬을 가동하여 적정온도를 유지하였다.
상토는 피트모스:펄라이트:질석=1:1:1(v/v)의 배합토를 이용하였고(Table 1), 관수는 이식 후, 주 2~3회 m2당 20 ℓ 관수를 실시하였다.
Table 1.
Physical and chemical properties of media used in container seedling production.
|
Soil media mixture (v/v) |
pH |
EC (ds·m−1) |
NO− (mg/kg−1) |
P2O5 (mg/kg−1) |
Ex-cations(cmol·kg−1) |
C.E.C. (cmol ·kg−1) |
|
Peat moss |
Perlite |
Vermiculite |
K+
|
Ca2+
|
Mg2+
|
|
1 |
1 |
1 |
6.0 |
0.1 |
0.1 |
123 |
4 |
10 |
7 |
17 |
Download Excel Table
시비는 2019년 6월 10일부터 2019년 8월 26일까지 약 12주간 주 1회씩 실시하였으며, 임업시설양묘에서 주로 사용되고 있는 수용성비료인 Multi Feed 19(N:P:K=19:19:19, Haifa chemicals, Israel)를 이용하여, 대조구(무처리)와 처리구인 0.5 g·L−1, 1.0 g·L−1, 1.5 g·L−1 및 2.0 g·L−1 등의 5가지 수준으로 시비를 실시하였다.
관수와 시비는 고온 및 햇빛에 의한 피해를 줄이기 위하여 오전 8~10시 또는 오후 4~6시 사이에 실시하였다.
2. 생장 특성 조사
상동나무의 생장 반응을 조사하기 위해 처리구 트레이별 10개씩 3반복으로 간장과 근원경을 반복 측정하여 생장량의 평균을 산출하였다. 측정은 시비처리 기간에 주 1회 측정하였으며, 시비처리 후 생장량 변화를 확인하기 위하여 3주간 추가 조사하여 총 15회 측정하였다. 측정된 생장량은 특성을 분석하기 위해 시비시점을 기준으로 상대생장량을 산출하였으며, 묘목의 건전도를 확인할 수 있는 H/D율 [SQ; Sturdiness Quotient = height (cm) / root collar diameter (mm)]을 계산하였다(Bayala et al., 2009). 간장의 측정에는 줄자(KOMERON, KMC-32D)를 근원경의 측정에는 디지털버니어 캘리퍼스(Cas, DC-200-1)를 이용하였다.
3. 광합성 특성
시비처리에 따른 광합성 반응은 8월 28일 휴대용 광합성 측정기(Portable Photosynthesis system, Li-6800, Li-Cor Inc., USA)를 이용하여 측정하였으며, CO2 농도(0 ~ 1400 μmol·mol−1)에 의한 엽육세포내 CO2 반응 곡선(PN-Ci)을 작성하고 최대 카르복실화 속도(Maximum carboxylation rate, Vcmax), 최대 전자전달 속도(Maximum electron transport rate, Jmax) 및 호흡율(dark respiration rate, RD)을 구하였다(Sharkey et al., 2007).
또한 광합성 측정장치에 부착된 LED light source를 이용하여 PPFD (Photosynthetic Photon Flux Density)를 1,000 μmol·m−2·s−1로 고정하고 최대광합성속도(maximum photosynthesis rate; PNmax), 기공증산속도(stomatal transpiration rate; E), 기공전도도(stomatal conductance; Gs) 및 엽육세포 내 CO2 농도(intercellular CO2 concentration, Ci)의 변화를 조사하였으며, 이때 공통된 측정 조건은 챔버로의 유입 공기유량을 600 μmol·s−1, 온도를 25 ± 2℃로 유지하였다. 측정된 결과를 통해 엽육세포 내의 CO2 농도의 비율인 CiㆍCa−1와 수분이용효율을 나타냈으며, 수분이용효율(water use efficiency)은 아래의 식을 이용해 내재적 수분이용효율(intrinsic water use efficiency, WUEi)과 순간증산효율(instantaneous transpiration efficiency, ITE)로 구분하여 나타내었다(Oh and Koh, 2017).
4. 엽록소 함량 및 엽록소 형광반응 분석
휴대용 엽록소 형광반응 측정기(Plant Efficiency Analyzer, Hansatech Instrument Ltd., King’s Lynn, UK)를 이용하여 20분간 암적응 시킨 엽에 3,500 μmol·m−2·s−1의 광량을 1초간 조사하고, Fo/Fm, Fv/Fm, Fv/Fo 및 PIabs 등의 엽록소 형광반응을 조사하였다(Strasser et al., 2000; Wang et al., 2012). 또한 엽록소 함량은 SPAD 측정기(CCM-200, opti-sciences Inc, USA)를 이용하여 조사하였다.
5. 통계분석
본 실험은 SPSS statistics program 19.0 (SPSS Inc., Chicago,. IL, USA)을 이용하였으며, 생장 특성은 시비량에 따른 연속변이로 수고, 근원경의 생장량 및 상대생장량의 반응은 반복측정분산분석(Repeated-measures analysis of variance), 광합성 및 엽록소 특성의 분석은 일원분산분석(One-Way ANOVA)을 실시하였다. 각 처리간의 유의성은 DMRT(Duncan's Multiple Range Test) 5% 수준에서 실시하였다(p < 0.05).
