Article
생육공간을 공유하는 소나무와 졸참나무의 수분 이용 전략 비교 분석
이기웅
1, 이보라
2, 조낭현
3, 임종환
1, 김은숙
1,*
Analysis of Water Use Strategies of Two Co-occurring Mature Tree Species, Pinus densiflora and Quercus serrata
Kiwoong Lee
1, Bora Lee
2, NangHyun Cho
3, Jong-Hwan Lim
1, Eun-Sook Kim
1,*
1Forest Ecology Division, National Institute of Forest Science, Seoul 02455, Korea
2Warm Temperature and Subtropical Forest Research Center, National Institute of Forest Science, Jeju 63582, Korea
3Department of Environmental Science, Kangwon National University, Chuncheon 24341, Koera
© Copyright 2022 Korean Society of Forest Science. This is an Open-Access article distributed under the terms of the
Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits
unrestricted non-commercial use, distribution, and reproduction in any
medium, provided the original work is properly cited.
Received: Apr 11, 2022; Revised: Jul 19, 2022; Accepted: Aug 01, 2022
Published Online: Sep 30, 2022
요 약
본 연구는 한국 산림생태계의 주요 우점종인 소나무와 졸참나무의 수분 이용 전략을 비교 분석하기 위해, 2019년 3월부터 12월까지 경기도 포천시에서 시행되었다. 소나무(n=6)와 졸참나무(n=3)의 수액속밀도와 환경 변수인 대기 온도, 상대습도, 강수량, 순복사량, 토양수분을 측정하여 계절변화 및 환경 변수에 따른 수액 이동 특성을 분석하였다. 그 결과, 수종의 최대 수액속밀도는 생장 기간(5-10월)에 졸참나무가 소나무보다 2배 가까이 높았으며, 졸참나무는 9월에 소나무는 8월에 가장 높았다. 수액속밀도에 영향을 주는 주요한 환경 변수들은 포화수증기압차와 순복사량이었으며, 대기 온도는 영향을 주지 못했다. 이력현상 분석에서 소나무는 건조해짐에 따라 기공을 닫고 졸참나무는 여전히 기공을 열어두는 전략을 사용하는 것을 발견하였다. 수관전도도 역시 최대 수액속밀도와 비슷한 경향을 보이며, 졸참나무의 수관전도도가 소나무의 수관전도도보다 약 2배 정도 더 높았다. 본 연구에서는 두 수종의 수분 이용 전략을 비교 분석하였고, 더 명확한 기작의 이해를 위해 생리적 특성뿐만 아니라 형태적 특성에 관한 연구도 병행되어야 할 것이다.
Abstract
The study was carried out in Pocheon-si, Gyeonggi-do from March to December in 2019 to compare and analyze the water use strategies of two co-occurring tree species, Pinus densiflora and Quercus serrata, both native and dominant in Korea's forest ecosystems. Through seasonal changes, we measured environmental variables such as air temperature, relative humidity, precipitation, net radiation, and soil water content. Sap flow densities of P. densiflora (n = 6) and Q. serrata (n = 3) were measured, along with environmental variables. The maximum sa pflow density for Q. serrata almost doubled that of P. densiflora during the growing season, while the maximum sap flow densities in both Q. serrata and P. densiflora peaked in September and August, respectively. Net radiation and vapor pressure deficit, but not air temperature, were the major environmental variables significantly affecting sap flow density. Analysis of hysteresis revealed that P. densiflora exhibited isohydric behavior, while Q. serrata showed anisohydric behavior. Analysis of crown conductance revealed similar trends as sap flow density, i.e., the crown conductance of Q. serrata was twice that of P. densiflora during the growing period. The study compared and analyzed the water use strategies between two co-occurring species. To better understand the underlying mechanisms of water use, more research on both physiological and morphological traits are needed.
Keywords: co-occurring species; sapflow density; water use strategies; crown conductance; hysteresis
서 론
우리나라 산림생태계는 소나무류와 참나무류가 우점하고 있는 임분으로 상당 부분 구성되어 있다. 2015년 1:5,000 수치 임상도 기준으로 소나무류는 전체 산림 면적의 약 26.2%, 참나무류는 약 25.4%를 차지하고 있으며, 침활혼효림의 면적 비율은 약 11.9%이다. 침활혼효림 중, 소나무류와 참나무류의 혼합림이 많은 것을 고려했을 때, 전국 산림의 약 3분의 2가량이 소나무류와 참나무류 수종으로 이루어져 있다고 말할 수 있다. 소나무류와 참나무류는 한국 산림에서 넓은 면적을 차지하고 있을 뿐만 아니라 경제적, 생태적으로도 중요한 부분을 차지하고 있어 우리 문화와도 밀접하게 연관되어 있다(Lee et al., 2006). 이러한 가치들과 맞물려 소나무류와 참나무류에 관한 연구들은 생장(Lee et al., 2000; Seo et al., 2001; Lee et al., 2004), 형태(Lee et al., 2003), 분포(Lee et al., 2006) 등 다양한 분야에서 많은 연구가 진행되었다.
최근에는 기후변화 시나리오에 따른 소나무류와 참나무류의 식생 분포 및 적지 변화에 관한 연구들도 진행되었다. Ko et al.(2014)은 기후변화 시나리오에 따른 소나무의 미래 분포도를 예측하였는데, 그 분포 면적이 크게 줄어든다고 보고하였다. 생태적 지위 모델을 이용한 분석에서는(Chun and Lee, 2013) 주로 남부 지방에 몰려 있던 소나무가 동쪽과 북쪽으로 이동하는 것으로 나타났다. Lee et al.(2014)은 기후변화 시나리오 RCP 4.5와 RCP 8.5를 적용하여 신갈나무의 적지 분포 변화를 추정하였다. RCP 4.5를 적용했을 때, 신갈나무는 2030년에서 2090년까지 74만ha에서 17만ha까지 감소하는 것으로 나타났다. 그리고 분포지 역시 2090년에는 강원도 일부 지역에만 국한된다고 예측하였다. 상수리나무의 분포 역시 일부 지역으로 제한될 것으로 예상된다(Korea Forest Research Institute, 2014).
기후변화 시나리오에 따른 수목의 적지 분포 감소뿐만 아니라 수분스트레스의 피해가 예상되는 상황에서, 이에 따른 식물의 수분 이용 전략에 대한 연구(Martin-Benito et al., 2017; Fauset et al., 2018)이 최근에 많이 수행되고 있다. 또 다양한 수종을 대상으로 수분과 관련된 생리적 기작과 서로 공존하는 식물의 수분 이용 전략에 관한 관심이 높아지고 있다(Sànchez-Costa et al., 2015). 식물의 수분 이용 전략의 대표적인 예로는 수분스트레스를 받을 때 기공을 조절하여 수분의 손실을 막는 방법(isohydric)과 기공을 조절하지 않고 그대로 열어 두는 방법(anisohydric)이 있다(Klein, 2014).
