서 론
대기오염은 현재 세계적으로 가장 중요한 환경 문제 중 하나로 농작물과 생태계에 심각한 악영향을 미치고, 호흡기 및 심혈관 질환을 유발하여 사람들의 건강에 위협이 되고 있다(Lee, 2019; Pandey et al., 2005; WHO, 2006). 대표적인 대기오염물질인 이산화질소(NO2)와 이산화황(SO2)은 석탄, 석유의 연소 과정에서 발생하며 주로 자동차 배기가스나 공장 매연에서 발생한다. NO2는 그 자체로 폐 질환을 유발하기도 하지만, 자외선에 의한 광화학 반응으로 오존(O3)과 질산과산화아세틸(PAN)의 발생에 중추적인 역할을 하며, 발생한 O3와 PAN은 사람의 눈, 호흡기 점막을 손상시킨다. SO2는 폐렴, 기관지염, 천식과 알레르기 및 염증 질환을 일으키며 산성비의 주된 원인이 된다. NO2와 SO2가 대기 중의 수증기와 결합하면 강산성의 황산(H2SO4)이나 질산(HNO3)을 형성하여 건축물을 부식시키고 토양과 수질을 산성화시켜 생태계에 피해를 준다(Cho et al., 2020; Larssen et al., 2006; Meng et al., 2008; Sudalma et al., 2015; Xie et al., 2009). 또한 NO2와 SO2는 휘발성유기화합물(VOCs)과 함께 2차 에어로졸의 전구체로 작용하여 인간의 건강 및 산업기반에 피해를 주는 미세먼지를 형성하기도 하며, 이에 따라 NO2와 SO2를 저감하기 위한 기술이 개발되거나 제도가 시행되는 등 대기오염물질 저감 방안이 강구되고 있다(Liu et al., 2010; Xie et al., 2015).
대기오염물질을 저감하는 방안 중 하나로 떠오르는 것이 산림과 식생을 이용한 방법으로, 이는 경제적이고, 친환경적이며, 또한 공학적 방법에 의한 부작용을 개선할 수 있다(Chaparro-Suarez et al., 2011; Jo and Ahn, 2001; Kim and Koh, 1996; Manninen and Huttunen, 2000). 산림과 식생을 통한 NO2와 SO2의 저감은 식물의 광합성 과정에서 잎의 기공을 통해 이산화탄소와 함께 흡수되어 이루어진다(Singh and Tripathi, 2007; Wei et al., 2017). 따라서 식생의 유형과 산림의 구조는 대기오염물질 저감 기작에 영향을 미치는데, Jo et al.(2002)에 의하면 동일 영급 내에선 침엽수림 보다 혼효림이나 활엽수림의 NO2 및 SO2 저감이 더 컸고, 식생의 단위면적당 저감량은 기저면적의 변화와 연관되어 영급이 높을수록 증가하는 것으로 나타났다. 산림 구조 측면에서는 산림 면적, 엽면적지수(LAI), 흉고 단면적 증가량(BAI) 등이 대기오염물질 저감과 직접적인 관련이 있는 것으로 나타났다(Muzika et al., 2004; Nowak et al., 2008). 특히 도시 지역은 많은 양의 공장 매연과 차량 배기가스, 공기의 흐름이 적은 구조로 인해 다른 지역에 비해 대기오염물질 농도가 높게 나타나며, 사람들의 일상에 연관되어 그 심각성이 떠오르고 있다. 도시 지역과 도로 인근에는 조경수 및 가로수를 심어 식생을 형성함으로써 대기오염물질을 저감하고 대기질을 개선할 수 있는데, 실제로 도시림 내부에서는 수목의 효과로 인해 도시림 외부보다 NO2와 SO2 농도가 더 낮은 것으로 나타났다(Manning, 2008; Yang et al., 2018). 앞서 언급한 식물의 NO2와 SO2 저감은 잎의 광합성 속도, 기공전도도, 엽록소 함량 등 잎의 생리적 특성과 생물량, 두께, 기공의 수 등 잎의 형태적 특성의 영향을 받는 것으로 알려져 있다(Dhir, 2016; Hu et al., 2011; Mukherjee and Agrawal, 2016; Woo et al., 2004). 무엇보다 수목의 NO2와 SO2 저감 능력은 수종에 따라 큰 차이가 나타나는데, 이는 앞서 말한 수목의 특성들이 수종에 따라 다르기 때문이다. 많은 연구에서 높은 단위 엽면적당 광합성능과 기공전도도로 인해 침엽수종에 비해 활엽수종이, 상록수종에 비해 낙엽수종이 NO2와 SO2 저감 능력이 높다고 설명한다(Jo and Ahn, 2001; Okano et al., 1989; Takahashi et al., 2005).
수목을 이용한 대기오염물질 저감의 중요성과 관심이 증가하고 있음에도 불구하고, 대부분의 선행연구들은 대기오염물질의 피해에 따른 수목의 생화학적, 생리적 반응에 관해서만 설명하고 있다(Singh and Tripathi, 2007; Wei et al., 2017). 하지만, 대기오염물질 저감 능력이 우수한 수종을 선발하고 저감 능력과 관련된 인자를 판명하기 위해서는 수종별 대기오염물질 저감 능력 정량화와 수종별 특성과의 관련성 연구가 필요하며, 특히 도로 및 도시 근처에서 발생하는 대기오염물질의 저감에 가장 중요한 역할을 하는 가로수 및 조경수에 대한 연구가 필요하다. 따라서 본 연구에서는 챔버를 이용한 실내 실험을 통해 (1) 주요 가로수 10 수종의 NO2와 SO2 저감 능력 정량화를 통한 대기오염물질 저감 능력 우수 수종을 선정하였고, (2) 수목 잎의 생리적, 형태적 특성과 NO2 및 SO2 저감량의 상관관계 분석을 통해 대기오염물질 저감 능력의 관련 인자를 판명하였다.
재료 및 방법
연구 수종은 조경수 및 가로수로 심는 주요 수종으로 느티나무(Zelkova serrata (Thunb.) Makino), 배롱나무(Lagerstroemia indica L.), 병꽃나무(Weigela florida (Bunge) A. DC.), 산수유(Cornus officinalis Siebold & Zucc.), 왕벚나무(Prunus × yedoensis Matsum.), 화살나무(Euonymus alatus (Thunb.) Siebold)의 피자식물 6 수종과 곰솔(Pinus thunbergii Parl.), 소나무(Pinus densiflora Siebold & Zucc.), 스트로브잣나무(Pinus strobus L.), 은행나무(Ginkgo biloba L.)의 나자식물 4 수종을 대상으로 하였으며, 수종별 3본을 사용하였다. 생장이 비슷한 실생 2년의 묘목(묘고 50-100 cm)을 묘목시장에서 구입하여 이용하였으며 배양토를 채운 5L 플라스틱 포트에 이식하여 실험을 진행하였다. 정상적인 생육을 위하여 식물은 실외에 위치해 있다가 가스 실험 직전에 실험실로 들여왔으며, 수분 스트레스를 방지하기 위하여 이틀에 한 번씩 충분한 양의 물을 관수하였다. 묘목들은 병충해나 물리적 피해가 없는 건강한 상태에서 실험에 사용되었으며, 실험 동안에는 가스가 토양으로 흡수되는 것을 방지하기 위하여 포트를 비닐로 감싸고 진행하였다.
