서 론
도서(섬)지역은 육지와의 지리적 격리성(isolation)과 해양 환경의 영향으로 생태적 특이성과 온전성 그리고 생물적 고유성과 잠재유전 자원적 가치도 높지만, 비교적 생태계가 연속되어 있는 내륙지역보다 외부 교란에 대한 생태적 저항력(resistance)이나 회복력(resilience)이 크게 떨어지는 취약성도 동시에 가지고 있다. 이에 최근 국내외적으로 도서지역 자연생태계와 생물다양성의 기반을 이루는 산림생태계에 대한 체계적 보전 및 관리에 대한 중요성이 한층 높아지고 있다(Berg et al., 2014; van Andel et al., 2016; Kueffer and Kinney, 2017). 미국, 캐나다 등에서는 도서지역 산림생태계의 단편화와 구성요소의 단순화, 고유 생물종의 서식지와 개체군의 크기 감소 및 유성번식력 저하 등에 따른 생물다양성 감소 원인을 규명하기 위한 보전생태학적 연구가 활발히 수행되고 있으며(Olson et al., 2009; Federman et al., 2014; Hanson and McElroy, 2015), 일본에서도 도서지역 산림생태계에 있어서 상록수림을 비롯한 자연식생의 고립화가 생물다양성에 미치는 영향을 파악하여, 도서지역 산림생태계 특유의 생물다양성 요소를 생태학적으로 보전하고 복원하는 방안을 모색하는 여러 연구를 수행하고 있다(Tsuyoshi, 2010; Enoki et al., 2014; Hiroya et al., 2015). 또한 도서의 식물종다양성과 지리적 조건과의 상관성을 분석하여 해발고도와 식물종수가 유의미한 상관관계에 있는 것을 밝혔다(Choi, 2004; Kim et al., 2017). 식물군락은 동일 장소에서 함께 생활하는 식물 집단을 가리키는 편의적인 식생단위로서, 그 크기와 범위에 대한 특별한 규정은 없으며, 대개 같은 입지에서는 상관, 구조, 조성 등이 비슷한 집단으로 분류 된다(Kim et al., 2016). 이런 점을 감안하여 생태학이 발달한 국가에서는 자국 산림생태계의 생태 및 생물 정보를 체계화하거나 생태적 관리 및 보전 계획 수립의 기초 단위로서 식물군락을 널리 활용하고 있다(Arponen et al., 2008; Ren et al., 2012).
본 연구 대상지인 욕지도는 식물구계지리학적으로 남해안아구에 속하고 식생은 난대 또는 난온대산림대에 해당하지만(Yim and Kira, 1975; Lee and Yang, 1978), 해방이후 국가 주도의 수종갱신사업, 임지전용, 산불 등 다양한 교란의 영향으로 고유 상록활엽수림을 비롯한 자연식생은 거의 파괴되어 소규모 패치상으로 드물게 분포하고 있다. 반면, 곰솔과 사방오리 등 대상식생(substitutional vegetation)이 광역으로 분포하고 있어 난온대림으로서의 생태적 특성과 생물적 고유성 회복이 시급한 상태이다. 욕지도 산림 식생에 관한 주요 선행연구로는 Kim(1997)의 ‘욕지도 식생에 관한 생태학적 연구’를 비롯하여, Oh et al.(1999)의 ‘한려해상국립공원 통영지구의 현존식생 및 구조’, Jung et al.(2008)의 욕지도 내 모밀잣밤나무군락의 종조성 및 군락구조, 그리고 Kim et al.(2016)의 ‘욕지도(통영시)의 식물다양성과 식생’ 등이 있으며, Kim(1997)과 Kim et al.(2016)은 욕지도 산림식생을 상관적 기준으로 모두 곰솔군락, 사방오리군락 등을 포함하여 각각 7개 유형, 19개 유형으로 구분하였고, 특히 Kim et al.(2016)은 구분된 상관식생의 공간분포에 의거하여 현존식생도를 작성한 바 있다. 그러나 이들 선행연구는 대부분 상관을 기준으로 산림식생을 구분하는데 그쳐 욕지도 산림생태계의 체계적 보전 및 관리 계획 수립의 기본단위로서의 잠재적 활용성이 높은 종조성적 산림식생 유형과 그 생태적 특성에 대한 정보는 아직 없는 실정이다. 이에 본 연구는 우리나라 도서지역 산림생태계의 생태적 기능과 생물적 가치 제고의 기반 조성을 위하여 산림식생을 유형화하고 그 생태적 특성과 공간분포 특성을 파악하고자 한다.