결과 및 고찰
1. 간장 및 근원경 생장
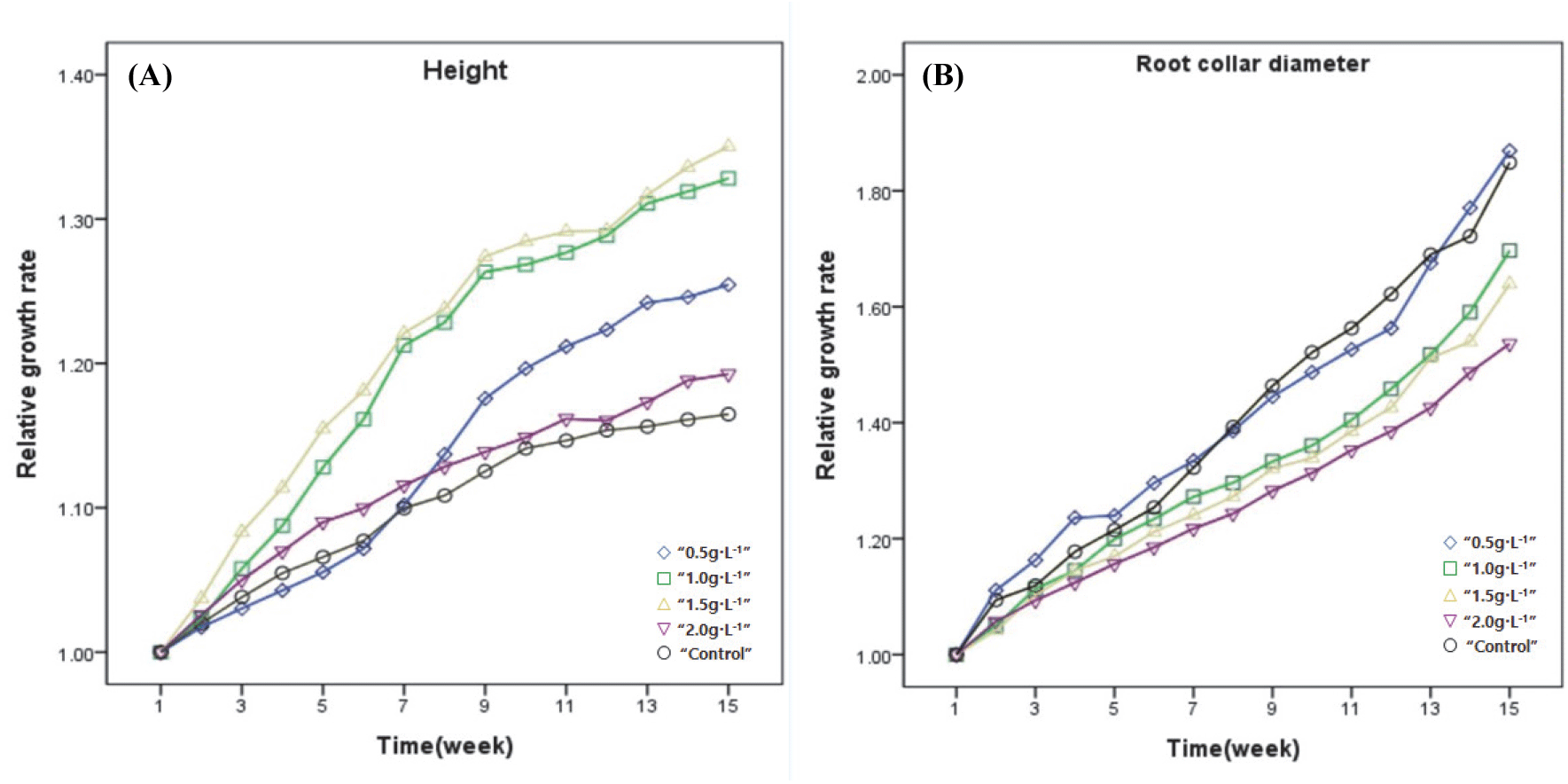
시비처리에 따른 상동나무의 간장 및 근원경의 생장에는 상이한 차이를 보이고 있었다(Table 2). 간장생장량은 1.5 g·L−1에서 가장 높았으며 2.0 g·L−1 < 1.0 g·L−1 및 0.5 g·L−1, 무처리구 순서로 차이를 보였다. 상대생장량은 1.5 g·L−1, 1.0 g·L−1에서 유의적으로 높은 경향을 보였으며 0.5 g·L−1, 무처리구 및 2.0 g·L−1 순으로 차이를 보였다[Figure 1(A)]. 근원경의 경우 생장량은 0.5 g·L−1 처리구에서 가장 높은 경향을 보였으며 1.5 g·L−1, 1.0 g·L−1, 2.0g·L−1, 무처리구 순으로 차이를 보였다. 상대생장량은 0.5 g·L−1, 무처리구가 가장 높은 경향을 보였으며 1.0 g·L−1 및 2.0 g·L−1 < 1.5 g·L−1 순서로 차이를 보였다[Figure 1(B)]. 간장 및 근 원경의 생장량은 대체적으로 1.5 g·L−1, 1.0 g·L−1 처리구에서 가장 우수한 생장을 보였다. 그러나 시비 수준이 가장 높은 2.0 g·L−1에서 낮아지는 것으로 나타났다[Figure 1(B)]. 이는 시비처리가 양묘의 간장 및 근원경의 생장을 향상시키는 것으로 알려져 있으나(Carlson, 1981; Imo and Timmer, 1999; Quoreshi and Timmer, 2000), 적절하지 못한 시비량으로 양분이용효율 감소 등을 일으킬 수 있어(Kwon et al., 2009) 수종에 따라 맞는 적정 시비 수준의 제시가 필요하다(Broschat, 1995; Dumroese et al., 2013; Landis et al., 1989).