시간에 따른 NO2와 SO2 저감 실험은 서울대학교(37°27’33” N, 126°57’7” E) 내 식물환경조절실에서 2020년 3월부터 9월까지 진행하였다. 0.28 m3[(0.3)2 × π × 1 m3] 크기의 아크릴 챔버를 사용하여 NO2와 SO2 농도를 한 종류씩 주입하여 측정하였으며, 흡입구를 챔버 상단부에 설치하여 가스가 챔버 내에서 균일하게 섞이도록 하였다. 30 L 가스 실린더에 부착한 레귤레이터와 챔버 흡입구에 설치한 유량계를 조절하여 500 cc min−1의 일정한 속도로 가스를 주입하였다(Figure 1). 챔버의 초기 가스 농도는 환경정책기본법 시행령 환경기준 제2조(국가법령정보센터, 2020. 5.27 개정)의 1시간 평균 기준치를 적용하여 NO2 농도는 0.1 ppm, SO2 농도는 0.15 ppm으로 설정하였다. NO2 농도 변화는 서울대학교 농업생명과학공동기기원(NICEM)의 질소산화물 분석기(NOx analyzer, Teledyne instruments, USA)를 이용하였으며, SO2 농도는 NICEM의 HCHO, VOCs 휴대용 분석기(AirWell Plus, ㈜한국산업기기, Korea)를 이용하여 1분 평균값을 2시간 이상 수종별 3반복 측정하였다.
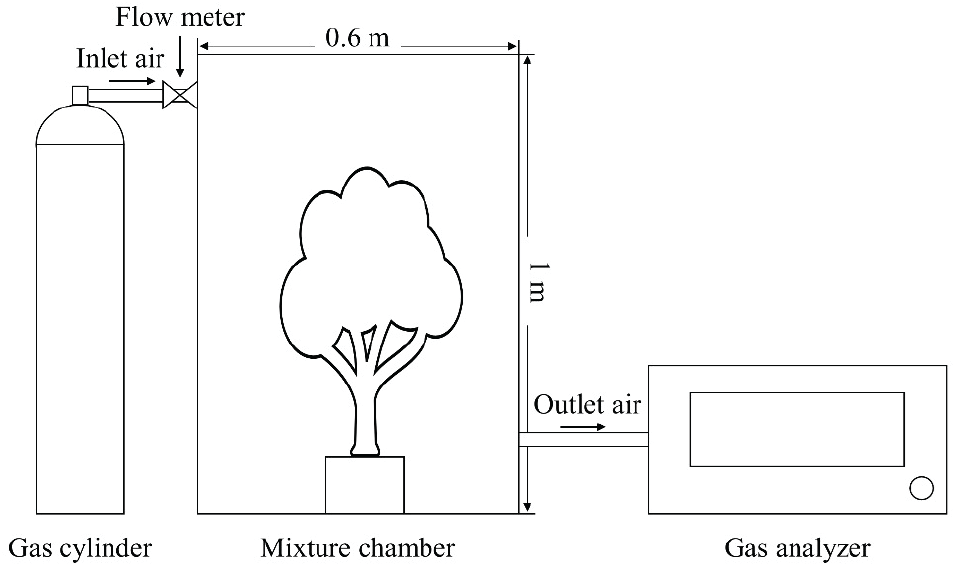
본 실험에서 수종별 NO2 저감량은 초기 농도(CONCi)와 실험 120분(CONC120) 시점의 농도 차이로 계산하였는데, 이는 실험 시작 후 120분경에 빈 챔버와 묘목을 넣은 챔버의 농도 차이가 가장 크게 나타나, 실질적인 수목의 가스 저감이 가장 큰 시간으로, 수종별 차이를 나타내기 가장 적합하다고 판단되었기 때문이다. 마찬가지로 수종별 SO2 저감량은 농도 차이가 가장 크게 나타나는 실험 시작 70분 후의 농도(CONC70)와 초기 농도의 차이로 계산하였다. 묘목을 넣지 않은 대조구 실험을 진행하여 자연적으로 감소하는 가스 농도(RCONCe)를 측정하여 제외함으로써 순전히 식물이 저감하는 가스량(RNO2, RSO2)을 계산하였다. 또한 가스 농도 측정기기에서 나타내는 ppm은 수종별 비교를 위해 25 ºC, 1기압에서의 기체 1몰의 부피(24.4513L)와 NO2, SO2 각각의 분자량, 챔버 부피(Vch), 묘목의 총 엽면적(TLA, Table 1)을 계산하여 엽면적당 가스 저감량(RNO2leaf, RSO2leaf)으로 나타내었으며, 그 식은 아래와 같다(식 1, 2). 묘목의 총 엽면적은 전체 잎의 건중량에 엽면적당 건중량(LMA)을 곱하여 계산하였으며, 단위 통일을 위해 모든 저감량은 시간당 저감량(ng cm−2 hr−1)으로 나타내었다. 측정 동안에는 LED 램프(240 W 식물생장용 LED, 빛솔 LED, Korea)를 이용해 챔버 내부의 수목이 광합성을 할 수 있는 충분한 광량을 제공하였고, 각 실험 후 챔버는 30분 이상 환기 후, 증류수로 세척하고 실험용 와이퍼로 닦았다.
Data are represented as mean ± standard error (SE).
잎 내 엽록소 함량은 Hiscox and Israelstam (1979)에 의해 개발된 다이메틸설폭사이드(DMSO) 추출법을 이용하여 분석하였다. 가스 실험 후 같은 가지에서 여섯 개의 잎을 샘플링한 후, 활엽수는 주맥이 포함되지 않도록 직경 1 cm의 디스크를 뚫었고, 침엽수는 1 mm 이내로 잘랐다. DMSO 5 ml 가 담긴 갈색 바이알에 활엽수의 경우 디스크 두 개, 침엽수의 경우 0.02 g의 생엽을 넣고 항온수조에서 65°C로 6시간 이상 중탕시켜 엽록소를 추출한 후 분광광도계(Optizen 2120 UV, Mecasys, Korea)를 이용하여 흡광도를 측정하였다.엽록소와 카로티노이드 함량은 Wellburn (1994)의 DMSO 식을 사용하였으며, 이때 A649, A665, A480는 649 nm, 665 nm, 480 nm 파장에서 측정된 흡광도이고, Ca 와 Cb는 엽록소 a와 b 함량이다(식 3~6).