우리나라에서도 기후변화에 민감하게 반응하는 이 두 수종의 생리 기작을 이해하기 위해 많은 연구가 시행되었다. 소나무 유묘를 이용한 온도 증가와 관수 조절 실험에서 광합성, 엽록소와 생물량의 변화(Park et al., 2016), 기공전도도, 수분이용효율 및 증산 속도의 변화(Chang et al., 2019), 온도와 CO2 농도 증가에 따른 바이오매스와 광색소 함량의 변화(Han and Shin, 2020)에 관해 연구가 진행되었다. 참나무류 중 하나인 굴참나무 묘목의 온난화 처리 실험에 따른 엽록소 및 광합성 변화(Lee et al., 2013), 그리고 Song et al.(2020)은 온도와 CO2 농도 처리에서 두 수종의 광합성과 바이오매스의 변화를 비교 실험하였다.
하지만 이런 실험들은 모두 유묘를 대상으로 시행되었으며, 생육공간을 공유하여 경쟁 관계(Lee et al., 2006)에 있는 두 수종을 대상으로 한 현장 실험은 국내에선 찾아보기 힘든 실정이다. 특히 기후변화로 점점 심각해지는 가뭄과 토양 건조에 대한 두 수종의 수분 이용 전략에 관한 연구는 미미하다. 이미 전 세계적으로 기후변화에 대응하여 효과적인 산림관리를 위해 수목들의 수분 이용 전략에 관한 연구들이 진행되고 있으며, 이런 연구는 국내에서도 필요하다고 생각된다. 본 연구의 가설은 생육공간을 공유하는 두 수종은 다른 수분 이용 전략을 사용한다는 것이다. 이 가설을 검증하고자, 소나무와 졸참나무의 수분 이용 전략에 관해 연구하였다.
재료 및 방법
1. 연구대상지 및 대상 수종
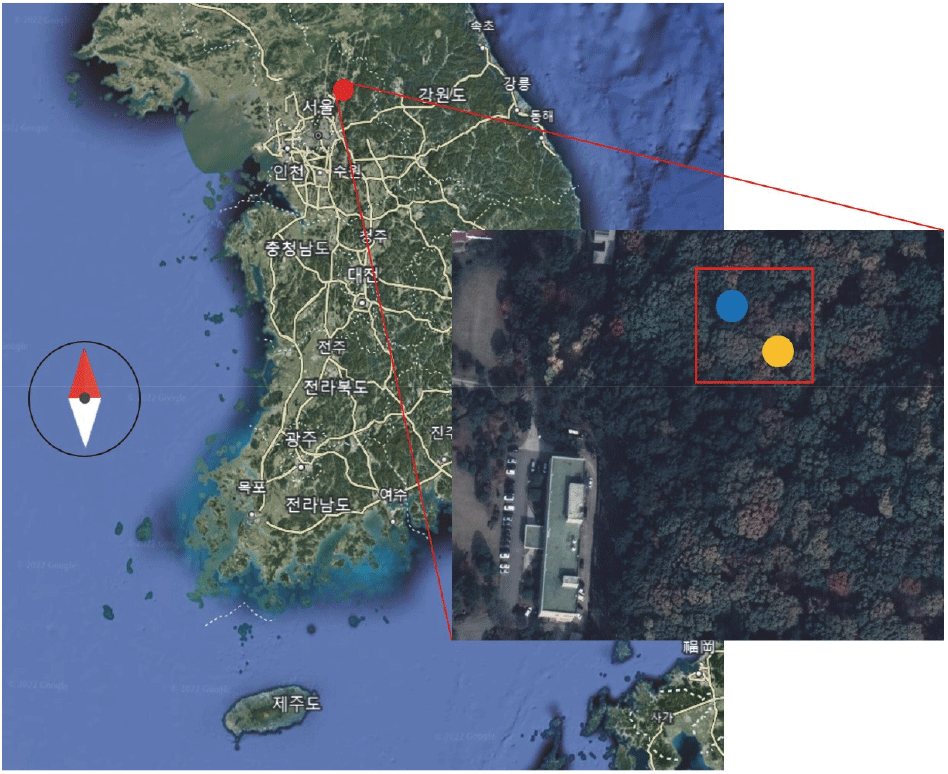
본 연구는 2019년 3월에서 12월까지 경기도 포천에 있는 국립산림과학원 산림기술경영연구소 내 시험림에서 시행되었다(Figure 1). 포천의 20년간의 평균 기온은 11.4°C이며 평균 연 강수량은 1402.2 mm이다. 시험림 내에 약 0.5 ha의 조사구를 설치하였다(37°75’N, 127°17’E). 조사구는 해발고도 약 150 m에 위치하고, 전반적으로 서사면이며, 경사는 약 10~15〬로 비교적 완만하였다. 토성은 사양토이며 pH는 4.51(±0.03), 유기물 함량은 8.9%(±0.32), 그리고 용적밀도는 1.04 g/cm3(±0.03)다. 조사구 내 소나무, 잣나무, 졸참나무가 상층을 우점하는 임분이 일정 영역을 차지하며 인접해 있다. 관목층에는 당단풍, 노린재나무, 다릅나무, 까치박달 등이 생육하고 있다. 소나무와 졸참나무 임분에 대해 지형과 입지가 동일하다고 판단되는 지역에 수종별 조사구를 1개씩 설치하였다(Figure 1). 소나무 임분과 졸참나무 임분 내 대경목인, 소나무 6본과 졸참나무 3본을 실험 대상으로 선정하였다. 소나무의 평균 흉고직경(DBH)은 37.8 cm(31.2~51.2 cm)이고 졸참나무의 평균 DBH는 46.2 cm(41.0~51.6 cm)였다. 소나무의 평균 흉고단면적(BA)은 1160.1 cm2며, 졸참나무의 평균 BA는 1687.8 cm2였다(Table 1).
Figure 1.
Location of study site, Pocheon. The Red square indicates the study site. The blue and yellow circles in the red square represent Pinus densiflora subplot and Quercus serrata subplot, respectively.
Download Original Figure
Table 1.
DBH and BA of target species(PD and QS represent P. densiflora and Q. serrata, respectively).
|
PD |
DBH(cm) |
BA(cm2) |
QS |
DBH(cm) |
BA(cm2) |
|
1 |
36.8 |
1063.1 |
1 |
45.9 |
1653.8 |
|
2 |
32.9 |
849.7 |
2 |
41.0 |
1319.6 |
|
3 |
31.2 |
764.2 |
3 |
51.6 |
2090.1 |
|
4 |
34.8 |
950.7 |
|
|
|
|
5 |
40.3 |
1274.9 |
|
|
|
|
6 |
51.2 |
2057.8 |
|
|
|
|
Mean |
37.8 |
1160.1 |
Mean |
46.2 |
1687.8 |
Download Excel Table
2. 연구 방법
1) 환경인자 측정
대기 온도(TA: °C), 상대습도(RH: %), 순복사량(Rn: W m−2), 강수량(Rainfall: mm)이 환경 변수로 이용되었고, 포화수증기압차(VPD: KPa)는 RH와 TA를 이용하여 계산하였다. 외부 기상 환경인자는 국립수목원 광릉 활엽수림 플럭스타워에서 측정된 자료를 사용하였고, 토양수분(SWC: m3 m−3)의 경우, 수종별 조사구마다 토양 깊이 0~20 cm사이에 설치된 2~4개의 토양수분 센서(S-SMC-M005, Onset Computer Corp., Pocasset, MA)를 이용하여 측정한 자료를 사용하여 평균값을 산출하였다.
