Introduction
Bamboo forests have been the focus of research because of the role of atmospheric carbon dioxide sequestration through rapid growth and a number of potential ecosystem services, besides being a traditional source of energy and other multiple uses (Isagi, 1994; Nath et al., 2009; Nath et al., 2015). Thus, bamboo forests are becoming an important resource for meeting the future demand for bamboo production and global climate change. The nutrient status of bamboo tissues and soils has been frequently used to evaluate bamboo growth and production (Embaye et al., 2005; Tang et al., 2016; Kim et al., 2018; Sun et al., 2019).
The role and importance of the nutrient status of bamboo tissues and soils are likely to be quite variable among different bamboo species because different nutrient requirements and allocation mechanisms are affected by different habitat and nutrient conditions. Although such information is useful for assessing the potential impacts of nutrient limitation based on bamboo tissues, few studies have assessed the nutrient relationship between leaves, roots, soils, and different bamboo species at a regional scale. In addition, foliar nutrients have been widely used as indicators of plant nutritional status, while nutrient concentrations in roots have been less studied (Alvarez-Clare and Mack, 2015).
Approximately 70 species of bamboo occur either naturally or artificially in the warm temperate forest zones of Korea (Lee et al., 2012). Three bamboo species (Phyllostachys bambusoides Sieb. et Zucc, P. nigra var. henonis Stapf ex. Rendle, and P. pubescens Ohwi) were intensively planted for edible shoots and multipurpose use, such as energy or timber production in southern Korea (Park and Ryu, 1996; Yoo et al., 2017). There is a need to evaluate different bamboo species for nutrient status to establish species-specific nutrient management systems (Zhang et al., 2014). Although many studies have evaluated the effect of bamboo species on biomass production and nutrients in different plant parts of bamboo forests on a stand scale (Park and Ryu, 1996; Hwang et al., 2005; Park et al., 2017), major uncertainties remain in the behavior of nutrient status of bamboo tissues and soils for nutrient diagnose by different bamboo species on a regional scale. The objective of this study was to evaluate the nutrient concentrations of leaves, roots, and soils from three bamboo species on a broad regional scale. In this study, we determined that the nutrient concentrations in bamboo stands may be useful for diagnosing nutrient conditions.
Materials and Methods
The study sites were located in three regions (Jinju-si, Damyang-gun, and Sacheon-si) for P. nigra var. henonis, five regions (Jinju-si, Goseong-gun, Damyang-gun, Sacheon-si, and Hadong-gun) for P. bambusoides, and three regions (Sacheon-si, Geojae-si, and Jinju-si) for P. pubescens stands in southern Korea (Figure 1, Table 1). The mean annual mean precipitation and temperature of 30 years are 1,556 mm yr−1 and 12.8°C for Sancheong-gun, 1,513 mm yr−1 and 13.1°C for Jinju-si, and 2,007 mm yr−1 and 14.2°C for Geojae-si, respectively (Korea Meteorological Administration, 2017). The soils are well-drained, slightly wet or dry, brown forest soil originating from granite or granite gneiss from Hadong-gun and Geojae-si, with a loamy texture. Whereas, the soils in Goseong-gun, Sacheon-si, and Jinju-si are slightly dry, dark reddish-brown forest soils originating from sandstone or shale.
Values in parenthesis are standard error. Different letters among bamboo species represent a significant difference among bamboo stands at P < 0.05.

The experimental design consisted of a completely randomized design involving three plots of 5 × 5 m in each bamboo stand at each site. Most bamboo stands in the study sites have been unmanaged and abandoned. The plots were established in the middle of the stands to reduce edge effects. The diameter at breast height (DBH) of all bamboo was measured at 1.2 m height in June 2017. All DBH in each plot were measured to estimate the bamboo biomass from the allometric regressions developed by these three bamboo species (Jung et al., 2017). The bamboo biomass was converted to organic C using the C concentration of the three bamboo species (Park et al., 2017).