재료 및 방법
욕지도는 한반도 동남부 남해안의 끝에 위치하며(Figure 1), 연화도, 두미도, 우도 등 10개의 유인도와 45개의 무인도가 있는 경상남도 통영시 욕지면의 모도(mother island)로 섬 크기 15 km2의 남해안에서 세 번째로 큰 섬이다(Moon and Sohn, 2013). 주요 산봉우리는 최고봉인 천황산(394 m)을 비롯하여 대기봉(355 m), 약과봉(315 m), 망대봉(205 m), 일출봉(190 m) 등이 있다. 기후적으로는 온대해양성 기후 특성을 나타내며 한서의 차이가 심하지 않고, 최근 30년간(1990~2020년) 연평균 기온은 14.5°C, 연평균 강수량은 1,495.2 mm, 온량지수(WI)는 117.2, 한랭지수(CI)는 –3.6이다(Korean Meterological Administration, 2021). 지질은 중생대 백악기의 유천층군에 속하며 하부로부터 화산력응회암, 용결응회암, 석영안산암, 화강섬록암으로 구성되어 있고, 후기의 다양한 암맥이 관입하고 있다(Ahn, 2013).

현지조사는 2018년 5월부터 8월 사이에 임분 크기와 입지환경 특성을 고려하여 선정된 총 134개 정방형 조사구(100 m2 또는 400 m2)에서 수행되었다. 식생조사와 식생유형 분류는 서식처 환경조건과 식물사회 종조성의 균질성과 대응성을 강조하는 Z-M학파의 식물사회학적 방법을 따랐다(Braun-Blanquet, 1965; Müller-Dombois and Ellenberg, 1974). 식생유형 간 구성종의 중요치(importance value: IV)는 Curtis and McIntosh(1951)의 평가 기준을 사용하였다. 평균상대중요치(mean relative importance value: MIV)는 수관층위별로 가중치를 부여하여 교목층 IV×3+아교목층×2+관목층×1+초본층×0.5)/6.5로 산출하였다(Kim et al., 2018). 종다양성 지수는 자연로그 기법을 바탕으로 하여 Shannon의 종풍부도(Species richness index: S), 종다양도(Shannon diversity index: H′), 종균재도(Pielou evenness index: J′), 그리고 종우점도(Simpson dominance index: D) 등을 각각 계산하였다(Shannon and Weaver, 1949; Brower and Zar, 1977). 구성종의 총피도에 의한 상대적 우점순위를 가늠하기 위해 해당 식물군집의 출현종에 대한 양적 및 질적 평가를 통합한 지수인 상대기여도(r-NCD: Kim and Manyko, 1994)를 산출하였다(Kim and Lee, 2006).
Σ Ci: 군집 내의 i종의 피도 적산값, N: 전체조사구 수, ni: i종이 출현한 조사구수, NCDi: 대상식물군집에 대한 i종의 기여도, NCDmax: 대상군락 내의 기여도 최대값을 각각 나타낸다. 또한, 각 식생유형과 입지환경, 총피도 및 종다양성 지수 사이의 차이를 비교하기 위해 ANOVA 분석(Zar, 1984)을 수행함과 아울러, 해발고도와 전체 조사구(134개 조사구)의 종다양성 지수 및 총피도 사이의 관계를 파악하기 위해 상관분석을 수행하였다(de Bello et al., 2006). 현존식생도는 기존에 조사되었던 상관식생만 구분하는 것이 아니라 현지조사를 통하여 상관식생유형을 동정하고 그 경계를 확정한 후, QGIS Desktop 2.18.15 프로그램(QGIS Development Team, 2017)을 이용하여 공간정보화하였다. 연구 수행 중 식물동정은 원색식물도감(Lee, 2003)을 참조하였으며, 그 명명은 국가표준식물목록(Korea National Arboretum, 2021)을 기준으로 적용하였다.
결과 및 고찰