2) 수액속밀도(Sapflow density) 측정
수액속밀도를 측정하기 위해 열손실탐침법인 Granier 타입 센서를 설치하였다(Granier, 1987). Granier 타입 센서는 길이 20 mm인 두 개의 탐침으로 구성되어 있으며, 하나는 열침이고 다른 하나는 기준침이다. 이 두 개의 탐침은 줄기와 수직이 되게 설치하며, 탐침간 10~15 cm의 거리를 두고 열침은 상단부, 기준침은 하단부에 줄기와 평행하게 설치한다. 열침에는 열선이 감겨있어 열을 발생시킨다. 증산작용을 통해 수액류의 이동 속도가 빠를 경우, 열침에서 더 많은 열을 빼앗고, 수액류의 속도가 느릴 경우, 적은 열을 빼앗게 된다. 이때, 열침과 기준침에서 발생하는 온도차를 이용하여 수액류를 산출한다. 두 탐침은 수목의 북쪽에 설치되었고 또, 알루미늄 포일로 감싸진 플라스틱 통을 씌워서 직사광선의 간섭을 최소화하였다. 수액속밀도는 아래의 식으로 계산되었다. 데이터는 30초마다 측정되었으며 30분마다 평균값이 데이터 로거에 저장되었다.
여기서, Js는 수액속밀도, ΔT는 두 탐침 간의 온도 차, ΔTmax는 두 탐침 간의 최대 온도 차를 의미한다.
3) 이력현상(Hysteresis)과 환경인자
이력현상은 식물의 생리적 변수와 기상변수 사이에서, 각 변수가 최대치에 도달할 때 발생하는 시차를 의미한다(Wang et al., 2019). 이력현상은 증산 또는 수액속밀도와 환경인자의 관계에서 발생하며, 이미 다양한 생태계와 식물에서 빈번하게 발견되는 중요한 현상이다(Zeppel et al., 2004; Ewers et al., 2005; O’Grady et al., 2008). 이는 식물의 수분 이용, 수분 이용 효율 그리고 토양의 수분 고갈 상태를 반영할 수 있다(Zhang et al., 2014). 이력현상에 기인하는 중요한 특성은 오전에 증산 또는 수액속밀도의 상승이 환경 인자의 상승보다 빠르거나 늦어 발생하는 시차이다(Zhang et al., 2019). 예를 들어, 수액속밀도의 상승이 먼저 발생하고 VPD의 상승이 발생하는데 이때 생기는 시차로 인해 수액속밀도와 VPD의 이력현상은 시계 방향의 움직임을 보인다. 또한, 이 현상은 식물이 과도한 수분 손실이나 탈수를 방지하기 위해 하나의 방어 기작으로도 알려져 있다(Chen et al., 2011).
Js와 환경인자인 TA, VPD, 그리고 Rn과의 관계를 분석하였고, 이력현상을 분석하기 위해 토양수분이 가장 낮은 5일 동안(186~190일)의 Js와 VPD의 데이터를 선택하여 표준화하였다.
4) 수관전도도(Crown conductance: Gs)
실험 대상 수종의 수고가 높아서 기공전도도와 수분포텐셜을 측정할 수 없었다. 그러므로, 기공의 반응을 분석하기 위해 Gs를 아래의 식을 이용하여 구하였다(Ewers et al., 2005).
여기서 KG는 전도계수(115.8 ± 0.4236 (TA) kPa m3 kg−1)이며, 이는 습도 상수, 기화 잠열, 일정 압력에서의 공기 비열, 그리고 공기 밀도를 의미한다(Phillips and Oren 1998). TA는 대기 온도(°)이며, EL는 증산량을 의미한다. 하지만 이 연구에서 수관면적을 측정할 수 없었기 때문에 Js를 EL의 프록시로 사용하였다(Meinzer et al., 2013). GS의 단위는 Pearcy et al.(1989)의 식에 의해 m s−1에서 mmol m−2 s−1로 변환되었다. 수관전도도를 계산할 때 낮은 광도가 Gs에 미치는 영향을 최소화하기 위해 Rn이 400 W m−2 이상(10:00~16:00 시)일 때, Js와 VPD(>0.5 kPa)를 사용하였다(Ewer and Oren, 2000; Meinzer et al., 2013).
5) 통계 분석
소나무와 졸참나무의 수액속밀도와 수관전도도를 비교하기 위해 t-test를 사용하였고, 수액속밀도와 환경인자를 분석하기 위해 회귀분석을 사용하였다. 모든 분석은 통계 분석 프로그램 R(R Core Team, 2021)을 통하여 수행되었고, 모두 유의수준 α=0.05에서 검정하였다.