활엽수의 경우 5개, 침엽수의 경우 20개의 신선한 잎을 샘플링하여, 엽면적, 건중량, 엽면적당 건중량(LMA, g m−2), 엽면적비(SLA, m2 kg−1), 잎의 둘레를 측정하였다. 하층부 엽량이 적은 것을 고려하여 수관 상층부, 중층부, 하층부에서 2:2:1의 비율로 샘플링하였다. 엽면적은 스캐너(Epson perfecture V370 Photo, Seiko, Japan)와 ImageJ(ver. 1.5.2, National Institutes of Health, USA)를 사용하여 측정하였다. 가스 저감량 계산시 활엽수와 은행나무의 경우 기공이 잎의 뒷면에 존재하는 이면기공엽이지만, 침엽수의 경우 모든 면에 존재하는 양면기공엽이기 때문에 이를 고려하여 활엽수와 은행나무의 경우 투영 면적을 적용하였고, 침엽수인 소나무와 곰솔의 경우 2.571, 스트로브잣나무의 경우 2.77의 비율을 곱하여 표면적을 계산하였다(Ellsworth et al., 2012). 잎의 건중량은 신선한 잎을 건조기에서 70°C 로 72시간 이상 건조한 후 측정하였다. 잎의 둘레는 파이썬(ver. 3.8, Python software foundation, USA) OpenCV 패키지의 가우시안 흐림 필터(Gaussian blur filter)와 문턱 값(threshold)을 스캔 사진에 적용하여 측정하였다.
수종별 대기오염물질 저감량의 유의한 차이를 확인하기 위해 일원분산분석(one-way ANOVA)을 수행하였다(P < 0.05). 통계적으로 유의한 차이가 확인된 경우, Duncan의 사후 검정을 통해 수종별 차이의 유의성을 검정하였다. 상관관계분석은 대기오염물질 저감량과 잎의 생리적(엽록소, 카로티노이드 함량), 형태적(SLA, 엽면적당 둘레비) 특성 사이의 선형 관계를 설명하기 위해 수행하였다(P < 0.05). 측정된 특성의 가장 적합한 조합을 찾기 위해 예측 변수를 순차적으로 추가 및 제거하는 단계적 다중 회귀분석(stepwise multiple regression)을 수행하였으며, 모델 정확도를 개선하고 과적합을 방지하기 위해 10겹 교차 검증(10-fold cross validation)을 수행하였다. 모든 통계분석은 R 프로그램(ver. 4.0.2, R Core Team, New Zealand)을 사용하여 수행하였다.
결과 및 고찰
Figure 2a는 수종별 120분 동안의 NO2 농도 변화이다. 대조구 실험인 비어 있는 챔버의 경우 NO2 농도는 실험 시작 시 0.105 ± 0.001 ppm에서 실험 종료 시 0.085 ± 0.002 ppm으로 0.02 ± 0.001 ppm 감소하였는데, 이는 농도 측정을 위해 분당 500 cc의 속도로 챔버내 공기를 측정기기로 배출하였기 때문이다. 수목을 넣은 챔버 실험에서 NO2 농도는 수종에 관계없이 120분 동안 지속적으로 감소하였으며, 따라서 본 연구에서는 실험 시작 후 120분까지의 NO2 저감량을 적용하여 수종별 저감 능력을 비교하였다. 또한 개체목별 총 엽면적의 차이를 고려하여 저감량을 엽면적으로 나누어 저감능력을 정규화하였다.
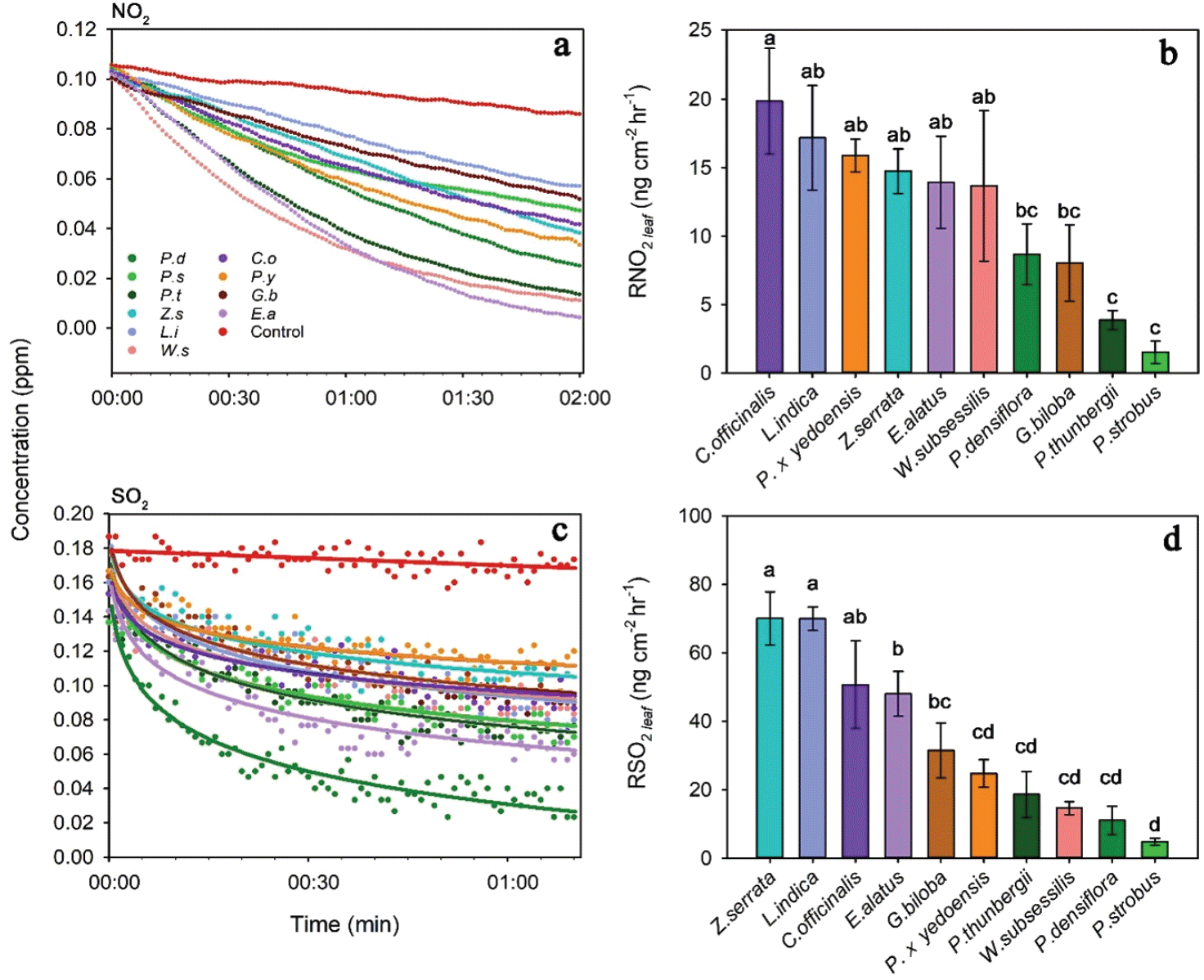
수목의 단위 엽면적당 NO2 저감 능력은 수종에 따라 유의하게 다르게 나타났다(P = 0.004) (Figure 2b). 엽면적당 NO2 저감량은 산수유(19.81 ± 3.84 ng cm−2 hr−1), 배롱나무(17.14 ± 3.79 ng cm−2 hr−1), 왕벚나무(15.85 ± 1.18 ng cm−2 hr−1), 느티나무(14.7 ± 1.61 ng cm−2 hr−1), 화살나무(13.9 ± 3.36 ng cm−2 hr−1), 병꽃나무(13.64 ± 5.47 ng cm−2 hr−1), 소나무(8.66 ± 2.2 ng cm−2 hr−1), 은행나무(8.01 ± 2.78 ng cm−2 hr−1), 곰솔(3.87 ± 0.69 ng cm−2 hr−1), 스트로브잣나무(1.51 ± 0.81 ng cm−2 hr−1) 순으로 나타났으며, 저감량이 가장 많은 산수유의 경우 가장 적은 스트로브잣나무의 약 13.1배였다. 기공 분포를 고려하여 분류한 결과 이면기공엽을 가진 활엽수 6수종과 은행나무의 평균 엽면적당 NO2 저감량은 14.72 ± 1.32 ng cm−2 hr−1으로 양면기공엽을 가진 침엽수 3수종의 평균 저감량인 4.68 ± 1.26 ng cm−2 hr−1의 약 3.1배로 나타나, 활엽수와 은행나무가 침엽수보다 단위면적당 NO2 저감능력이 높은 것으로 확인되었다(P < 0.001).