욕지도 산림식생 유형을 전통적인 Z-M학파의 식물사회학적 방법으로 분류한 결과, 총 8개 유형이 구분되었고, 그 식생단위 체계는 2개 군락(community), 4개 아군락(subcommunity), 6개 변군락(variant), 그리고 2개 아변군락(subvariant)으로 구성되어 있었다(Table 1). 이들 유형은 인간간섭, 입지환경, 성립기간 그리고 천이단계 등을 반영하여 종조성적 차이를 나타내고 있었다. 최상위 식생단위인 군락 수준에서는 종군 1의 구성종인 찔레꽃, 예덕나무, 사방오리, 장딸기, 두루미천남성 등이 강한 식별종으로 특징짓는 예덕나무-찔레꽃군락(Rosa multiflora-Mallotus japonicus community, Table 1: Vegetation unit Ⅰ; Vegetation type codes VT1~VT4)과 그 대립 종군인 종군 6의 구성종인 상록만경성의 마삭줄이 강한 식별종으로 특징짓는 마삭줄군락(Trachelospermum asiaticum var. intermedium community, Table 1: Ⅱ; VT5~VT8)의 2개 단위로 크게 구분되었고, 전자는 대개 사면중부이하에서, 후자는 대개 사면중부 이상에서 분포하고 있었다. 예덕나무-찔레꽃군락은 다시 종군 2의 사위질빵, 팽나무, 산괴불주머니 등이 강한 식별종으로 나타나는 팽나무-사위질빵아군락(Clematis apiifolia-Celtis sinensis subcommunity, Table 1: Ⅰ-A; VT1~ VT2)과 하위 식별종군을 갖지 않는 전형아군락(Typical subcommunity, Table 1: Ⅰ-B; VT3~VT4)의 2개 상위 단위로 구분되었으며, 전자는 다시 종군 3의 산쑥, 종군 8의 굴피나무, 그리고 종군 9의 멍석딸기 등이 강한 식별종으로 특징짓고 굴피나무, 예덕나무, 느티나무 등이 상층 우점종으로 나타나는 굴피나무-산쑥변군락(Artemisia montana-Platycarya strobilacea variant, Table 1: Ⅰ-A-1; VT1)과 하위 식별종군을 갖지 않으며 대개 사방오리가 상층 우점종으로 나타나는 전형변군락(Typical variant, Table 1: Ⅰ-A-2; VT2)의 2개 하위 단위로, 후자는 다시 종군 4와 그 대립종군인 종군 5의 구성종 차이에 의해 죽림인 왕대가 특징짓는 왕대변군락(Phyllostachys bambusoides variant, Table 1: Ⅰ-B-1; VT3)과 사방오리와 곰솔이 상층 우점종으로 나타나는 비목나무-줄딸기변군락(Rubus oldhamii-Lindera erythrocarpa variant, Table 1: Ⅰ-B-2; VT4)의 2개 하위 단위로 최종 구분되었다. 한편, 마삭줄군락은 종군 7과 그 대립종군인 종군 12의 구성종 차이에 의해 곰솔-청미래덩굴아군락(Smilax china-Pinus thunbergii subcommunity, Table 1: Ⅱ-A; VT5~VT7)과 천선과나무아군락(Ficus erecta subcommunity, Table 1: Ⅱ-B; VT8)의 2개 상위 단위로 구분되었으며, 전자는 다시 종군 8과 그 대립종군인 종군 11의 구성종 차이에 의해 굴피나무-소사나무변군락(Carpinus coreana-Platycarya strobilacea variant, Table 1: Ⅱ-A-1; VT5~VT6)과 곰솔과 구실잣밤나무가 상층 우점종으로 나타나는 구실잣밤나무-사스레피나무변군락(Eurya japonica-Castanopsis sieboldii variant, Table 1: Ⅱ-A-2; VT7)의 2개 하위 단위로 구분되었고, 전자는 다시 종군 9와 그 대립종군인 종군 10의 구성종 차이에 의해 굴참나무가 상층 우점종으로 나타나는 굴참나무-쌀새아변군락(Melica onoei-Quercus variabilis subvariant, Table 1: Ⅱ-A-1-a; VT5)과 곰솔이 상층 우점종으로 나타나는 애기등아변군락(Milletia japonica subvariant, Table 1: Ⅱ-A-1-b; VT6)의 2개 최하위 단위로 최종 구분되었다. 