Table 2.
The root collar diameter, height, and H/D ratio of S. thea under different fertilizing treatment.
|
Species |
fertilizing treatment |
Height |
Root collar diameter |
H/D (cm·mm) |
|
Growth (cm) |
Relative growth rate (%) |
Growth (mm) |
Relative growth rate (%) |
|
S. thea
|
Control |
0.523±0.113b,c
|
0.017±0.012b
|
0.162±0.024b
|
0.057±0.012a
|
10.90±2.22a,b
|
|
0.5 g·L−1
|
0.469±0.161c
|
0.017±0.008a,b
|
0.251±0.092a
|
0.058±0.028a
|
10.16±1.60b
|
|
1.0 g·L−1
|
0.680±0.329a,b
|
0.022±0.015a
|
0.191±0.066b
|
0.047±0.027a,b
|
12.20±2.94a
|
|
1.5 g·L−1
|
0.746±0.332a
|
0.023±0.016a
|
0.200±0.052b
|
0.043±0.015b
|
11.50±1.94a,b
|
|
2.0 g·L−1
|
0.621±0.290a,b
|
0.013±0.009b
|
0.171±0.041b
|
0.036±0.013b
|
10.94±1.87a,b
|
Download Excel Table
Figure 1.
The growth patterns of root collar height (A) and root collar diameter (B) of fertilizing treatment of S. thea during 15 weeks measurement. Lines show each relative growth rate with repeated measures of ANOVA by Duncan’s multiple range test at 5% levels.
Download Original Figure
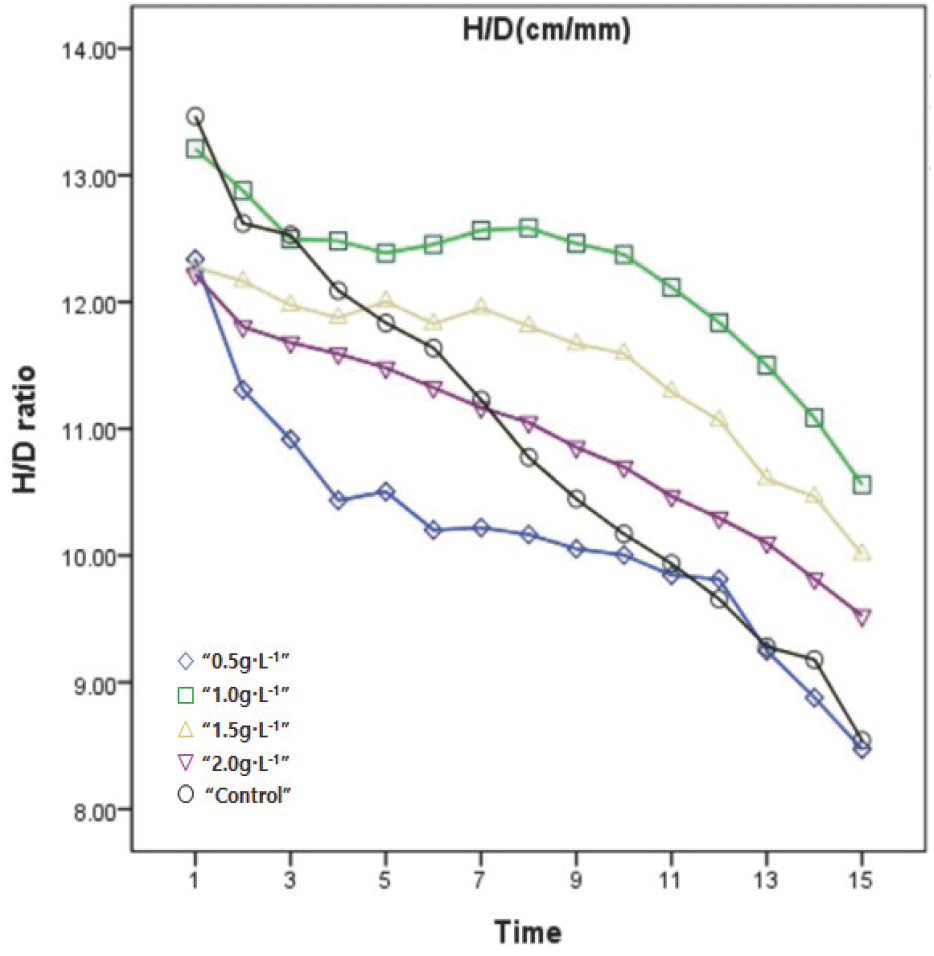
H/D율은 간장과 근원경의 비율로, 생산 묘목이 다부진(stocky)유형에 가까운지, 가늘고 약한(spindly)유형에 가까운지를 수치화한 것으로(Thomson, 1985; Haase 2007), 1.0 g·L−1이 가장 높았으며, 1.5 g·L−1, 2.0 g·L−1, 무처리구, 0.5 g·L−1 순으로 나타났다(Figure 2). 지표상으로 파악하면 무처리구와 0.5g·L−1는 초기 H/D율에 비했을 때 근원경의 생장이 두드러져 다부진 유형에 가까웠으나, 1.0 g·L−1, 1.5 g·L−1, 2.0 g·L−1는 간장 생장이 촉진된 가늘고 약한 유형에 가까웠다. 이러한 노지묘와 용기묘의 생장차이는 기존의 내용과 일치(Johnson et al., 1996) 하는 경향을 보였으며, 국내에서 실시하는 산림용 묘목규격표 활엽수 합격기준 적용 H/D율 값인 9~22 cm·mm(KFS, 2014) 기준을 만족하였다. 이러한 결과는 묘목의 지상부와 지하부 생장 균형이 적절히 이루어진 것으로 시비처리에 따른 이상은 없었다고 판단된다.