결과 및 고찰
1. 환경 변수
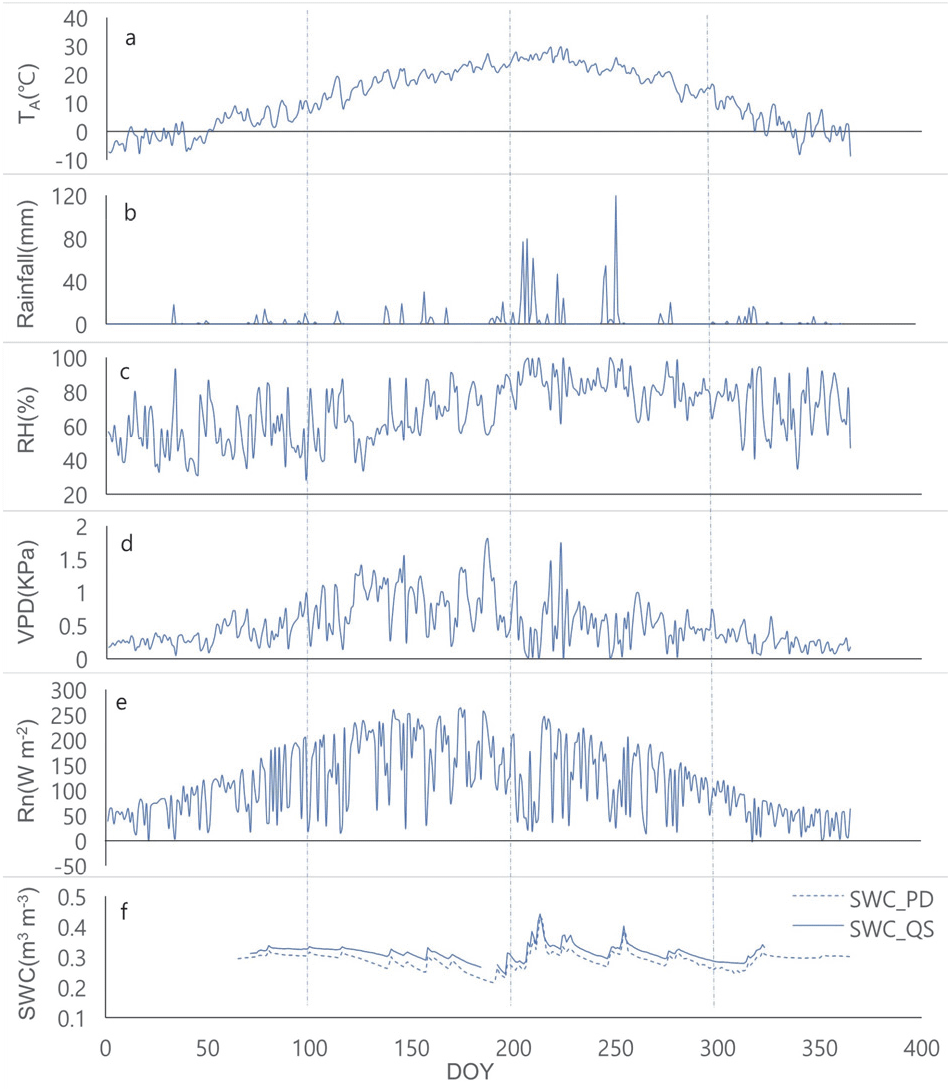
2019년 평균 온도는 11.8°C이고, 최대 일 평균 온도는 29.6°C, 최소 일 평균 온도는 −8.7°C였다. 총강수량은 961.2 mm였고, 7월의 강수량이 337.5 mm로 가장 많았다. 연평균 RH는 68.6%(28.6-99.6%), VPD는 0.52 KPa(0.01-1.81 KPa), Rn은 114.4 W m−2(−2.37-263.4 W m−2)였다. SWC는 소나무 조사구에서 0.29 m3 m−3(0.21-0.43 m3 m−3), 졸참나무 조사구에서 0.31 m3 m−3(0.24-0.44 m3 m−3)였다. SWC는 두 수종 모두 6월에 가장 낮았으며, 7월에 비가 온 후에 가장 높았다(Figure 2).
Figure 2.
Environmental variables in 2019. a, b, c, d, e, and f represent air temperature, rainfall, relative humidity, vapor pressure deficit, net radiation, and soil water contents, respectively. PD, QS, and DOY on Figure f mean Pinus denisflora, Quercus serrata, and day of year, respectively.
Download Original Figure
2. 수액속밀도
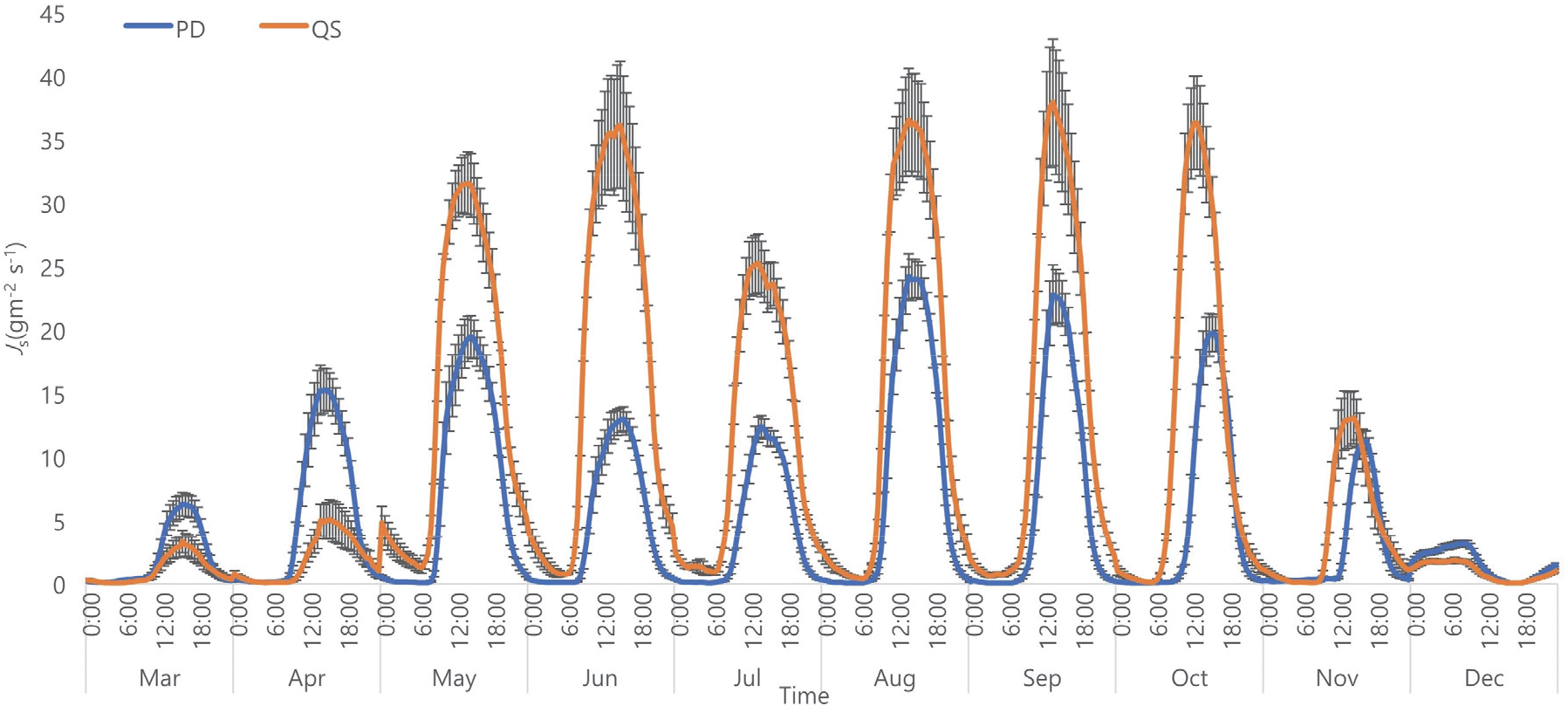
생장기간(5~10월) 졸참나무의 최고 수액속밀도(Js_max: 33.9 g m−2 s−1)는 소나무의 Js_max(18.6 g m−2 s−1)보다 약 2배 정도 높았고 유의한 차이를 보였다(p<0.05)(Figure 3). 소나무의 Js_max는 4월부터 급격히 증가하여 5월까지 증가하고 그 이후로 7월까지 점진적으로 감소하였다. 그리고 8월에 가장 높은 Js_max(24.2 g m−2 s−1)을 보이며 10월까지 점진적으로 감소하였다. 반면, 졸참나무의 Js_max는 개엽 시기인 5월에 급격히 증가하여 6월까지 증가하였다. 그러나 7월에 감소 한 후, 8월에 다시 증가하여 9월에 최고치(37.9 g m−2 s−1)에 도달하고 11월에 낙엽이 지는 시기와 맞물려 급격히 감소하였다. 소나무는 졸참나무보다 토양수분의 고갈에 대해 더 민감하게 반응하였다. 토양수분의 감소에 따라 소나무의 Js_max 또한 감소하였지만(5~6월, 8~10월)(Figure 3), 졸참나무는 토양수분이 감소하여도 상대적으로 비슷한 Js_max를 유지하였다. 두 수종의 수액속밀도의 차이는 환공재와 가도관의 구조적 차이에서 발생하는 것으로 생각되며, Zeppel et al.