한편, 욕지도 산림식생단위를 한반도 산림식물군락체계(Černý et al., 2015)와 비교해 보면, 전반적으로 동백나무군강(Class Camellietea japonicae Miyawaki et Ohba 1963), 구실잣밤나무-자금우군목(Order Ardisio japonica-Castanopsietalia sieboldii Miyawaki et al. ex Hattori et Nakanishi 1983), 구실잣밤나무군단(Alliance Castanopsion sieboldii Suzuki 1952)의 식별종인 참식나무, 큰천남성, 마삭줄, 예덕나무, 동백나무, 보리밥나무 등이 우점종으로 나타나 이에 귀속되는 것으로 판단되었다.
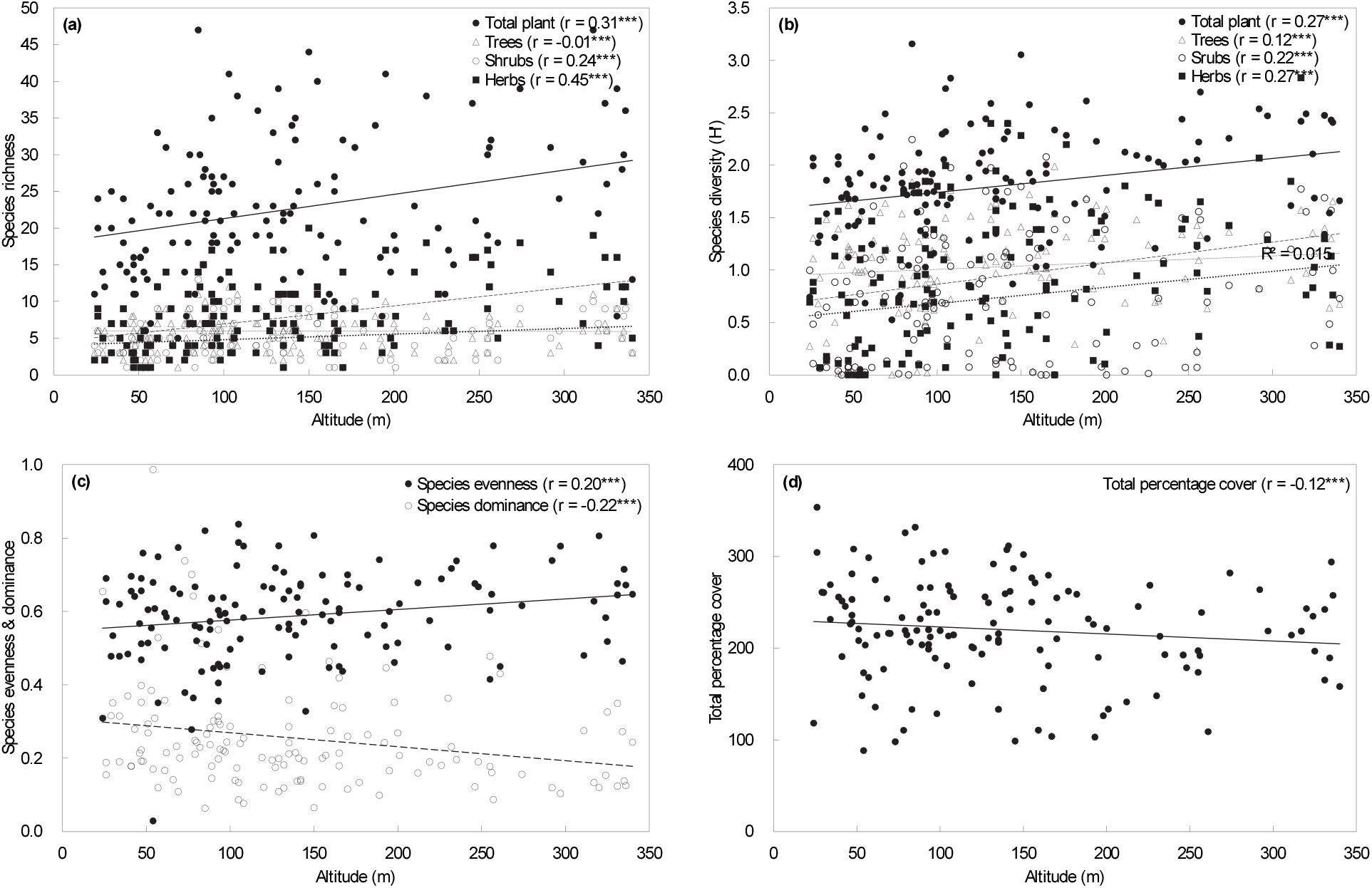
식생유형별 입지환경 및 종다양성 요소에 대한 분산분석을 수행한 결과(Table 2), 종풍부도는 전체 평균 22.6종이었고, 비목나무-줄딸기변군락(VT4)이 29.2종으로 가장 높았고, 왕대변군락(VT3)이 6.5종으로 가장 낮게 나타났는데 대개 초본식물의 종풍부성이 영향을 미친 것으로 보였다. 종균재도는 전체 평균 0.588이었고, 팽나무-사위질빵아군락의 전형변군락(VT2)이 0.646으로 가장 높은 값을 나타내어 다른 유형에 비해 구성종의 수도(abundance)가 다소 균질한 집단임을 알 수 있었다. 종다양도는 전체 평균 1.804이었고, 종풍부도와 마찬가지로 비목나무-줄딸기변군락(VT4)이 2.094로 가장 높은 값을 나타내어 8개 식생유형 중 군락 안정성이 가장 높음을 알 수 있었다. 종우점도는 전체 평균 0.254로 나타나 2~3종이 우점하는 왕대변군락(VT3), 구실잣밤나무-사스레피나무아변군락(VT7)과 천선과나무아군락(VT8)을 제외하면 전반적으로 다양한 종이 우점하고 있는 것으로 나타났다. 한편, 식생유형별 입지환경 인자 중 해발고도 및 낙엽층 깊이와 종다양성 요소 간 통계적으로 유의한 차이를 보이고 있었다(Table 2).
Note: a,b,c,d Duncan grouping code; P* <0.05; P** <0.01.
일반적으로 해발고도가 증가함에 따라 종다양성 역시 감소하는 단순 일차 선형 형태와 해발고도가 증가함에 따라 종다양성이 증가하여 중간 고도에서 가장 높아진 후 다시 감소하는 이차 포물선 형태의 두 가지 유형으로 구분되고 있다(Wang et al., 2007). McCain(2004)은 낮은 해발고도에서는 해발고도에 따른 종다양성과 종풍부도 등이 오히려 증가하다 특정 해발고도 이상이 되어서야 종다양성과 종풍부도가 감소한다고 하였는데 이러한 패턴의 이유는 중간 영역 효과(mid-domain effect: MDE) 때문인 것으로 보고하였다. 본 연구 결과, 욕지도 산림식생의 경우, 최대 해발고도가 394 m로 비교적 낮아 전반적으로 종풍부도, 종다양도, 그리고 종균재도는 해발고도가 높아짐에 따라 다소 증가하는 경향을 나타내었으며, 이러한 경향성은 통계적으로 유의하였고, 생육형 간에는 초본성 식물종이 가장 높은 유의성을 나타내고 있었다. 한편, 종우점도와 총피도는 해발고도가 높아짐에 따라 다소 감소하는 경향을 나타내고 있었다(Figure 2). 이를 종합하여 보면 욕지도 산림식생의 종풍부도와 종다양도는 해발고도가 높아짐에 따라 다소 증가하는 McCain의 중간영역효과에 따른 것으로 판단되었다.