Figure 2.
The Height divided by Root collar diameter (H/R) patterns of S. thea during 15 weeks measurement. Repeated measures of ANOVA by Duncan’s multiple range test at 5% levels.
Download Original Figure
2. 광합성 반응
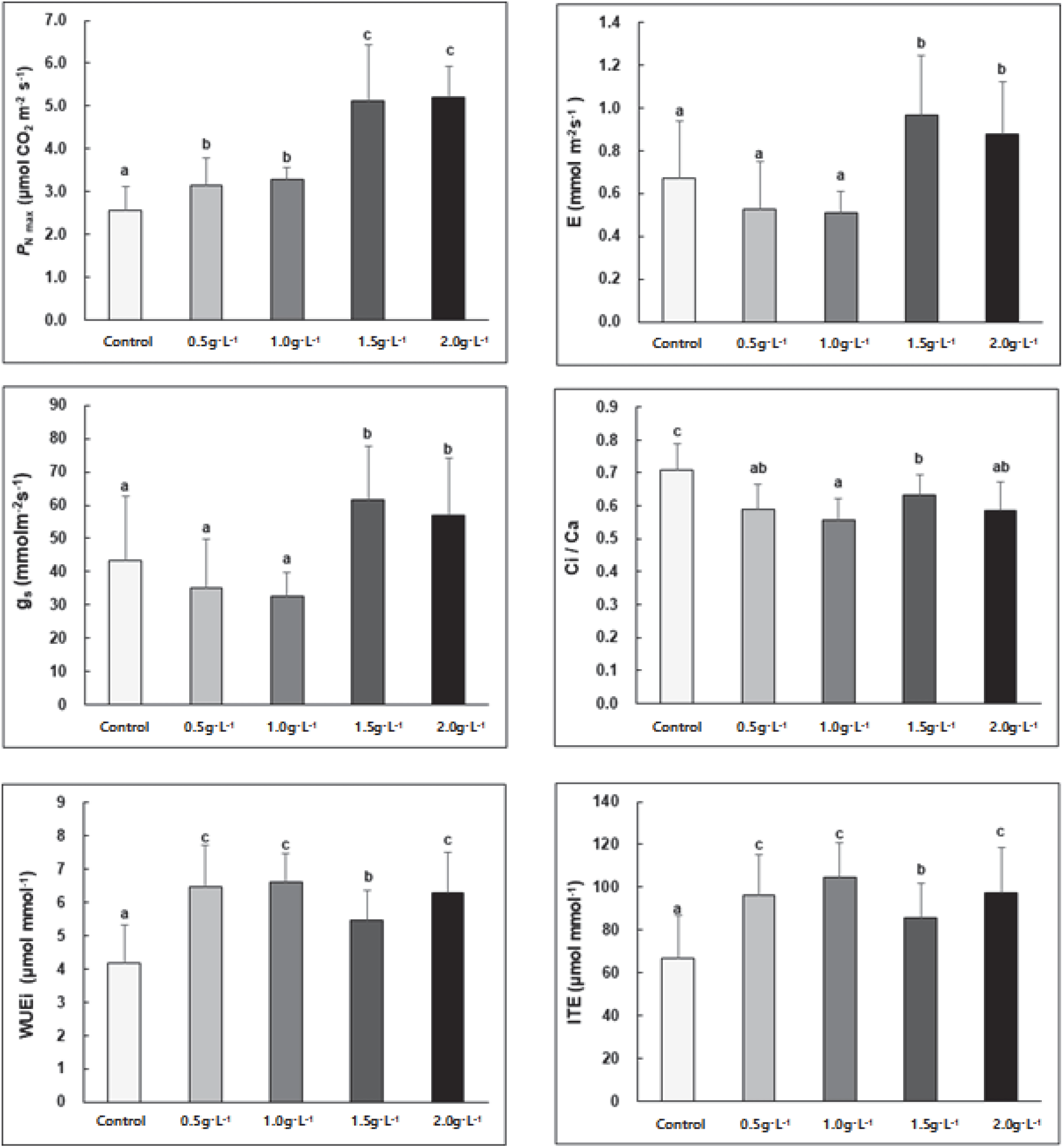
상동나무는 시비처리에 따라 전반적으로 최대광합성속도(maximum photosynthesis rate; PN max), 기공증산속도(stomatal transpiration rate; E), 기공전도도(stomatal conductance; gs), 내재적 수분이용효율(intrinsic water use efficiency, WUEi) 및 순간증산효율(instantaneous transpiration efficiency, ITE)이 증가하는 경향을 보였으며, 엽육세포 내의 CO2 이용효율을 보여주는(Lee et al., 2012) Ci/Ca는 다소 감소하는 것으로 나타났다(Figure 3). 특히 PN max는 무처리구 < 0.5 g·L−1 및 1.0 g·L−1 < 1.5 g·L−1 및 2.0 g·L−1의 순서로 차이를 보였으며, E와 gs는 1.5 g·L−1 및 2.0 g·L−1 처리구에서 유의적으로 높은 경향을 보였다. 내재적 수분이용효율 및 순간증산효율 역시 무처리구에서 상대적으로 가장 낮은 경향을 보였다. 이러한 결과는 시비처리 수준이 높아짐에 따라 기공개폐와 증산속도의 증가로 인해 원활한 수분 및 가스교환이 이루어지고 있음을 보여주며, 엽육세포 내 CO2와 수분을 효율적으로 소비하므로 상대적으로 높은 광합성 능력을 유지하는 것을 알 수 있다.