Figure 2c는 수종별 70분 동안의 SO2 농도 변화이다. 대조구 실험인 비어 있는 챔버의 경우 SO2 농도는 실험 시작 시 0.186 ± 0.012 ppm에서 실험 종료 시 0.173 ± 0.008 ppm으로 0.013 ± 0.003 ppm 감소하였으며, 이는 NO2의 경우와 마찬가지로 농도 측정을 위해 분당 500 cc의 속도로 챔버내 공기를 측정기기로 배출하였기 때문이다. 수목을 넣은 챔버 실험에서 SO2 농도는 초기에 급격히 감소하고 시간이 지날수록 저감 속도가 감소하여 70분 후에는 일정해지는 경향이 나타났다. 따라서 본 연구에서는 실험 시작 후 70분 까지의 SO2 저감량을 적용하여 수종별 저감 능력을 비교하였으며 저감량을 개체목별 엽면적으로 나누어 NO2의 경우와 같은 방법으로 저감능력을 정규화하였다.
수목의 단위 엽면적당 SO2 저감 능력은 수종에 따라 유의하게 다르게 나타났다(P < 0.001) (Figure 2d). 엽면적당 SO2 저감량은 느티나무(70.04 ± 7.74 ng cm−2 hr−1), 배롱나무(69.94 ± 3.44 ng cm−2 hr−1), 산수유(50.67 ± 12.76 ng cm−2 hr−1), 화살나무(48.01 ± 6.48 ng cm−2 hr−1), 은행나무(31.46 ± 8.05 ng cm−2 hr−1), 왕벚나무(24.71 ± 3.99 ng cm−2 hr−1), 곰솔(18.6 ± 6.67 ng cm−2 hr−1), 병꽃나무(14.61 ± 1.89 ng cm−2 hr−1), 소나무(11.01 ± 4.1 ng cm−2 hr−1), 스트로브잣나무(4.79 ± 1.02 ng cm−2 hr−1) 순으로 나타났으며, 엽면적당 SO2 저감량이 가장 많은 느티나무의 경우 가장 적은 스트로브잣나무의 약 14.6배였다. 기공 분포를 고려한 분류 결과 활엽수 6수종과 은행나무의 평균 엽면적당 SO2 저감량은 44.21 ± 5.01 ng cm−2 hr−1으로 침엽수 3수종의 평균 저감량인 11.47 ± 3.03 ng cm−2 hr−1의 약 3.9배로 나타나, NO2의 경우와 마찬가지로 활엽수와 은행나무가 침엽수보다 엽면적당 SO2 저감능력이 높은 것으로 확인되었다(P < 0.001).
Figure 2a에서 NO2 농도는 상대적으로 일정한 속도로 감소하는 반면, Figure 2c의 SO2 농도는 초반에 급격히 감소한 후 점차 저감 속도가 느려지는 것으로 나타났는데, 이는 기공 폐쇄에 따른 결과로 생각된다. NO2와 SO2에 결합하고 있는 산소 분자는 기공을 통해 흡수된 후 산소 원자로 분리되며, 광합성 과정에서 발생한 전자와 결합하여 활성산소종(reactive oxygen species)을 생성한다. 활성산소는 수목의 체내에서 산화작용, 세포막 기능 저하, 효소 활성 저해 등의 피해를 입히며(Woo, 2021), 기공의 폐쇄에도 영향을 주는 것으로 알려져 있다(Yan et al., 2007; Zhang et al., 2001). Mansfield and Freer-Smith (1984)는 0.07 ppm SO2에 노출된 백자작나무(Betula pendula)의 순광합성이 19%, 증산량이 46% 감소하는 것을 확인하였으며, 이는 수목이 대기오염물질에 대한 방어기작으로 잎의 기공을 닫음으로써 발생하는 현상임을 확인하였다. 이러한 단기간의 SO2 노출에 따른 기공 폐쇄 반응은 많은 연구에서 나타나고 있으나(Biggs and Davis, 1980; Kimmerer and Kozlowski, 1981), NO2의 영향에 따른 기공 폐쇄는 이보다 장기간, 고농도에서 발생하는 것으로 보인다(Gessler et al., 2000). 또한, Amundson and Weinstein(1981)과 Ashenden (1979)에서는 SO2와 NO2 혼합물에 노출된 식물의 경우 기공전도도가 감소하지만, 단일 NO2에 노출된 식물의 경우 기공전도도가 감소하지 않는 것을 확인하였다. 결과적으로 본 연구에서 SO2에 노출된 수목은 기공을 닫아 SO2의 흡수 속도가 시간이 지남에 따라 점차 감소하는 것으로 생각된다.
수목의 NO2 저감 능력과 생리적 특성의 관계를 파악하기 위해 잎의 광합성 색소 함량과 엽면적당 가스 저감량과의 상관관계를 종 평균 수준에서 분석하였다. 광합성 색소는 수목의 광합성을 위한 광수확 및 과도한 빛 에너지로부터 광계 보호, 기타 생장 기능에 중심 역할을 하며 많은 선행 연구에서 광합성 능력의 대용물로 여겨진다(Croft et al., 2017; Shah et al., 2017; Wang et al., 2020). 본 연구에서는 그중 가장 주요한 색소인 엽록소 a와 b, 카로티노이드 함량을 측정하였다(Figure 3).