한편, 과(family)별 속풍부도(genus richness) 및 종풍부도(species richness) 그리고 구성종의 총피도에 의한 상대적 우점순위를 가늠하는 합성지수인 상대기여도(r-NCD)를 분석한 결과, 전체적으로 속풍부도와 종풍부도에 있어서는 대개 식생천이 초기 단계에 출현하는 벼과식물(Poaceae), 콩과식물(Fabaceae), 국화과식물(Asteraceae) 등의 선구식물이 상위 그룹으로 나타났다. 상대기여도는 참식나무가 중심을 이루는 녹나무과(Lauraceae)가 100으로 가장 높았고, 이어 소나무과(Pinaceae) 96, 장미과(Rosaceae) 51, 두릅나무과(Araliaceae) 50, 자작나무과(Betulaceae) 36 등의 순이었으며, 전체 과의 85%인 68과가 10이하로 나타났다(Table 3).
Note : G, genus richness; S, species richness; r, r-NCD.
욕지도 산림식생의 중심을 이루는 교목성 구성종의 상대적 우세정도를 파악하기 위해 식생층위별 중요치(IV)와 전층 기준의 평균상대중요치(MIV)를 전체 산림식생과 현존 및 잠재적 우점성이 높다고 판단되는 곰솔림, 사방오리림, 구실잣밤나무림, 굴참나무림, 그리고 참식나무림 등 5개 주요 상관식생을 대상으로 산출한 후, 상위 10종을 기준으로 비교하였다(Figure 3). 먼저 전체 산림식생 구성종
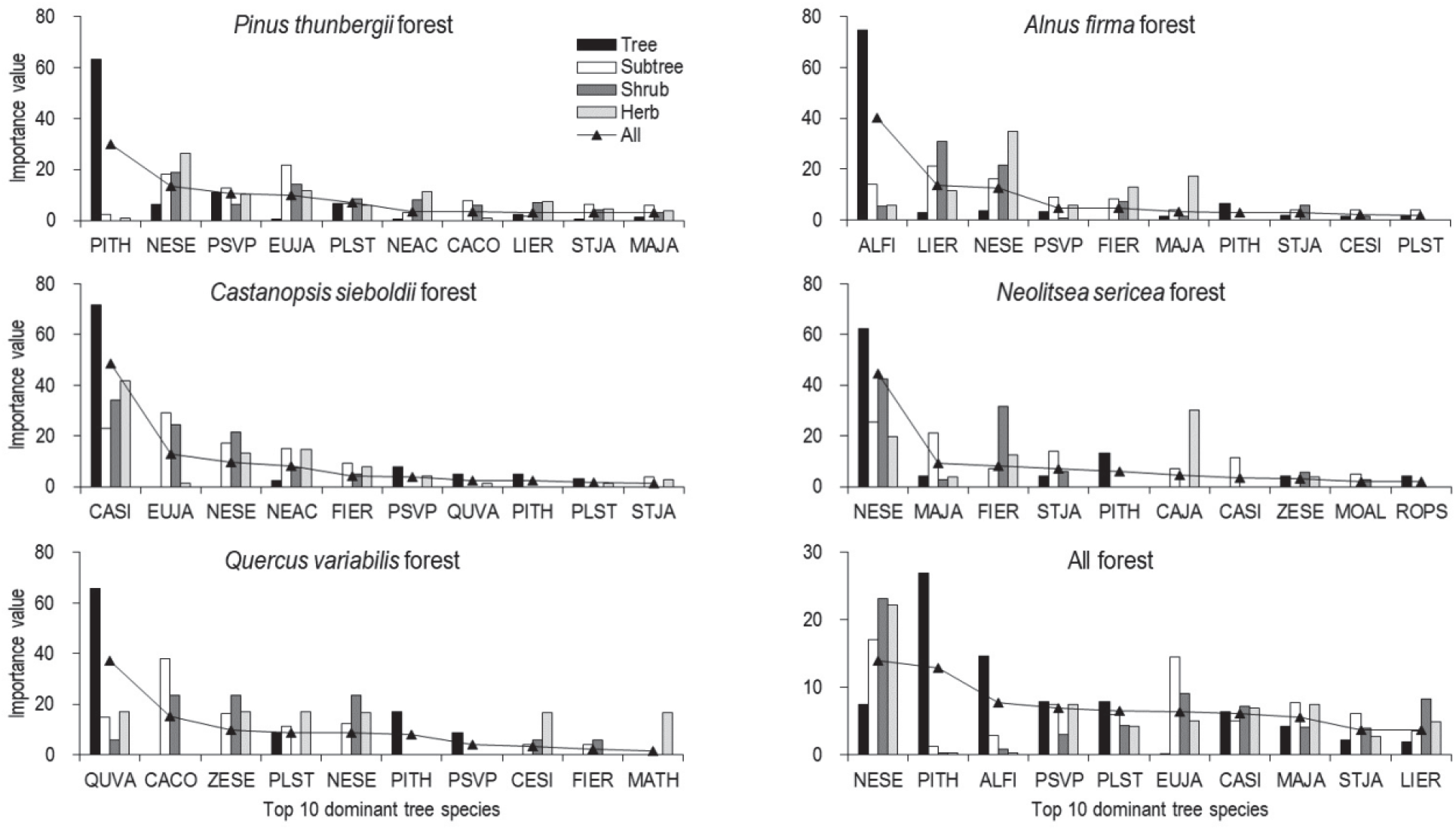
의 MIV에서는 상록활엽수인 참식나무가 13.9%로 가장 높게 나타났고, 다음으로 곰솔(12.8%), 사방오리(7.8%), 잔털벚나무(6.9%), 굴피나무(6.5%), 사스레피나무(6.3%), 구실잣밤나무(6.1%), 예덕나무(5.5%), 때죽나무(3.7%), 그리고 비목나무(3.6%) 등의 순으로 높게 나타났다. 식생층위 간에는 욕지도 산림식생의 광역 우점종인 곰솔과 사방오리의 경우, 교목층에서는 두 수종 모두 IV가 단연 높게 나타났지만, 아교목층 이하에서는 아주 낮게 나타났으며, 반면 상록활엽수인 참식나무와 구실잣밤나무는 식생층위에 관계없이 모든 층위에서 다른 교목성 수종 보다 상대적으로 높은 IV를 나타내고 있었다. 이로 미루어 보아 욕지도 산림식생의 천이경향은 더 이상의 외부교란 영향이 없는 한 이들 상록활엽수가 잠재자연식생으로 자리할 것으로 판단되었다.
한편, 주요 상관식생별 구성종의 MIV와 식생층위 간 IV 구성경향을 보면, 먼저 곰솔림(48개 조사구)에서는 총 48종(교목층 15종, 아교목층 23종, 관목층 34종, 초본층 25종)의 교목성 식물이 출현하였으며, MIV는 상관 우점종인 곰솔(30.0%)이 가장 높았고, 다음으로 참식나무(13.4%), 잔털벚나무(10.7%), 사스레피나무(10.0%), 그리고 굴피나무(6.8%) 등의 순으로 높게 나타났다. 