Figure 3.
Effects of different fertilization treatments on maximum photosynthesis rate (PNmax), stomatal transpiration rate (E), stomatal conductance (gs), intercellular (Ci) / atmospheric CO2 concentration (Ca), intrinsic water use efficiency (WUEi) and instantaneous transpiration efficiency(ITE) in the leaves of S. thea (on PPFD 1,000 μmol·m−2·s). Each value is expressed as the means ± SD (n =5). *Means with difference letters are significantly different by DMRT (Duncan's Multiple Range Test) at 5% level (p < 0.05).
Download Original Figure
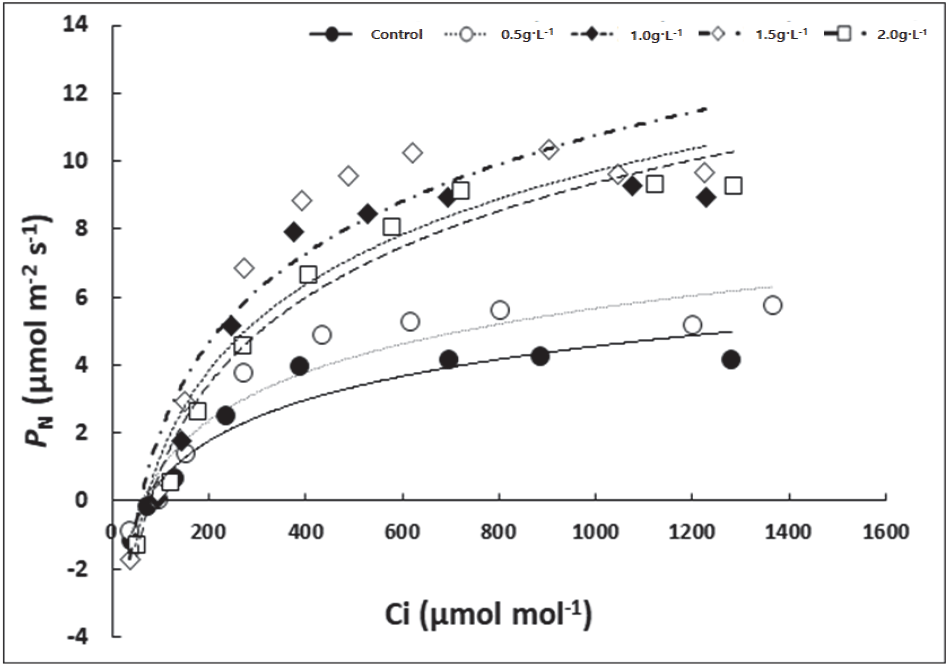
식물의 광합성 능력은 루비스코(Rubisco)의 활성에 따른 카르복실화율 및 리불로오스 이인산 (Ribulose, 5-bisphosphate, RuBP)의 재생 속도간 균형에 의해 결정되며, RuBP의 재생 속도는 전자전달 효율에 의해 제한된다(Hopkins and Hüner, 2008; Ryu et al., 2014). PN-Ci 곡선을 통해 산출한 상동나무의 최대 카르복실화 속도(Maximum carboxylation rate, Vcmax), 최대 전자전달 속도(Maximum electron transport rate, Jmax)는 무처리구와 0.5 g·L−1에서 유의적으로 낮았으며, 1.0 g·L−1 < 1.5 g·L−1 및 2.0 g·L−1 처리구에서 비교적 높은 경향을 나타냈다(Figure 4, Table 3). 특히 무처리구의 경우 가장 높은 경향을 나타낸 1.5 g·L−1에 비해 Vcmax는 약 48% Jmax는 약 45% 수준에 그치는 것으로 나타나 시비처리가 상동나무 광합성 기능 향상에 도움을 주는 것을 알 수 있다. 또한 0.5 g·L−1 시비처리구의 경우 Vcmax와 Jmax는 무처리구에 비해 큰 차이가 없었으나 호흡율(dark respiration rate, RD)을 낮게 유지하여 상대적으로 높은 최대광합성속도(PN max)를 보이는 것으로 나타났다.
Figure 4.