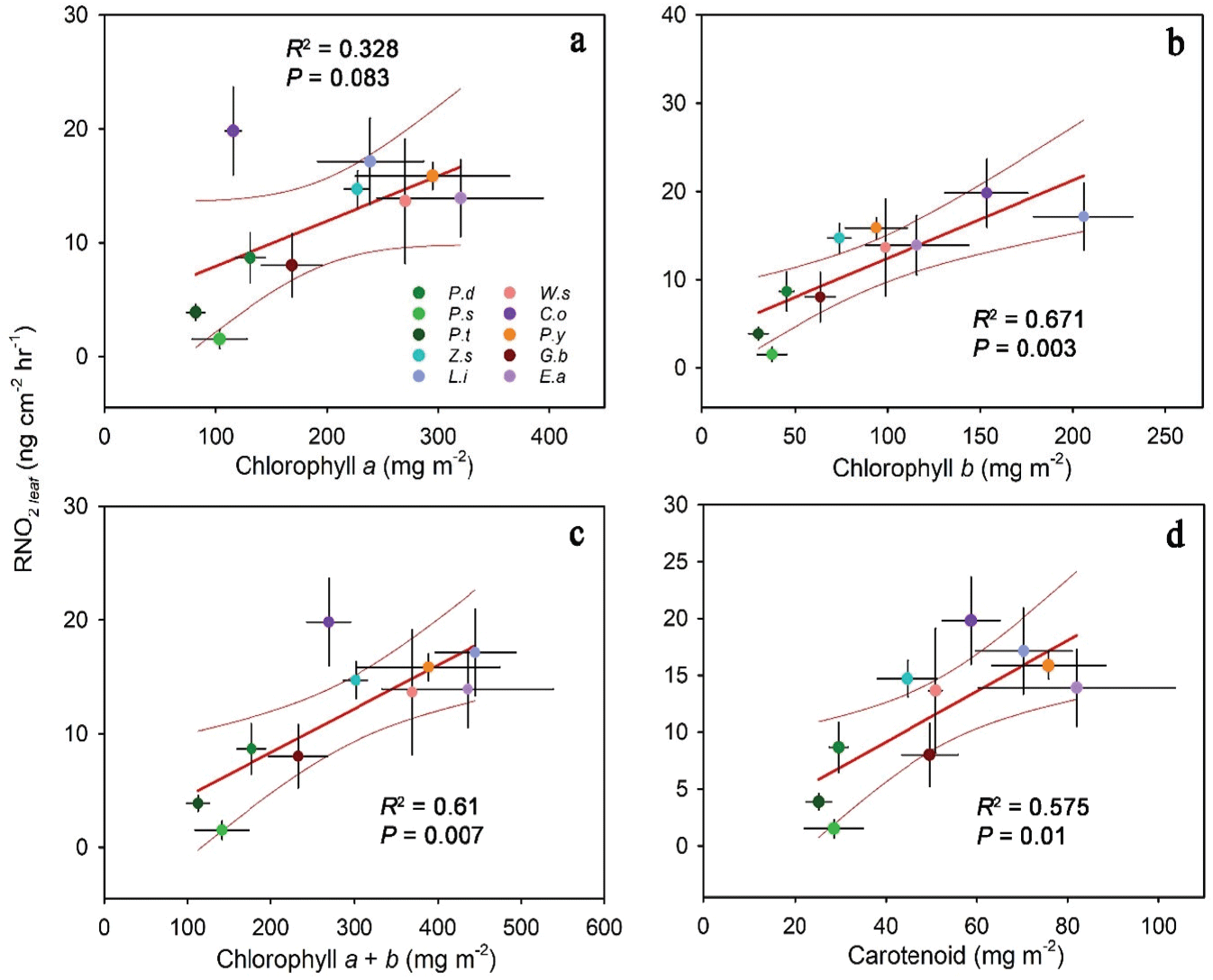
엽록소 a 함량은 엽면적당 NO2 저감량과양의 상관관계를 보였지만 통계적으로는 오차 수준 10%에서만 유의하였다(R2 = 0.328, P = 0.083, Figure 3a). 엽면적당 NO2 저감량이많은 배롱나무와 왕벚나무의 엽록소 a 함량은 각각 239.21 ± 47.74 mg m−2, 294.85 ± 69.55 mg m−2으로 높게 나타났고, 저감량이 적은 스트로브잣나무와 곰솔의 엽록소 a 함량은 각각 103.39 ± 24.33 mg m−2, 82.13 ± 8.11 mg m−2으로 낮게 나타났다. 엽록소 b 함량의 경우 엽면적당 NO2 저감량과 유의한 양의 상관관계를 보이며, 엽록소 a 함량에 비해 수목의 NO2 저감 능력에 밀접한 관계가 있는 것으로 나타났다(R2= 0.671, P = 0.003, Figure 3b). 엽면적당 NO2 저감량이 많은 산수유와 배롱나무의 엽록소 b 함량은 각각 153.48 ± 22.47 mg m−2, 205.81 ± 26.79 mg m−2으로 높게 나타났고, 저감량이 적은 스트로브잣나무와 곰솔의 엽록소 b 함량은 각각 37.58 ± 7.97 mg m−2, 30.27 ± 5.21 mg m−2으로 낮게 나타났다. 엽록소 a와 b의 합인 총 엽록소 함량 또한 엽면적당 NO2 저감량과 유의한 양의 상관관계를 보였다(R2 = 0.61, P = 0.007, Figure 3c). 엽면적당 NO2 저감량이 많은 배롱나무와 왕벚나무의 총 엽록소 함량은 각각 445.02 ± 48.64 mg m−2, 388.7 ± 86.09 mg m−2으로 높게 나타났고, 저감량이 적은 스트로브잣나무와 곰솔의 총 엽록소 함량은 각각 140.98 ± 32.21 mg m−2, 112.41 ± 13.30 mg m−2으로 낮게 나타났다. 예외로 엽면적당 NO2 저감량이 가장 많은 산수유의 경우 총 엽록소 함량은 269.03 ± 26.12 mg m−2으로 중간 정도로 나타났다. 엽록소와 마찬가지로 광합성 기작에 참여하는 카로티노이드 또한 수목의 NO2 저감 능력에 관계가 있는 것으로 나타났으며, 엽면적당 NO2 저감량과 유의한 양의 상관관계를 보였다(R2= 0.575, P = 0.01, Figure 3d). 엽면적당 NO2 저감량이 많은 산수유, 배롱나무, 왕벚나무의 카로티노이드 함량은 각각 58.71 ± 6.32 mg m−2, 70.31 ± 10.6 mg m−2, 75.77 ± 12.48 mg m−2으로 높게 나타났고, 저감량이 적은 스트로브잣나무와 곰솔의 카로티노이드 함량은 각각 28.65 ± 6.42 mg m−2, 25.34 ± 2.79 mg m−2으로 낮게 나타났다.
NO2에 노출된 수목의 경우 광합성 색소 함량이 높은 수목이 NO2 저감능력 또한 우수한 것으로 나타났다(Figure 3). 하지만 수목의 NO2 저감 능력이 광합성 색소 함량과 양의 상관관계를 보인 것과 달리, 수목의 SO2 저감 능력은 엽록소, 카로티노이드 함량과 유의한 상관관계를 나타내지않았는데(Table 2), 이는 SO2의 영향을 받은 수목의 기공 폐쇄 작용에 의한 것으로 생각된다(Biggs and Davis, 1980; Kimmerer & Kozlowski, 1981). 잎의 엽록소 파괴는 고농도의 오염원에 노출되었을 때 발생하는 반면(Joshi and Swami, 2009), 기공전도도의 감소는 저농도의오염원 노출에도 일어날 수 있다(Mansfield and Freer-Smith, 1984). 또한 기공의 폐쇄 작용은 NO2 보다 SO2에 더 민감하게 반응하는데(Biggs and Davis, 1980; Gessler et al., 2000; Kimmerer and Kozlowski, 1981), Bennett et al. (1990)에 따르면 SO2에 노출된 강낭콩(Phaseolus vulgaris L.)의 경우 0.17 ppm에서 처음 CO2 교환 속도의 저감이 관찰된 반면, NO2에 노출된 경우 0.38 ppm에서 CO2 교환 속도의 저감이 확인되었다. 본 연구에서 또한 SO2에 노출된 수목의 경우에만 기공 폐쇄 반응이 일어난 것으로 생각되는데(Figure 2a, c), 수목의 기공 폐쇄가 광합성 속도와 연관된 것을 고려한다면(Downton et al., 1988; Santos et al., 2018), 기공 폐쇄 반응이 광합성 색소 함량과 SO2 저감 능력이 상관관계를 보이지 않는 원인으로 생각된다.