이 임분은 상관우점종인 곰솔이 교목층에서 IV가 30.0%로 가장 높게 나타났지만, 아교목층 이하에서는 아주 낮은 IV를 나타내었고, 관목층과 초본층에서는 모두 이 지역의 대표적인 잠재자연식생 요소이자 상록활엽수인 참식나무의 IV가 가장 높게 나타났다. 이에 곰솔림은 후계수 이입 단절과 아교목층 이하에서 상대적으로 높은 IV를 나타내고 있는 참식나무, 잔털벚나무, 사스레피나무, 굴피나무 등의 영향으로 점차 쇠퇴할 것으로 판단되었다. 사방오리림(24개 조사구)은 총 29종(교목층 11종, 아교목층 16종, 관목층 25종, 초본층 7종)의 교목성 식물이 출현하였으며, MIV는 상관우점종인 사방오리(40.1%)가 가장 높았고, 다음으로 비목나무(13.5%), 참식나무(12.7%), 잔털벚나무(4.8%) 등의 순으로 높게 나타났다. 이 임분은 상관우점종인 사방오리가 교목층에서 IV가 74.7%로 단연 높게 나타났지만, 아교목층 이하에서는 비교적 낮은 IV를 나타내었고, 반면 온난화 지표종인 비목나무와 참식나무의 IV가 15~30내외로 높게 나타나 점차 쇠퇴할 것으로 판단되었다. 구실잣밤나무림(12개 조사구)은 총 19종의 교목성 식물(교목층 8종, 아교목층 7종, 관목층 8종, 초본층 15종)
이 출현하였으며, MIV는 상관 우점종인 구실잣밤나무(48.6%)가 단연 높았고, 다음으로 사스레피나무(12.9%), 참식나무(9.6%), 새덕이(8.0%) 등의 순으로 모두 상록활엽수가 상위 IV 그룹을 이루고 있었다. 이 임분은 상관우점종인 구실잣밤나무가 모든 식생층위에서 상위 IV를 나타내고 있어 잠재적으로 더욱 안정된 임분으로의 발달이 예상되었다. 굴참나무림(3개 조사구)은 총 12종의 교목성 식물(교목층 4종, 아교목층 7종, 관목층 8종, 그리고 초본층 6종)이 출현하였으며, MIV는 상관우점종인 굴참나무(37.2%)가 가장 높았고, 다음으로 소사나무(15.2%), 느티나무(9.8%), 굴피나무(8.7%), 그리고 참식나무(8.6%) 등의 순으로 높게 나타났다. 이 임분은 상관우점종인 굴참나무가 교목층에 IV가 65.8%로 단연 높았으나 아교목층 이하에서는 소사나무, 느티나무, 참식나무 등이 상대적으로 높은 IV를 나타내고 있어 향후 이들과의 종간경쟁이 점차 심화될 것으로 판단되었다. 참식나무림(6개 조사구)은 총 19종의 교목성 식물(교목층 8종, 아교목층 9종, 관목층 8종, 그리고 초본층 9종)이 출현하였으며, MIV는 상관우점종인 참식나무의 IV가 44.7%로 단연 높았고, 다음으로 예덕나무(9.1%), 천선과나무(8.0%), 때죽나무(7.1%) 등 소교목성 식물 등의 순으로 높게 나타났다. 이 임분은 상관우점종인 참식나무가 모든 식생층위에서 상위 IV를 나타내고 있고 교목성 경쟁 수종의 이입 및 정착이 거의 없어 더 이상의 외부교란 영향이 없는 한 생태적으로 보다 안정된 군락으로의 발달이 예상되었다.