The PN-Ci curves of S. thea grown under five different fertilization treatments. Each value is expressed as the mean (n = 3).
Download Original Figure
Table 3.
The photosynthetic parameters of S. thea grown under five different fertilization treatments.
|
Treatment |
Vcmax (µmol m−2 s) |
Jmax (µmol m−2 s) |
RD (µmol m−2 s) |
|
Control |
16.81±1.74a
|
24.07±4.75a
|
1.02±0.24a
|
|
0.5 g·L−1
|
16.88±1.24a
|
28.08±2.75a
|
0.89±0.01a
|
|
1.0 g·L−1
|
27.17±0.63b
|
45.61±2.53b
|
1.51±0.01b
|
|
1.5 g·L−1
|
34.68±2.17c
|
53.06±9.64b
|
1.62±0.14b
|
|
2.0 g·L−1
|
27.53±1.14b
|
46.63±2.39b
|
1.69±0.08b
|
Download Excel Table
상동나무의 광합성량과 생장량은 무처리, 0.5 g·L−1, 1.0 g·L−1, 1.5 g·L−1에서 정비례하였으나 2.0 g·L−1 생장량이 상대적으로 낮은 수치를 보였다. 이는 용기묘의 수종의 생장 패턴에 맞는 시기별 적정 시비처리가 중요하며, 순수 생장량과 광합성량이 일부 일치하지 않는 경향은 시비 방법이 용기묘의 생장 및 생리적 특성에 미치는 기존의 연구결과와 유사한 경향을 보이고 있다(Jo et al., 2012.)
이는 용기묘의 생육 초기에 세근발달, 간장 생장, 근원경 생장 순으로 발달하는 과정에서 보이는 현상으로 참나무속 수목의 용기 내 생장 및 생리적 반응과 유사한 경향을 보이고 있다(Kim et al., 2014).
3. 엽록소 함량 및 엽록소 형광반응
시비처리에 따른 상동나무의 엽록소 형광반응과 SPAD 값을 Table 4에 나타냈다. 식물은 일반적으로 광합성에 불리한 조건에서 초기형광(Fo)이 높아지고, 최대형광(Fm)은 낮아지는 경향을 보이는데 상동나무는 무처리구에서 최대 형광수율에 대한 초기 형광수율의 비율(Fo/Fm)이 다소 높은 경향을 보이는 것으로 나타났으며. 이와 반대로 광계 2의 최대양자수율을 나타내는 광화학반응 효율(Fv/Fm)과 잠재적 광합성능력을 나타내는 Fv/Fo는 무처리구에서 유의적으로 낮은 경향을 보이고 있다. 에너지 보존효율을 의미하는 PIabs는 광계 II의 최대양자수율(Fv/Fm)보다 더욱 민감한 환경스트레스 지표로 알려져 있으며(Holland et al., 2013; Strasser et al., 2000; Wang et al., 2012), 상동나무의 경우 무처리구가 시비 처리구에 비해 약 3.4배 낮은 경향을 보이는 것으로 나타났다. 또한 엽록소함량을 간접적으로 나타내는 SPAD 역시 무처리구에서 유의적으로 낮은 것으로 나타났다. 이를 통해 시비처리가 광합성 기구의 효율을 높이고 있는 것을 알 수 있다.
Table 4.
The chlorophyll fluorescence parameters of S. thea grown under five different fertilization treatments.
|
Treatment |
Fo/Fm |
Fv/Fm |
Fv/Fo |
PIabs
|
SPAD |
|
Control |
0.20±0.01b
|
0.80±0.01a
|
3.94±0.30a
|
1.47±0.42a
|
34.6±2.6a
|
|
0.5 g·L−1
|
0.18±0.01a
|
0.82±0.01b
|
4.43±0.16b
|
4.98±0.64b
|
42.7±3.8bc
|
|
1.0 g·L−1
|
0.19±0.02a
|
0.81±0.02b
|
4.36±0.53b
|
5.04±1.28b
|
39.8±6.5abc
|
|
1.5 g·L−1
|
0.18±0.01a
|
0.82±0.01b
|
4.54±0.13b
|
4.53±1.50b
|
38.9±3.3ab
|
|
2.0 g·L−1
|
0.18±0.01a
|
0.82±0.01b
|
4.54±0.19b
|
5.03±1.28b
|
45.3±3.7c
|
Download Excel Table
결 론
본 연구는 상동나무 용기묘를 대상으로 시비처리에 따라 나타나는 생장특성인 간장, 근원경, H/D율과 기초적인 대사특성인 광합성, 엽록소 함량 및 형광반응을 분석하여 시설양묘에 필요한 최적의 시비 수준을 제안하고자 실시하였다.
시비처리에 따른 상동나무의 간장생장은 시비 수준에 따라 유의미하게 상승했으나 2.0 g·L−1 처리구의 경우 상대생장률이 크게 낮아졌다. 근원경의 생장은 시비 수준에 따라 상대생장률이 유의미하게 떨어졌으나 전체적으로 1.5 g·L−1, 1.0 g·L−1 처리구가 우수한 형질을 나타냈다. 또한 상동나무에 대한 시비처리가 간장생장을 촉진하면서 근원경의 생장은 상대적으로 낮아지는 경향을 보이고 있었다. 하지만 전체적으로 시비처리구가 무처리구에 비해 전체적인 생장량이 높아졌으며 효과가 유의미했다고 볼 수 있었다. 광합성 능력을 확인할 수 있는 최대광합성속도, 기공증산속도 등이 0.5 g·L−1, 무처리구와 비교해서 1.5 g·L−1, 2.0 g·L−1 처리구가 상대적으로 높은 수치를 나타내고 있었으며, 엽록소 함량을 간접적으로 나타내는 SPAD 또한 시비처리구가 무처리구에 비해 3.4배 높은 경향을 띄고 있었다. 이를 통해 상동나무에 대한 시비처리는 광합성과 광합성기구의 효율을 높이고 있는 것을 확인하였다.