수목의 수종별 NO2 저감능력을 설명하는 형태적 특성을 밝히기 위해 상관관계 분석을 실시하였다. 일반적으로 생각할 수 있는 수목의 총 엽면적과 NO2 저감량은 상관관계가 없는 것으로 나타났는데(P = 0.335, Table 1), 이는 Figure 2b에서 설명하는 것과 같이 단위 엽면적당 NO2 저감능력이 수종별로 차이가 커, 같은 엽면적의 나무도 수종에 따라 NO2 저감능력에 차이가 있기 때문이다(P = 0.004). 이를 통해 수목의 대기오염물질 능력은 잎의 생리적인 능력과 밀접한 관계를 가질 것으로 예상할 수 있다. 엽건중량 대비 엽면적을 나타내는 SLA는 엽면적당 NO2 저감량과 양의 상관관계를 보였지만 통계적으로는 오차 수준 10%에서만 유의한 것으로 나타났으며(R2 = 0.385, P = 0.055, Figure 4a), 엽면적당 NO2 저감량이 많은 배롱나무와 느티나무의 SLA는 각각 34.97 ± 0.84 m2 kg−1, 30.82 ± 2.22 m2 kg−1로 높게 나타난 반면, 저감량이 적은 스트로브잣나무와 곰솔의 SLA는 각각 9.24 ± 0.33 m2 kg−1, 7.97 ± 1.1 m2 kg−1로 낮은 값을 보였다. LMA는 잎의 두께를 설명하기 때문에(Vilar et al., 2013), SLA가 크다는 뜻은 잎의 두께가 얇은 것으로 생각할 수 있다. 즉 두꺼운 잎을 가진 수종에 비해 얇은 잎을 가진 수종의 NO2 저감능력이 뛰어난 것으로 확인되었다. 하지만, 예외로 엽면적당 NO2 저감량이 가장 많은 산수유의 경우 SLA는 중간 정도인 16.52 ± 0.15 m2 kg−1로 나타났다. 수목의 NO2 저감능력과 관계있는 형태적 특성 중 다른 하나는 엽면적당 둘레비(MLleaf)로 나타났다. 엽면적당둘레비는 개체목 수준에서 엽면적당 NO2 저감량과양의 상관관계를 보여, 엽면적 대비 엽둘레가큰 수목이 NO2 저감량이 많은 것으로 나타났다(Figure 4b). 상관관계는 이면기공엽 수종과 양면기공엽 수종 모두 유의하였으나 두 개의 다른 관계식에 의해 설명되었으며, 이면기공엽 수종에 비해 양면기공엽 수종에서특히 설명력이 높은 것으로 나타났다(각각 R2 = 0.211, P = 0.036과 R2 = 0.489, P = 0.035). 하지만 이러한 관계는 종 평균 수준에서는 유의하게 나타나지 않았다(각각 P = 0.823과 0.213).
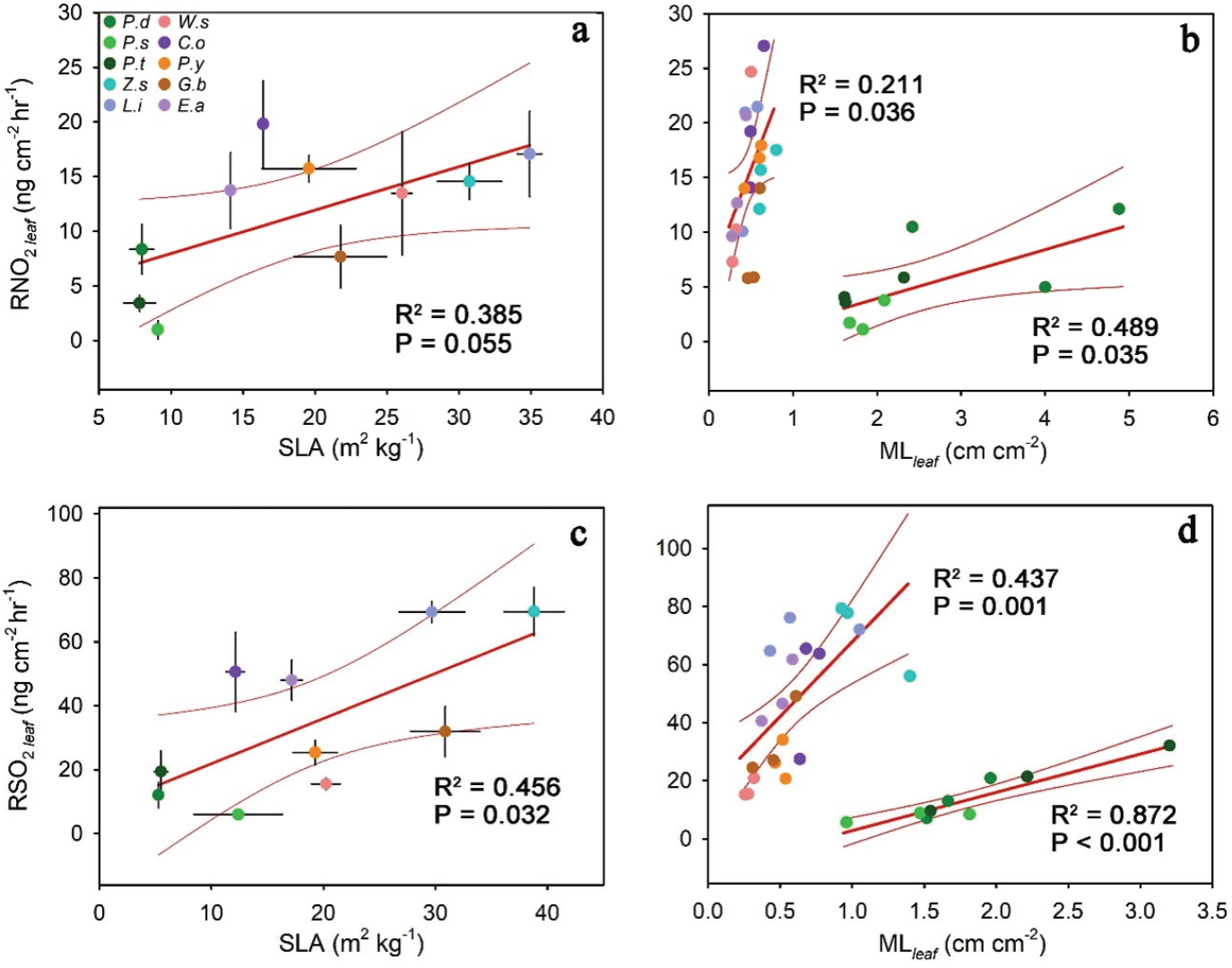
마찬가지로 수종별 SO2 저감능력 차이의 원인 구명을 위해 잎의 형태적 특성과 SO2 저감량과의 상관관계를 분석하였다. 수목의 총 엽면적과 SO2 저감량은 상관관계가 없는 것으로 나타났으며(P = 0.317, Table 1), 이는 NO2의 경우와 마찬가지로 단위 엽면적당 SO2 저감능력이 수종에 따라 차이가 크기 때문이다(Figure 2d, P < 0.001). 하지만, 엽면적당NO2 저감량이 SLA와 오차 수준 10%에서만 유의한 상관관계를 보인 것과 달리, 엽면적당 SO2 저감량은 SLA와 오차 수준 5% 미만에서 유의하였으며 더 높은 상관계수를 보였다(R2 = 0.456, P = 0.032, Figure 4c). 엽면적당 SO2 저감량이 많은 느티나무와 배롱나무의 SLA는 각각 38.66 ± 2.69 m2 kg−1, 29.5 ± 2.95 m2 kg−1로 높게 나타난 반면, 저감량이적은 스트로브잣나무, 소나무, 곰솔의 SLA는 각각 12.14 ± 3.95 m2 kg−1, 4.99 ± 0.21 m2 kg−1, 5.22 ± 0.61 m2 kg−1로 낮게 나타났다. 