(2010)은 도관의 지름이 더 클수록 더 높은 수분 이동 효율을 가진다고 설명하였다. 그리고 두 수종의 토양수분에 대한 반응은 두 수종의 기공이 토양 건조에 대해 반응하는 차이라고 생각된다. 소나무는 건조할 때 기공을 닫아 잎 내 수압을 유지하는 isohydric 방법(Matyssek et al., 2014)을 이용하고, 반면에 졸참나무는 건조에도 기공을 열어두는 anisohydric 방법(Harayama et al., 2019)을 사용한다. 이 결과들은 선행연구의 결과와 유사하다. Menizer et al.(2013)는 환공재 수종인 참나무류의 Js가 산공재 수종과 침엽수종보다 더 높다는 것을 발견하였다. 그리고 이 수종들의 토양 건조와 VPD에 대한 반응이 다른 수종들(산공재와 침엽수종)보다 민감하지 않으며, 이에 따라 기공에 의한 잎의 수분포텐셜 조절도 덜 민감하다. 이러한 기능적 특성들이 환공재라는 구조적 특성과 맞물려 더 넓은 범위의 토양수분을 이용할 수 있고, 또 건조한 상황에서도 더 많은 토양수분을 이용할 수 있다고 설명하였다.
Figure 3.
Daily variations of averaged sapflow density in different months between two species. PD and QS indicate Pinus densiflora and Quercus serrata, respectively. The error bars present standard errors.
Download Original Figure
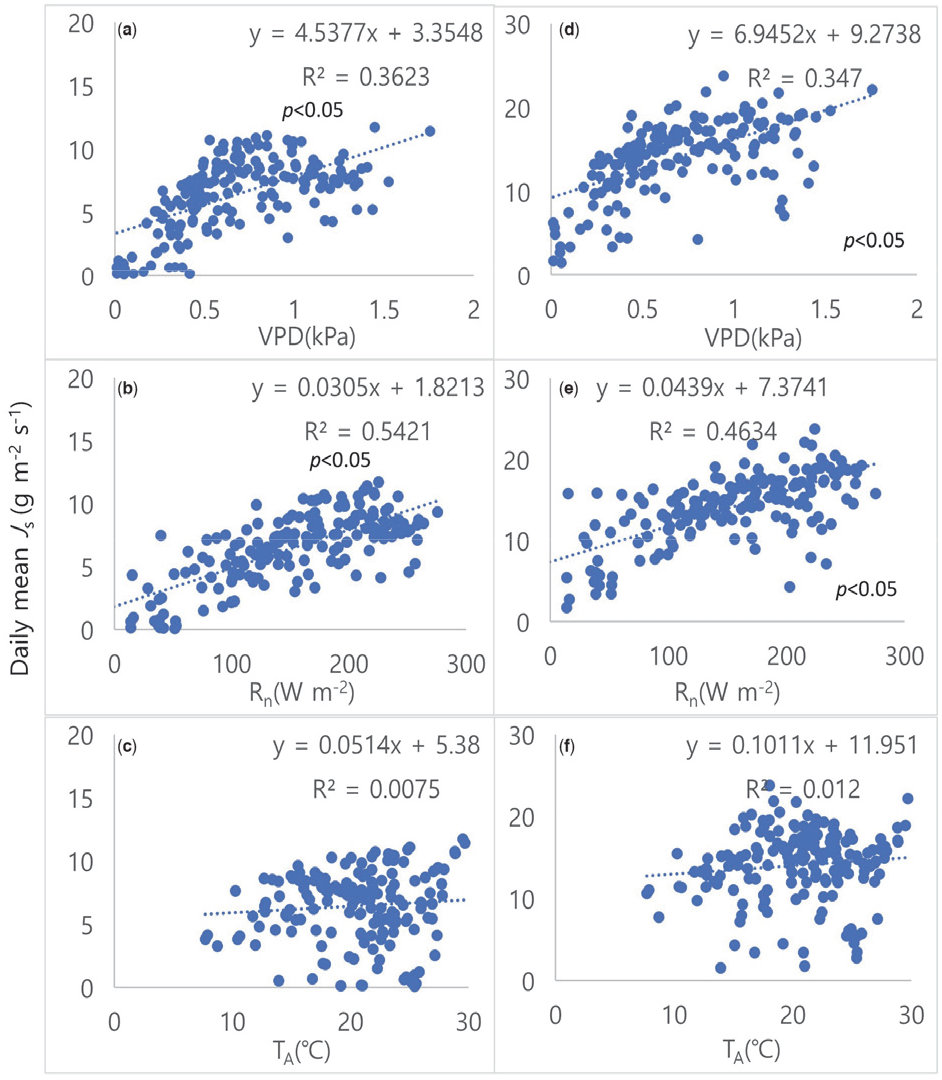
3. 수액속밀도와 환경 변수의 관계
일 평균 Js와 환경 변수와의 관계는 다음과 같다(Figure 4.). 두 수종 모두, VPD와 Rn이 Js를 조절하는 주요한 환경 변수였으며[Figure 4(a), 4(b), 4(d), and 4(e)], TA는 Js의 유의한 영향을 미치지 않았다[Figure 4(c) and 4(f)]. VPD와 Rn이 Js에 영향을 주는 주요한 요인이라는 것은 많은 선행연구의 결과와 유사하다(Sanchez-Costa et al., 2015; Liu et al., 2017; Zhu et al., 2017; Oogathoo et al., 2020). 반면에, TA 역시 Js에 영향을 주는 환경 변수로 생각되지만(Zheng and Wang., 2015) 일차적인 주요 환경 변수에 속하지 않고(Wang et al., 2014), TA보다는 토양 또는 잎 온도가 Js에 더 유의한 영향을 준다고 보고되었다(Pallas et al., 1967; Mellander et al., 2004; Wieser et al., 2015).