따라서 수준별 시비처리는 상동나무 용기묘의 생장량을 높임과 동시에 묘목 건전성을 높이는 것으로 확인되었으며, 적정한 시비처리 수준은 1.0 g·L−1 ~ 1.5 g·L−1 로 판단된다.
감사의 글
본 연구는 국립산림과학원 석⋅박사연구원 지원 사업과 일반연구과제를 통해 이루어진 것으로 이에 감사드립니다.
References
Bayala, J., Dianda, M., Wilson, J., Ouedraogo, S.J. and Sanon, K. 2009. Predictiong field performance of five irrigated tree species using seedling quality assessment in Burkina Faso, West Africa, New Forests 38(3): 309-322.

Broschat, T.K. 1995. Nitrate, phosphate, and potassium leachingfrom container-grown plants fertilized by several methods. Hortscience 30: 74-77.
Bumgarner, M.L., Salifu, K.F. and Jacobs, D.F. 2008. subirrigation of
Quercus rubra seedlings: Nursery stock quality, media chemistry, and early field performance. Hortscience 43(7): 2179-2185.

Burdett, A.N. 1990. Physiological processes in plantation establishment and the development of specifications for forest planting stock. Canadian Journal of Forest Research 20(4): 415-427.
Carlson, W.C. 1981. Effects of controlled-release fertilizers on the shot and root development of outplanted western hemlock (
Tsuga heterophylla Raf. Sarg.) seedlings. Canadian. Journal of Forest Research 11(1981): 752-757.
Cho, E.J. 2000. A survey on the usage of wild grasses. Korean J. Dietary Culture 15(1): 59-68.
Choo, G.C. 1992. Systematic studies of the family Rhamnaceae in Korea Ph.D. Thesis. Konkuk University. pp. 22-70.
Chung, J.M., Cho, S.H., Kim, Y.S., Kong, K.S., Kim, H.J., Lee, C.H. and Lee, H.J. 2017, Ethnobotany in Korea: The traditional knowledge and use of indigenous plants. Korea National Arboretum. Pocheon, Korea. pp. 1048.
Dumroese, R.K., Sung, S.S., Pinto, J.R., Davis, AS. and Scott, D.A. 2013. Morphology, gas exchange, and chlorophyll content of long leaf pine seedlings in response to rooting volume, copper root pruning, and nitrogen supply in a container nursery. New Forests 44(2013): 881-897.
Grossnickle, S.C. 2005. Importance of root growth in overcoming planting stress. New Forests 30(2005): 273-294.
Haase, D.L., Riley, L.E., Dumroese, R.K. and Landes, T.D. 2007 Morphological and physiological evaluation of seedling quality. Proceedings of the Conference “Forest and Conservation Nursery Associations. 2006. pp. 3-8.
Holland, V., Koller, S. and Brüggemann, W. 2013. Insight into the photosynthetic apparatus in evergreen and deciduous European oaks during autumn senescence using OJIP fluorescence transient analysis. Plant Biology 16(4): 801-804.
Hopkins, W.G. and Hüner, N.P.A. 2008. Introduction to plant physiology. (4nd ed.). John Wiley and Sons. New York, NY, USA. pp. 223-230.
Imo, M. and Timmer, V.R. 1999. Vector competition analysis of black spruce seedling responses to nutrient loading and vegetation control. Canadian Journal of Forest Research 29(4): 474-486.
Johnson, F., Paterson, J., Leeder, G., Mansfield, C., Pinto, F. and Watson, S. 1996. Artificial regeneration of Ontario’s forest: Species and stock selection manual. Forest Research Information Paper 1996(131): pp. 51.
KFS (Korea Forest Service). 2014. The guidelines for seed and nursery practices. pp. 100.
Jo, M.S., Lee, S.W. and Park, B.B. 2012. Effects of three fertilization methods on the growth performances and physiological characteristics of container seedling. Korean Forest Society 2012,-(2012): 570-573.
Kim, D.K., Oh, K.H., Lee, B.Y., Kim, M.H., Kim, T.K., Lee, E.Y., Roh, H.C., Lee, M.H. and Rhee, D.G. 2004. Management of korean biological resources for access regulation and benefit-sharing. Korean Journal of Environmental Biology 22(2): 259-264.
Kim, H.N., Park, G.H., Kim, J.D., Park, S.B., Eo, H.J. and Jeong, J.B. 2019a. Effect of the extracts from the leaves and branches of
Sageretia thea on β-catenin proteasomal degradation in human colorectal and lung cancer cells. Korean Journal of Plant Resources 32(2): 153-159.