욕지도 산림식생의 생태적 관리의 기반을 조성하기 위하여 상층 우점종에 의한 상관식생유형(physiognomic vegetation types)을 기준으로 현존식생도(1/5,000)를 작성하고(Figure 4), 상관식생유형별 분포 면적과 패치 수를 산출하여 그 공간분포 특성을 파악하였다(Table 4). 상관식생도의 최소 작도 면적은 0.25 ha 이상을 기준으로 하였지만 특이하거나 보전 가치가 높은 식생유형에 대해서는 0.01 ha 이상도 포함하였다.
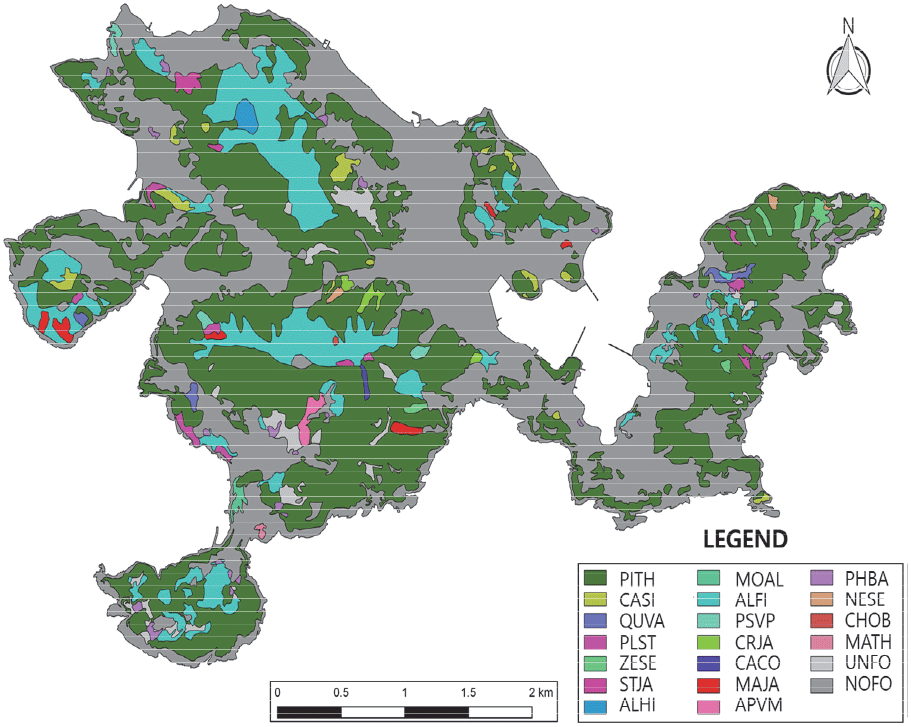
Note: NV, natural vegetation; AV, artificial vegetation.
산림식생의 공간분포 면적은 욕지도 총면적(1368.35 ha)의 57.8%인 790.7 ha로 나타났으며, 임상별 구성비는 침엽수림이 72.7%로 단연 높았고, 이어 낙엽활엽수림(22.0%), 상록활엽수림(1.6%), 죽림(0.7%) 등의 순이었다. 상관식생유형(무입목지 제외)은 곰솔림, 사방오리림, 구실잣밤나무림(모밀잣밤나무림 포함), 참식나무림, 예덕나무림 등을 포함하여 총 18개 유형으로 나타났다. 상관식생유형 간 분포면적은 곰솔림이 전체 산림식생 면적의 72.3%인 571.5 ha로 가장 크게 나타났고, 다음으로 사방오리림 17.4%(137.3 ha), 구실잣밤나무림 1.4%(10.9 ha) 등의 순으로 높게 나타났으며 무입목지는 2.9%(23.2 ha)이었다. 한편, 산림식생의 단편화 정도를 가늠하는 패치(patch) 개수는 총 230개로 나타났으며, 이 중 곰솔림(58개)이 가장 많았고, 이어 사방오리림(41개), 왕대림(25개), 구실잣밤나무림(16개) 등의 순이었다. 산림식생의 패치당 평균 면적은 3.4 ha이었고, 상관식생유형 간에는 곰솔림(9.9 ha)이 가장 크게 나타났고, 이어 사방오리림(3.4 ha), 고로쇠나무림(2.9 ha) 등의 순이었으며, 구실잣밤나무림, 굴피나무림, 느티나무림, 왕대림, 예덕나무림, 참식나무림, 소사나무림, 후박나무림, 편백림 등 9개 유형은 모두 1 ha이하이었다.