Figure 4.
The relationship between sapflow density and environmental variables. Pinus densiflora (left) with VPD(a), Rn(b) and TA(c), and Quercus serrata (right) with VPD(d), Rn(e) and TA(f).
Download Original Figure
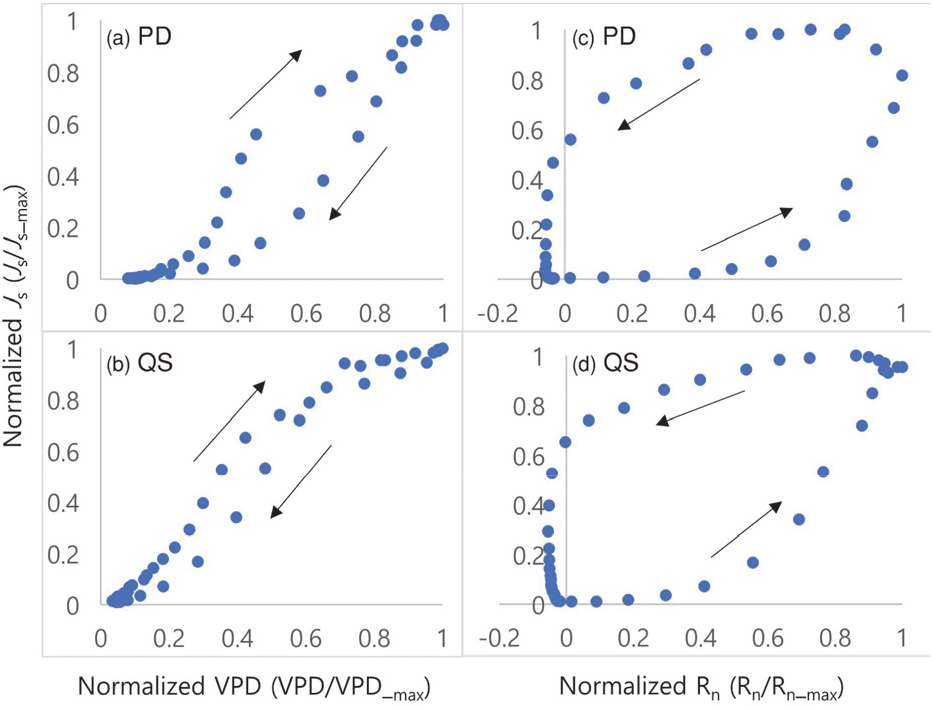
4. 이력현상
Figure 5에서 두 수종의 Js-VPD와 Js-Rn의 이력현상 관계를 나타내었다. 소나무의 경우, 토양수분이 가장 낮았던 5일 동안(186~190일)의 이력현상(Js-VPD)을 보면 동일한 VPD의 값에서 오전의 Js가 오후의 Js에서 보다 높았다. 이는 소나무가 수분의 손실을 줄이기 위해 기공을 조절하기 때문이라고 생각된다. 수목 줄기에 저장된 수분의 소모는 이력현상의 크기를 증대시킬 수 있다는 선행연구와 같이, 줄기에 저장된 수분을 오전에 사용하고 오후에 다시 보충하는 것과 어느 정도가 관계가 있다고 판단된다(Zheng et al., 2014). 반면에 졸참나무는 소나무와 다르게 동일한 VPD에서의 Js의 값이 차이가 없었으며, 이에 이력현상의 크기가 작았다. 그러므로, 졸참나무는 토양 건조에도 기공을 열어두는 anisohydric 방법을 사용하는 것으로 생각된다[Figure 5(b)]. Pappas et al.(2018)에 의해 캐나다 한대림에서 선행되었던 isohydric 수종인 Lalix laricina와 anisohydric 수종인 Picea mariana의 이력현상을 비교한 연구와 유사한 결과이다.
Figure 5.
Location of study site, Pocheon. The Red square indicates the study site. The blue and yellow circles in the red square represent Pinus densiflora subplot and Quercus serrata subplot, respectively.
Download Original Figure
Js-VPD의 hysteresis는 시계 방향으로 이동하였고[Figure 5(a) and 5(b)], Js-Rn의 이력현상은 반시계 방향으로 이동을 하였다(Zeppel et al., 2004; Pappas et al., 2018: Oogathoo et al., 2020)[Figure 5(c) and 5(d)]. Zeppel et al.(2004)은 VPD와 Rn에 의한 이력현상 방향이 다른 것에 대해 Rn과 VPD가 최고치에 도달하는 시차 때문이라고 설명하였다.
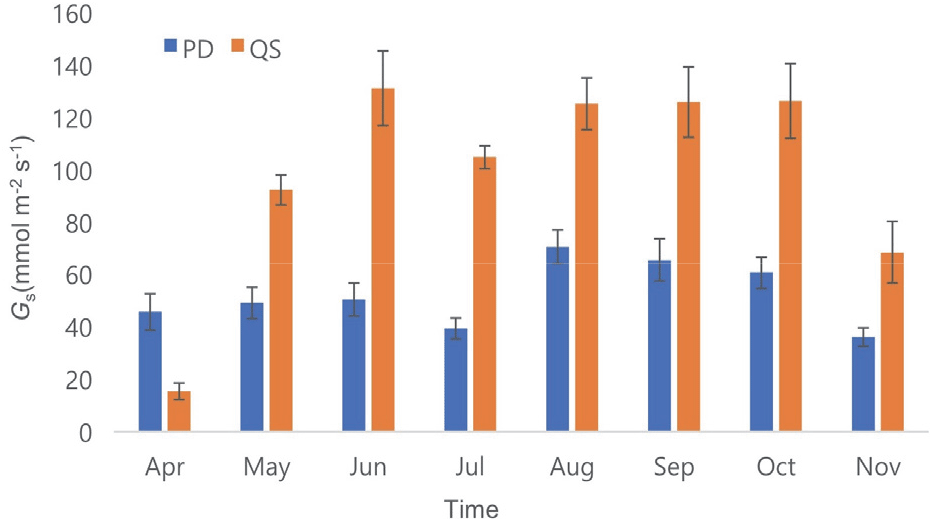
5. 수관전도도
수관전도도(diurnal crown conductance)는 수액속밀도와 비슷한 경향을 나타내었다(Figure 6). 졸참나무의 수관전도도는 토양수분이 감소하는 5~6월과 9~10월에도 감소하지 않고 상대적으로 비슷한 값들을 보였다. 이는 토양수분의 손실에도 기공을 여전히 열어둔다는 것을 의미한다. Schäfer(2011)는 참나무류 세 수종을 대상으로 건조 시기와 건조 시기 전의 수관전도도를 비교하였다. 건조 시기 전인 6월에는 세 수종의 수관전도도가 PPFD(photosynthetic photon flux density)의 증가와 비례하여 계속 증가하지만, 건조 시기인 8월에는 PPFD의 증가에도 불구하고 수관전도도는 감소하거나 증가하지 않았다. 이는 기공을 조절하여 과도한 수분 손실을 줄이고 hydraulic failure을 방지하기 위함이라고 설명하였다.