Kim, H.N., Park, G.H., Park, S.B., Kim, J.D., Eo, H.J., Son, H.J., Song, J.H. and Jeong, J.B. 2019b. Extracts from
Sageretia thea reduce cell viability through inducing cyclin D1 proteasomal degradation and HO-1 expression in human colorectal cancer cells. BMC Complementary and Alternative medicine 19(1): 43.
Kim, H.N., Park, G.H., Park, S.B., Kim, J.D., Eo, H.J., Son, H.J., Song, J.H. and Jeong, J.B. 2019c.
Sageretia thea inhibits inflammation through suppression of NF-κB and MAPK and activation of Nrf2/HO-1 signaling pathways in RAW264.7 cells. The american Journal of Chinese Medicine 47(2): 385-403.
Kim, J. S. and Hyun, T. K. 2015. Nutrients and antioxidative activities of
Sageretia theezans fruit. The Korean Society of Medicinal Crop Science 2015(5): 225-226.
Kim, J. S. and Kim, T.Y. 2011. Woody plants of Korean peninsula. Dolbegae. Seoul, Korea. pp. 688.
Kim, J.D., Park, S.B., Eo, H.J. Park, G.H. and Jeong. J.B. 2020. Induction of apoptosis by
Sageretia thea branch extracts through activation of NF-κB signaling pathway in human colorectal cancer cells. Korean Journal of Plant Resources 33(5): 428-435.
Kwon, K.W., Cho, M.S., Kim, G.N., Lee, S.W. and Jang, K.H. 2009. Photosynthetic characteristics and growth performances of containerized seedling and bare root seedlingof Quercus acutissima growing at different fertilizing schemes. Journal of Korean Forest Society 98(3): 331-338.
Landis, T.D., Tinus, R.W., McDonald, S.E. and Barnett, J.P. 1989. Seedling nutrition and irrigation. The container tree nursery manual Vol. 4. USDA Forest Service. AgricultureHandbook. Washington. pp. 674.
Lee, K.C., Noh, H.S., Kim, J.W. and Han, S.S. 2012. Physiological responses of
Cirsium setidens and
Pleurospermum camtschaticum under different shading treatments. Journal of Bio-environment Control 21: 15361.
Oh, S.J. and Koh, S.C. 2017. Photosynthesis and growth responses of soybean (
Glycine max Merr.) under Elevated CO
2 Conditions. Journal of Environmental Science International 26(5): 601-608.
Pyo, S.J., Lee, Y.J., Park, S.I., Lee, C.I., Park, J.Y. and Sohn, H.Y. 2020. Evaluation of the Anti-thrombosis Activities of the Aerial Parts of
Sageretia thea. Jounal of Life Science 30(5): 443-451.
Quoreshi, M. and Timmer, V.R. 2000. Early outplanting performance of nutrient-loaded containerized black spruce seedlings inoculated with Laccaria Biocolor: A bioassay study. Soil Science Society of America Journal 30(5): 744-752.
Ryu, D.U., Bae, J.H., Park, J.H., Cho, S.S., Moon, M.K., Oh, C.Y. and Kim, H.S. 2014. Responses of native trees species in Korea under elevated carbon dioxide condition-open top chamber experiment. Korean Journal of Agricultural and Forest Meteorology 16(3): 199-212.
Sharkey, T.D., Bemacchi, C.J., Farquhar, G.D. and Singsaas, E.L. 2007. Fitting photosynthetic carbon dioxide response curves for C3 leaves. Plant, Cell and Environment 30: 1035040.
Song, J.H., Yang, S.G., Choi, G.Y. and Moon, B.C. 2020. Analysis on the trends of Korean health functional food patent based on the medicinal plant resources. Korean Herbal Medicine Informatics 8(1): 25-44.
Song, S.C., Song, C.K. and Kim, J.S. 2014. Vegetation and habitat environment of Sageretia thea in Jeju Island. The Korean Society of Medicinal Crop Science 22(4): 301-305.
Song, S.C., Song, C.K. and Kim, J.S. 2015a. Characteristics of seed-germination and fruit for
Sageretia thea in Jeju region. The Korean Society of Medicinal Crop Science 23(1): 8-12.
Song, S.C., Song, C.K. and Kim, J.S. 2015b. Effect of plant growth regulators on fruit enlargement and optimal harvest time in
Sageretia thea (Osback) M. C. Johnst. The Korean Society of Medicinal Crop Science 23(1): 311-318.
Strasser, R.J., Srivastava, A. and Tsimilli-Michael, M. 2000. The fluorescence transient as a tool to characterize and screen photosynthetic samples.
In Yunus M, Pathre U and Mohanty P. (eds.). Probing Photosynthesis: Mechanism, regulation and adaptation. Taylor and Francis. London and New York. pp. 445-483.
Thompson, B.E. 1985. Seedlings morphological evaluation. What you can tell by looking: In Duryea, M.L. (ed.), Evaluating seedling quality: Principles, procedures, and predictive abilities of major tests. For. Res. Lab., Oregon State Univ., Corvallis., OR, USA. pp. 59-72.
Wang, Z.X., Chen, L., Ai, J., Qin, H.Y., Liu, Y.X., Xu, P.L., Jiao, Z.Q., Zhao, Y. and Zhang, Q.T. 2012. Photosynthesis and activity of photosystem II in response to drought stress in amur grape (
Vitis amurensis Rupr.). Photosynthetica 50(2012): 18996.