Leaf samples from mature and healthy bamboo were obtained from approximately 5 m in height using a tree pruner in June 2017. Leaf samples were transported to a laboratory and dried to a constant mass at 65°C for one week. To obtain fine root samples, three cores were taken using 405 cm3 soil sampling cores (7.04 cm inner diameter, 10.4 cm height) up to a soil depth of 10 cm, from each treatment plot. All roots were transported to a laboratory and separated carefully by washing with tap water to remove the mineral soil. The separated roots were dried to a constant weight at 65°C for one week. The dried leaf and root samples of each bamboo species were ground in a Wiley mill and passed through a 40-mesh stainless-steel sieve. Carbon and N concentrations from the ground materials were determined using an elemental analyzer (Thermo Fisher Scientific Flash 2000, Milan, Italy). After dry ashing the ground material (0.5 g) at 470°C for 4 h, the ash was digested with 3 mL of concentrated 5 M HCl with 0.25 mL of concentrated HNO3. Macronutrient (P, K, Ca, and Mg) concentrations from the digest were determined using ICP-OES (Perkin Elmer Optima 5300 DV, Shelton CT, USA).
Soil samples from each plot were collected from three randomly selected points during leaf sampling. Soil samples from each treatment plot were collected at three locations at soil depths of 0–10 cm using 405 cm3 soil sampling cores by a core sampling technique (Xu et al., 2016) after removing the organic horizons. Soil samples were transported to a laboratory and sieved through a 2 mm mesh sieve. Soil C and N concentrations were determined using an elemental analyzer (Vario Macro cube, Langenselbold, Germany). The available soil P concentration extracted by NH4F and HCl solutions was determined using a UV spectrophotometer (Jenway 6505, Staffordshire, UK). Exchangeable cation (K+, Ca2+, Mg2+) concentrations extracted by NH4Cl solution (Kalra and Maynard, 1991) with a mechanical vacuum extractor (Model 24 VE, SampleTeck, Science Hill, KY, USA) were determined using ICP-OES (Perkin Elmer Optima 5300 DV, Shelton, CT, USA).
The nutrient concentrations of leaves, roots, and soils were analyzed by one-way analysis of variance (ANOVA) to determine the significance of bamboo species (P<0.05) (SAS Institute Inc., 2003). A significant difference among the three bamboo species was performed by applying Tukey’s multiple comparison test at P<0.05.
Results and Discussion
The density of bamboo culms was highest in P. nigra var. henonis (8,640 culm ha−1) stands, followed by P. bambusoides (6,800 culm ha−1), and P. pubescens (5,680 culm ha−1) stands. The mean DBH was significantly higher in P. pubescens (8.45 cm) with big growth characteristics than in P. nigra var. henonis (4.97 cm) or P. bambusoides (4.37 cm) stands (Table 1). The aboveground biomass C stocks were highest in P. pubescens (28.30 Mg C ha−1), followed by P. nigra var. henonis (19.29 Mg C ha−1) and P. bambusoides (12.04 Mg C ha−1) stands. This result indicated that the biomass C stocks in the three bamboo species were determined by DBH rather than by culm density. In this study, C stocks in three bamboo species were slightly higher or lower than the 25.26 Mg C ha−1 for P. pubescence, 25.07 Mg C ha−1 for P. bambusoides, and 13.93 Mg C ha−1 for P. nigra var. henonis stands reported by Jung et al. (2017).