Figure 6.
Monthly crown conductance between PD and QS. PD and QS represent Pinus densiflora and Quercus serrata, respectively. The error bars indicate standard errors.
Download Original Figure
졸참나무의 수관전도도는 개엽 전인 4월을 제외한 모든 달에서 소나무의 수관전도도보다 유의하게 높았다(p<0.05). 소나무의 수관전도도는 8월에 70.7 mmol m−1 s−1로 가장 높았고 7월에 39.3 mmol m−1 s−1으로 가장 낮았다. 그러나, 졸참나무는 6월에 131.2 mmol m−2 s−1로 가장 높았고 5월에 92.4 mmol m−1 s−1로 가장 낮았다. 소나무와 졸참나무는 각각 8월과 6월에 가장 높은 수관전도도를 보였지만 다른 달과 비교하여 통계적으로 차이가 없었다. 소나무의 수관전도도가 7월에 가장 낮았던 것은 토양 건조가 가장 심했고(SWC<0.25), 또 그 후 강우가 집중되어 RH가 높아지고 이에 VPD가 낮아졌기 때문이라고 생각된다. 졸참나무의 경우, 5월에 가장 낮았는데 이는 개엽이 시작되는 시기이기 때문이라고 생각된다.
결 론
본 연구에서는 우리나라에 우점하고 있는 대표적인 두 수종의 수분 이용 전략을 비교 분석하였다. 두 수종의 수분포텐셜은 측정하지 못했지만 수액속밀도, 이력현상, 그리고 수관전도도를 이용하여 두 수종의 수분 이용 전략을 분석하였다. 소나무는 건조에 대응해 기공을 조절하여 수분 손실을 줄이는 isohydric 방법을 이용하였고, 졸참나무는 기공을 지속해서 열어두어 증산을 유지하는 anisohydric 방법을 이용하지만 환공재라는 구조적 특성과 함께 수액속밀도를 유지하며 건조에 반응하였다. 이 연구에서는 두 수종의 수액속밀도와 수관전도도만을 비교하였지만, 향후 기후변화에 대응하는 수목들의 수분 이용의 명확한 기작을 연구하기 위해서는 탄소동위원소, 수분이용효율, 그리고 광합성과 같은 다른 생리적 특성뿐만 아니라 잎, 수관 등과 같은 형태적 특성들에 관한 연구도 같이 수행되어야 할 것이다.
감사의 글
본 연구는 국립산림과학원의 “환경변화 및 산림교란에 대응한 소나무림 보전 · 관리 전략 및 기술 개발 연구(과제번호: FE0100-2019-05)”의 지원으로 수행되었습니다.
References
Chang, H., Han, S.H., An, J., Park, M.J. and Son, Y. 2019. Relationship between soil water and physiological and growth responses of
Pinus densiflora seedlings under open-field experimental warming and precipitation manipulation. Journal of Climate Change Research 10(2): 145-152.


Chen, L., Zhang, Z., Li, Z., Tang, J., Caldwell, P. and Zhang, W. 2011. Biophysical control of whole tree transpiration under an urban environment in Northern China. Journal of Hydrology 402(3-4): 388-400.
Chun, J.H. and Lee, C.B. 2013. Assessing the effects of climate change on the geographic distribution of
Pinus densiflora in Korea using ecological niche model. Korean Journal of Agricultural and Forest Meteorology 15(4): 219-233.
Ewers, B.E., Gower, S.T., Bond-Lamberty, B. and Wang, C.K. 2005. Effects of stand age and tree species on canopy transpiration and average stomatal conductance of boreal forests. Plant, Cell & Environment 28(5): 660-678.
Ewers, B.E. and Oren, R. 2000. Analyses of assumptions and errors in the calculation of stomatal conductance from sap flux measurements. Tree Physiology 20(9): 579-589.
,

Fauset, S., Freitas, H.C., Galbraith, D.R., Sullivan, M.J., Aidar, M.P., Joly, C.A. and Gloor, M.U. 2018. Differences in leaf thermoregulation and water use strategies between three co-occurring Atlantic forest tree species. Plant, Cell & Environment 41(7): 1618-1631.
,
,

Granier, A. 1987. Evaluation of transpiration in a Douglas-Fir stand by means of sap flow measurements. Tree Physiology 3(7): 309-319.
,
Han, S.H. and Shin, S.J. 2020. Effects of increased temperature and CO
2 concentration on biomass and physiological changes in
Pinus densiflora and
P. thunbergii seedlings. Forest Bioenergy 30(1): 21-28.
Harayama, H., Kitao, M., Agathokleous, E. and Ishida, A. 2019. Effects of major vein blockage and aquaporin inhibition on leaf hydraulics and stomatal conductance. Proceedings of the Royal Society B 286(1904): 20190799.
,
,
Klein, T. 2014. The variability of stomatal sensitivity to leaf water potential across tree species indicates a continuum between isohydric and anisohydric behaviours. Functional Ecology 28(6): 1313-1320.
Ko, S.Y., Sung, J.H., Chun, J.H., Lee, Y.G. and Shin, M.Y. 2014. Predicting the change of yearly productive area distribution for
Pinus densiflora in Korea based on climate change scenarios. Korean Journal of Agricultural and Forest Meteorology 16(1): 72-82.
Korea Forest Research Institute. 2014. Predicting the changes of productive areas for major tree species under climate change in Korea. pp. 240.
Lee, C.S., Lee, W.K., Yoon, J.H. and Song, C.C. 2006. Distribution pattern of
Pinus densiflora and
Quercus spp. stand in Korea using spatial statistics and GIS. Journal of Korean Forest Society 95(6): 663-671.
Lee, S.J., Han, S., Yoon, T.K., Jo, W., Han, S.H., Jung, Y. and Son, Y. 2013. Changes in chlorophyll contents and net photosynthesis rate of 3-year-old
Quercus variabilis seedlings by experimental warming. Journal of Korean Society of Forest Science 102(1): 156-160.
Lee, W.K., Gadow, K.v., Chung, D.J., Lee, J.L. and Shin, M.Y. 2004. DBH growth model for
Pinus densiflora and
Quercus variabilis mixed forests in central Korea. Ecological Modeling 176(1/2): 187-200.
Lee, W.K., Seo, J.H. and Bae, S.W. 2000. Maximum stem number and mortality model for even-aged
Pinus densiflora stand in Kangwon-Province, Korea. Journal of Korean Forest Society 89(5): 634-644.
Lee, W.K., Seo, J.H., Son, Y.M., Lee, K.H. and Gadow, K.v. 2003. Modeling stem profiles for
Pinus densiflora in Korea. Forest Management and Ecology 172(1): 67-77.