결 론
일반적으로 도서지역 산림생태계는 지리적 격리성과 해양 환경의 영향으로 생태적 특이성과 생물적 고유성이 높지만 외부 교란 영향에 대한 생태적 저항력 및 회복력이 낮은 취약성도 갖고 있어 세계 각국에서 산림식생을 중심으로 그 생태적 보전과 관리를 위한 다양한 연구가 진행되어 오고 있다(Ren et al., 2012; Hanson and McElroy, 2015; Balzan et al., 2018; Silva et al., 2019). 본 연구는 우리나라 남해안에서 세 번째로 큰 유인도서인 욕지도의 산림생태계의 기반을 이루는 산림식생의 생태적 보전 및 관리를 위한 기초자료를 제공하기 위하여 식물사회학적 방법으로 식생유형을 구분함과 아울러 그 생태적 특성을 비교하고 상관식생에 의한 현존식생도를 작성하여 산림식생의 공간분포 특성을 파악하였다. 식생유형은 총 8개 유형이 구분되었고 2개 군락(community), 4개 아군락(subcommunity), 6개 변군락(variant), 그리고 2개 아변군락(subvariant)의 식생단위 체계를 갖고 있었는데, 전반적으로 한반도 산림식물군락체계(Černý et al., 2015)상 동백나무군강(Class Camellietea japonicae Miyawaki et Ohba 1963), 구실잣밤나무-자금우군목(Order Ardisio-Castanopsietalia sieboldii Miyawaki et al. ex Hattori et Nakanishi 1983), 구실잣밤나무군단(Alliance Castanopsion sieboldii Suzuki 1952)의 식별종인 참식나무, 마삭줄, 예덕나무, 동백나무, 보리밥나무, 큰천남성 등이 우점으로 나타나 이에 귀속되는 것으로 판단되었다. 한편 식생유형별 입지환경인자와 종다양성 지수에 대한 분산분석을 수행한 결과, 해발고도와 낙엽층 깊이에서 통계적으로 유의한 차이를 보이고 있음을 알 수 있었다. 해발고도와 종다양성 지수와의 상관관계를 분석한 결과, 전반적으로 종풍부도, 종다양도, 그리고 종균재도는 해발고도가 높아짐에 따라 다소 증가하는 경향을 나타내었는데 주로 초본성 식물종이 영향을 큰 것으로 판단되었다. 이러한 경향성은 통계적으로 유의한 결과를 보였다. 또한, 종우점도와 총피도는 해발고도가 높아짐에 따라 다소 감소하는 경향을 보였다. 해발고도가 높아짐에 따라 종풍부도와 종다양도가 증가하는 이러한 결과는 비교적 낮은 해발고도를 가진 산지에서는 중간영역효과(MDE)로 종다양성이 증가한다는 McCain의 연구 결과를 반영하고 있었다. 욕지도 산림식생의 교목성 구성종의 상대적 우세정도를 파악하기 위한 평균상대중요치(MIV)는 상록활엽수인 참식나무(13.9)가 가장 높았고, 다음으로 곰솔(12.8), 사방오리(7.8) 등의 순이었으며, 식생층위 간에는 잠재적 천이경향을 가늠할 수 있는 아교목층 이하에서 대부분 참식나무, 구실잣밤나무, 사스레피나무 등 상록활엽수 요소와 온난화 지표종인 비목나무가 상대적으로 높은 중요치를 나타내고 있었다. 과별 구성종의 총피도에 의한 상대적 우점순위를 가늠하는 합성지수인 상대기여도(r-NCD) 산출 결과, 참식나무와 비목나무가 중심을 이루는 녹나무과가 100으로 가장 높게 나타났고, 다음으로 소나무과 96.0, 장미과 50.8, 두릅나무과 49.5, 자작나무과 35.9 등의 순이었으며, 욕지도 산림식생은 외부간섭이 없는 한 혼효림 도중 단계를 거쳐 잠재적으로 상록활엽수가 우점하는 숲으로 진행천이(progressive succession)가 이루어질 것으로 판단되었다. 끝으로 본 연구는 욕지도 산림식생의 생태적 보전 및 관리를 위한 기초정보 제공을 위하여 식생유형을 구분하고 그 생태적 및 공간분포 특성을 비교하였지만 향후 욕지도 산림식생의 특이성과 고유성 그리고 주요 개체군의 크기 감소 및 유성번식력 저하 등에 따른 생물다양성 감소 원인을 규명하는 후속 연구가 뒤따라야 보다 실질적인 생태적 보전 및 관리 방안이 모색될 것으로 판단되었다.