Species-specific differences were observed in the C and Ca concentrations of leaves, and in N and Mg of roots among the three bamboo species (Figure 2). The C concentration in bamboo tissues could be affected by inter-and intra-specific variation, as determined by the interaction of genetic and environmental factors (Bert and Danjon, 2006; Thomas and Martin, 2012). In addition, the C concentration in tree tissues was significantly correlated with the nutrients in tree tissues (Tang et al., 2018). The low C concentration in the leaves of P. bambusoides could be due to the high Ca concentration in the leaves of P. bambusoides, compared to the other two bamboo species. Similar results were reported by Tang et al. (2018), who reported that low C concentrations in leaves were due to high leaf nutrient concentrations. The mean C concentrations (420–434 g C kg−1) in the leaf of the three bamboo species were similar to the global mean value of 430 g C kg−1 for the C concentration of bamboo leaves (Ma et al., 2018). In contrast to leaf C concentration, the C concentration in the roots was not significantly different among the three bamboo species. The mean C concentration in the roots of the three bamboo species was 426 g C kg−1, which was slightly lower than the global mean value of 451 g C kg−1 for the C concentration of bamboo roots (Ma et al., 2018). Calcium concentrations in the leaves were significantly lower in P. pubescens (7.83 g Ca kg−1) than in P. bambusoides (11.94 g Ca kg−1), whereas the K concentration was lowest in P. bambusoides. The lowest K concentration in the leaves of P.bambusoides could be associated with the competition between K+ and Ca2+ at the cellular level. Similarly, Kim et al. (2018) reported a strong antagonism between K+ and Ca2+ or Mg2+ uptake in fertilized bamboo stands. In addition, the low concentrations of soil N and K+ in P. bambusoides are characterized by fast growth, and higher uptake and storage of nutrients in biomass (Kaushal et al., 2020).
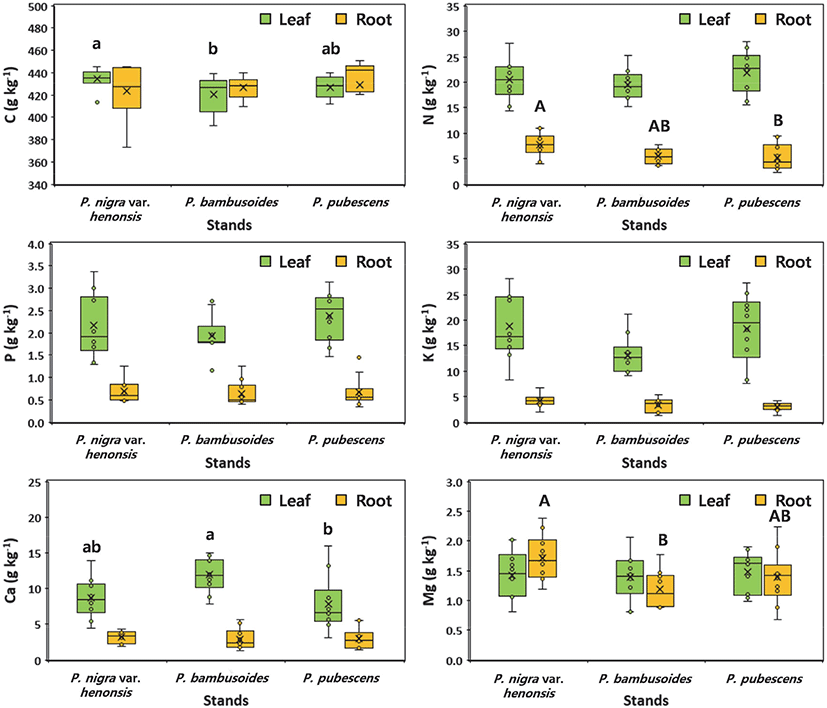
Nitrogen concentrations in the roots were significantly lower in P. pubescens (5.23 g N kg−1) than in P. nigra var. henonis (7.72 g N kg−1). This result could be due to dilution effects caused by the differences in biomass between P. pubescens and the other two bamboo species (Figure 3). In addition, a negative correlation (r = –0.88) was found between the N concentration of culm and culm biomass in P. bambusoides (Park et al., 2017). In contrast to the N concentration, the Mg concentration in the roots was significantly higher in P. nigra var. henonis (1.72 g Mg kg−1) than in P. bambusoides (1.25 g Mg kg−1). The nutrient (N, P, K, and Ca) concentrations in P. nigra var. henonis were generally higher than those of the other two bamboo species. This result suggests that more nutrients could be allocated to roots to increase the uptake capacity of soil resources in high culm densities in P. nigra var. henonis stands. In contrast to roots, the low nutrient concentration of leaves in this bamboo species could be due to high culm density.