Lee, Y.G., Sung, J.H., Chun, J.H. and Shin, M.Y. 2014. Effect of climate changes on the distribution of productive areas for
Quercus mongolica in Korea. Journal of Korean Forest Society 103(4): 605-612.
Liu, X., Zhang, B., Zhuang, J.Y., Han, C., Zhai, L., Zhao, W.R. and Zhang, J.C. 2017. The relationship between sap flow density and environmental factors in the Yangtze River delta region of China. Forests 8(3): 74.
Martin-Benito, D., Anchukaitis, K.J., Evans, M.N., Del Río, M., Beeckman, H. and Cañellas, I. 2017. Effects of drought on xylem anatomy and water-use efficiency of two co-occurring pine species. Forests 8(9): 332332.
Matyssek, R., Kozovits, A.R., Wieser, G., Augustaitiene, I. and Augustaitis, A. 2014. Biological reactions of forests to climate change and air pollution. European Journal of Forest Research 133(4): 671-673.
Meinzer, F.C., Woodruff, D.R., Eissenstat, D.M., Lin, H.S., Adams, T.S. and McCulloh, K.A. 2013. Above-and belowground controls on water use by trees of different wood types in an eastern US deciduous forest. Tree Physiology 33(4): 345-356.
,
Mellander, P.E., Bishop, K. and Lundmark, T. 2004. The influence of soil temperature on transpiration: a plot scale manipulation in a young Scots pine stand. Forest Ecology and Management 195(1-2): 15-28.
O’Grady, A.P., Worledge, D. and Battaglia, M. 2008. Constraints on transpiration of
Eucalyptus globulus in southern Tasmania, Australia. Agricultural and Forest Meteorology 148(3): 453-465.
Oogathoo, S., Houle, D., Duchesne, L. and Kneeshaw, D. 2020. Vapour pressure deficit and solar radiation are the major drivers of transpiration of balsam fir and black spruce tree species in humid boreal regions, even during a short-term drought. Agricultural and Forest Meteorology 291: 108063.
Pallas Jr, J., Michel, B.E. and Harris, D.G. 1967. Photosynthesis, transpiration, leaf temperature, and stomatal activity of cotton plants under varying water potentials. Plant Physiology 42(1): 76-88.
,
,
Pappas, C. et al. 2018. Boreal tree hydrodynamics: asynchronous, diverging, yet complementary. Tree Physiology 38(7): 953-964.
,
Park, M.J., Yun, S.J., Yun, H.M., Chang, H., Han, S.H., An, J. and Son, Y. 2016. Effects of open-field artificial warming and precipitation manipulation on physiological characteristics and growth of
Pinus densiflora seedlings. Journal of Climate Change Research 7(1): 9-17.
,
Pearcy, R.W., Ehleringer, J., Mooney, H.A. and Rundel, P.W. 1989. Plant Physiological Ecology: field methods and instrumentation. 1st Ed. Chapman and Hall. London. UK. pp. 457.
Phillips, N. and Oren, R. 1998. A comparison of daily representations of canopy conductance based on two conditional time-averaging methods and the dependence of daily conductance on environmental factors. Annals of Forest Science 55(1-2): 217-235.
R Core Team. 2021. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
https://www.R-project.org/.
Sànchez-Costa, E., Poyatos, R. and Sabaté, S. 2015. Contrasting growth and water use strategies in four co-occurring Mediterranean tree species revealed by concurrent measurements of sap flow and stem diameter variations. Agricultural and Forest Meteorology 207: 24-37.
Seo, J.H., Lee, W.K., Son, Y. and Ham, B.Y. 2001. Dynamic growth model for
Pinus densiflora stands in Anmyun-island. Journal of Korean Society of Forest Science 90(6): 725-733.
Song, W., Park, G.E., Je, S.M., Kim, S.H. and Lim, J.H. 2020. Growth and physiological characteristics of
Abies koreana,
Pinus densiflora,
Quercus serrata seedlings under elevated temperature and CO
2 concentration. Journal of Climate Change Research 11(1): 1-9.
Schäfer, K.V. 2011. Canopy stomatal conductance following drought, disturbance, and death in an upland oak/pine forest of the New Jersey Pine Barrens, USA. Frontiers in Plant Science 2: 15.
,
,
Wang, H., Guan, H., Deng, Z. and Simmons, C.T. 2014. Optimization of canopy conductance models from concurrent measurements of sap flow and stem water potential on Drooping Sheoak in South Australia. Water Resources Research 50(7): 6154-6167.
Wang, H., Tetzlaff., D. and Soulsby, C. 2019. Hysterestic response of sap flow in Scots pine(
Pinus sylvestris) to meteorological forcing in a humid low-energy headwater catchment. Ecohydrology 12(6): e2125.
Wieser, G., Grams, T.E., Matyssek, R., Oberhuber, W. and Gruber, A. 2015. Soil warming increased whole-tree water use of
Pinus cembra at the treeline in the Central Tyrolean Alps. Tree Physiology 35(3): 279-288.
,
,
Zeppel, M.J., Murray, B.R., Barton, C. and Eamus, D. 2004. Seasonal responses of xylem sap velocity to VPD and solar radiation during drought in a stand of native trees in temperate Australia. Functional Plant Biology 31(5): 461-470.
,
Zeppel, M.J., Taylor, A., Macinnis-Ng, C. and Eamus, D. 2010. Rates of nocturnal transpiration in two evergreen temperate woodland species with differing water-use strategies. Tree Physiology 30(8): 988-1000.
,
Zhang, Q., Manzoni, S., Katul, G., Porporato, A. and Yang, D. 2014. The hysteretic evapotranspiration-vapor pressure deficit relation. Journal of Geophysical Research: Biogeosciences 119(2): 125-140.
Zhang, R., Xu, X., Liu, M., Zhang, Y., Xu, C., Yi, R. and Soulsby, C. 2019. Hysteresis in sap flow and its controlling mechanisms for a deciduous broad-leaved tree species in a humid karst region. Science China Earth Sciences 62(11): 1744-1755.
Zheng, C. and Wang, Q. 2015. Seasonal and annual variation in transpiration of a dominant desert species,
Haloxylon ammodendron, in Central Asia up-scaled from sap flow measurement. Ecohydrology 8(5): 948-960.
Zheng, H., Wang, Q., Zhu, X., Li, Y. and Yu, G. 2014. Hysteresis responses of evapotranspiration to meteorological factors at a diel timescale: patterns and causes. PLoS One 9(6): e98857.
,
,
Zhu, L., Hu, Y., Zhao, X., Zeng, X., Zhao, P., Zhang, Z., and Ju, Y. 2017. The impact of drought on sap flow of cooccurring
Liquidambar formosana Hance and
Quercus variabilis Blume in a temperate forest, Central China. Ecohydrology 10(4): e1828.