개체목 간의 저감량 차이는 엽면적당 둘레비와의 상관관계로 잘 설명되었으며 엽면적 대비 엽둘레가 큰 개체목의엽면적당 SO2 저감량이 많은 것으로 나타났다(Figure 4d). 상관관계는 NO2의 경우와 마찬가지로 이면기공엽과 양면기공엽 두 개의 관계식으로 나누어졌지만, 설명력은 두 관계식 모두 SO2에서 높은 것으로 나타났다(각각 R2= 0.437, P = 0.001과 R2 = 0.872, P < 0.001). 또한 SO2 저감량과 엽둘레비의 상관관계는 개체목 수준뿐만아니라 종 평균 수준에서도 유의하게 나타나, 저감량이 가장 많은 느티나무의 엽면적당 둘레비는 1.08 ± 0.15 cm cm−2로 가장 높았고, 저감량이 가장 적은 병꽃나무의 둘레비는0.26 ± 0.01 cm cm−2로 가장 낮은 값을 가졌다(R2 = 0.7, P = 0.018).
수목의 NO2와 SO2 저감 능력은 잎의 형태적 특성 중 SLA에 의해 설명되어, 두 경우 모두 두꺼운 잎을 가진 수종 보다 얇은 잎을 가진 수종의 저감능력이 뛰어난 것으로 나타났다(Figure 4a, c). Bennett and Hill(1973)에 의하면 가스 형태의 오염물질은 잎의 기공, 상처 부위, 엽육의 자유 공간에서 확산 작용을 통해 잎의 내부로 이동한다. 특히 두꺼운 잎보다는 얇은 잎에서 오염물질 저감이 유리한데, 이는 두꺼운 잎에서는 엽육의 자유 공간 감소로 인해 엽육 전도도(mesophyll conductance)가 감소하는 반면, 얇은 잎은 상대적으로 원활한 확산 작용을 하기 때문이다(Clarke et al., 2021; Terashima and Ono, 2002; Veromann-Jürgenson et al., 2017). 저감 능력의 차이를 설명하는 형태적 특성 중 다른 하나는 엽면적당 둘레비로 확인되었으며, 엽면적 대비 엽둘레가 큰 수목의 저감능력이 뛰어난 것으로 나타났다(Figure 4b, d) (Chakre, 2006; Sgrigna et al., 2020). 엽면적 대비 엽둘레가 크다는 뜻은 결각이 크다는 뜻이며, 이에 따라 대기와 잎 사이의 가스와 에너지를 교환하는 능력인 경계층 전도도(boundary layer conductance)가 높게 나타나는 것으로 생각된다. 경계층 전도도가 높을수록 잎의 증발산량 또한 많아지며(Martin et al., 1999), 이는 챔버내 상대습도를 증가시켜 NO2와 SO2 같은 수용성 물질의 습성침착(wet deposition)을 가속화한다(Cho and Choi, 2009; Ryu et al., 2019). 결과적으로 대기오염물질 저감능력이 엽둘레비와 상관관계를 보이는 것은 증발산의 영향이 주요한 것으로 생각된다.
Figure 4b와 d에서 이면기공엽 수종과 양면기공엽 수종은 각각 다른 관계식에 의해 설명되었는데, 양면기공엽 수종의 경우 침 모양의 잎으로 인해 결각이 잘 발달하지 않음에도 불구하고 잎의 크기가 작아 엽둘레비가 크게 나타났다. 하지만 양면기공엽 수종은 큰 엽둘레비에도 불구하고 이면기공엽 수종에 비해 NO2 및 SO2 저감량이 적었는데, 이는 둘의 생리적 활성 차이에 의해 설명된다. 이면기공엽 수종은 생리적 활성과 광합성 능력을 나타내는 엽록소 함량이 양면기공엽 수종보다 높게 나타났고, 이에 광합성에 따른 기공의 흡수 기작으로 인한 대기오염물질 저감의 차이가 크기 때문에 둘의 엽둘레비와 NO2 및 SO2 저감량 사이의 관계식이 두 개로 나누어진 것으로 생각된다. 같은 맥락에서 산수유는 중간 정도의 SLA 값을 가짐에도 불구하고 전체 수종 중 가장 높은 NO2 저감 능력을 보이는데, 이는 높은 엽면적당 엽둘레비에 의한 경계층 전도도의 영향으로 생각된다.