Mean N, P, and K concentrations of the three bamboo species in this study were 20.71 g N kg−1, 2.17 g P kg−1 and 16.71 g K kg−1 for leaves; and 6.15 g N kg−1, 0.67 g P kg−1, and 3.57 g K kg−1 for roots, respectively. The nutrient concentrations (N, P, K, and Ca) in bamboo tissues were generally higher in the actively growing parts of bamboo (e.g., leaves) than in the structural growing parts (e.g., roots). The nutrient concentrations in this study were comparable to the range of values reported by Kleinhenz and Midmore (2001). They reported that the mean N, P, and K concentrations of bamboo forests were 22.7 g N kg−1, 3.9 g P kg−1, and 19.2 g K kg−1 for leaves; and 7.40 g N kg−1, 0.95 g P kg−1, and 4.20 g K kg−1 for roots, respectively. However, the N concentration in leaves was considerably lower than the value of 30 mg N kg−1 recommended by Kleinhenz et al. (2003) for optimal yield. In addition, the N concentration in leaves was similar to the N concentration (19.8 mg N kg−1) in leaves of unfertilized P. pubescens stands reported by Kim et al. (2018). The results suggest that the low N concentrations of bamboo tissues in the three different bamboo stands could be due to the increase of unmanaged bamboo stands.
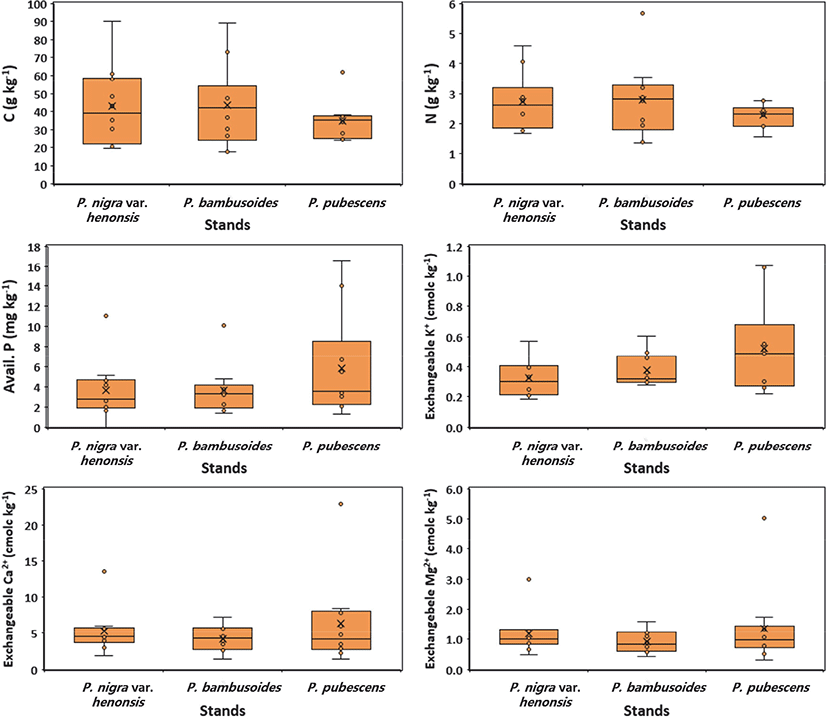
Soil nutrients, such as total N, organic C, and exchangeable Ca2+ and Mg2+, were not significantly affected by the bamboo species. Kaushal et al. (2020) observed similar results that did not reveal any significant changes in organic C and available P among different bamboo species in India. Although soil nutrient concentrations among different bamboo species may be largely responsible for the differences in litterfall, decomposition rate, rhizome or root distribution, nutrient requirement, nutrient uptake, and nutrient allocation throughout bamboo stand development (Kleinhenz and Midmore, 2001), a high degree of soil heterogeneity at broad regional scales may be associated with high variation in soil chemistry (Figure 3) among different bamboo species.
Conclusions
The nutrient concentrations in bamboo tissues were likely to be affected by species-specific variation among bamboo. In addition, roots may be useful for diagnosing N and Mg conditions among different bamboo species. However, soil nutrients were not affected by different bamboo species. The results indicate that the nutrient concentration of bamboo tissues was affected more by species-specific variation than by soil nutrients.