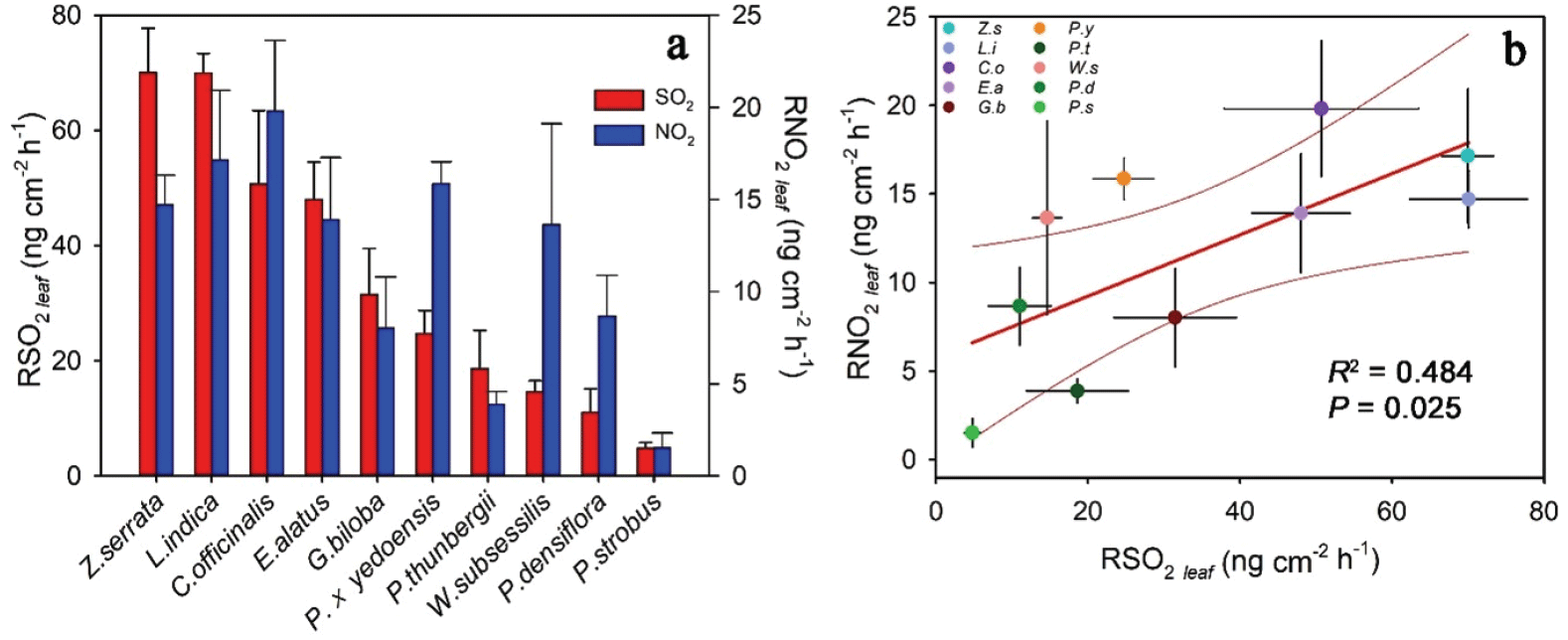
수목의 NO2 및 SO2 저감능력은 수종별로 유의하게 달랐는데, 이면기공엽을 갖는 활엽수종과 은행나무가 양면기공엽을 갖는 침엽수종보다 저감능력이 뛰어난 것으로 나타났다(Figure 2). 이는 엽면적당 광합성능과 기공전도도로 인해 활엽수종이 침엽수종보다, 낙엽수종이 상록수종보다 NO2와 SO2 저감능력이 뛰어나다고 하는 선행 연구들과 같은 결과이다(Jo andAhn, 2001; Okano et al., 1989; Takahashi et al., 2005). 수종별 엽면적당 NO2 저감량과 SO2 저감량은 Figure 5a와 같이 나타났는데, 오염물질의 종류에 따라 저감 능력은 다소 차이가 있는 것으로 나타났다. 이는 앞서 설명한 것과 같이 NO2 저감능력의 경우 광합성(광합성 색소)의 영향을 크게 받으며, SO2 저감능력의 경우 잎의 두께(SLA)와 증발산(엽둘레비)의 영향을 크게 받기 때문으로 생각된다(Figure 2, 3). 하지만 그럼에도 불구하고 상관 분석 결과 NO2 저감량과 SO2 저감량은 유의한 양의 상관관계를 보였는데(R2 = 0.484, P = 0.025, Figure 5b), 이를 통해 수목의 오염물질 흡수 능력이 광합성, 증발산, 기공전도도, 잎의 두께와 같은 수종별 특성과 연관되며, 이러한 특성이 우수한 수종은 두 가지 오염물질의 저감 능력이 상대적으로 뛰어나기 때문에 나타나는 결과로 생각된다. 본 연구에서 두 가지 오염물질의 저감능력이 모두 우수한 수종은 느티나무, 배롱나무, 산수유로 나타났으며, 스트로브잣나무, 곰솔, 소나무의 경우 두 가지 오염물질의 저감능력이 모두 저조한 것으로 나타났다.
수목의 NO2 저감능력과 SO2 저감능력을 가장 잘 설명하는 특성의 조합을 찾고 비교하기 위해 앞서 분석했던 잎의 생리적, 형태적 특성을 독립변수로, 엽면적당 NO2 및 SO2 저감량을종속변수로 하여 단계적 다중 회귀분석을 실시하였다. 분석 결과 수종별 NO2 저감능력은 잎의 생리적 특성 중 엽록소 b 함량에 의해 가장 잘 설명되었으며, 수종간 차이의 67.1%를 설명하는 것으로 나타났다(P = 0.003, Table 3). 하지만 NO2의 경우 이외의 다른 요인을 추가하더라도 설명력이 증가하지 않는 것으로 나타났다. 반면, 수종별 SO2 저감능력은 잎의 형태적 특성 중 SLA에 의해 가장 잘 설명되어, 수종간 차이의 45.6%를 설명하였다(P = 0.032). 단계적 선택 과정에서 엽면적당엽둘레비와 엽록소 b 함량은 함께 추가되어 수종간 차이의 설명력을 64.8%로 개선하였으며(P = 0.081), 이를 통해 잎의 두께만으로는 설명력이 부족했던 수종간 차이가 증발산과 광합성의 영향을 받는 것으로 생각되었다. 다만 SO2의 경우 기공 폐쇄에 의한 영향으로 광합성 색소 함량과의 정확한 상관관계는 파악할 수 없어, 잎의 두께가 가장 주요한 변수인지에 대해서는 추가적인 연구가 필요할 것으로 생각되며, 추후 수목의 광합성 능력, 증발산량, 기공전도도를 직접 측정하는 것이 고려된다.
| Gas types | Explanatory variables | R 2 | P |
|---|---|---|---|
| NO2 | Chl b | 0.671** | 0.003 |
| SO2 | SLA | 0.456* | 0.032 |
| Chl b, MLleaf | 0.648 | 0.081 |
결 론
대표적인 조경수종들의 NO2와 SO2의 저감 능력을 정량화한 결과, 저감 능력이 우수한 수종으로는 느티나무, 배롱나무, 산수유로 나타났으며, 저조한 수종으로는 스트로브잣나무, 곰솔, 소나무로 확인되었다. 또한 수목의 NO2 및 SO2 저감 기작은 잎의 생리적, 형태적 특성의 영향을 받는 것으로 나타났다. 수목의 NO2 저감 기작은 광합성 색소와 SLA의 영향을 받았으며, SO2 저감 기작은 SLA, 엽면적당엽둘레비와 관계 있는 것으로 나타났다. 이러한 수목의 특성은 광합성, 증발산, 기공전도도와 연관되어 대기오염물질의 저감 능력을 나타낸 것으로 생각된다. 이러한 결과로부터 가로수 또는 도시림에 있어 NO2 및 SO2 저감 능력이 우수한 수종 선정과 더불어, 식재목이 원활한 생리 활동을 할 수 있도록 생육 상태를 관리하는 것이 도시 지역의 심해지는 대기오염물질 저감에 도움이 될 것이라 생각된다. 본 연구는 잎의 생리적, 형태적 특성을 통한 단기간의 대기오염물질 저감 능력만을 고려하였지만 실제 도시 지역의 수목 선정 및 관리에 있어서는 대기오염물질에 장기간 노출되었을 때의 저감 능력 및 병충해 저항성 등의 종합적인 능력이 연구되어야 하며, 이를 통해 도시인들의 건강에 도움이 되는 조경수 선정이 완성될 것으로 사료된다.