Article
남산, 오대산, 울릉도 지역의 주요 참나무과 산림식생에 대한 군락구조 비교
윤이슬
1, 송주현
2, 변성엽
3, 김호진
4, 이정은
4, 김지동
5, 윤충원
4,*
Community Structure Comparison of Fagaceae Forest Vegetation in Namsan, Odaesan, and Ulleungdo
I-Seul Yun
1, Ju Hyeon Song
2, Seong Yeob Byeon
3, Ho Jin Kim
4, Jeong Eun Lee
4, Ji-dong Kim
5, Chung-Weon Yun
4,*
1Division of Urban Biodiversity, Korea Arboreta and Gardens Institute, Sejong 30129, Korea
2Division of Forest Conservation and Restoration, Korea Arboreta and Gardens Institute, Sejong 30129, Korea
3Division of Planning and Coordination, Korea Arboreta and Gardens Institute, Sejong 30129, Korea
4Department of Forest Resources, Kongju National University, Kongju 32588, Korea
5Division of Business Management, Korea Arboreta and Gardens Institute, Sejong 30129, Korea
© Copyright 2022 Korean Society of Forest Science. This is an Open-Access article distributed under the terms of the
Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits
unrestricted non-commercial use, distribution, and reproduction in any
medium, provided the original work is properly cited.
Received: Nov 01, 2022; Revised: Nov 22, 2022; Accepted: Nov 24, 2022
Published Online: Dec 31, 2022
요약
한반도의 산림식생은 낙엽참나무림으로 우점되는 대륙형의 한반도형으로 특징지어진다. 따라서 본 연구는 참나무과 산림식생이 우점하는 남산, 오대산, 울릉도 지역을 대상으로 이질적인 생태 특성에 따른 종조성과 환경인자의 상관관계를 비교하였다. 식생조사는 2018년 5월부터 8월까지 수행하였으며 총 75개소의 조사를 실시하였다. 식생유형분류결과 총 7개 유형으로 구분되었으며, 최상위 수준에서는 내륙지역의 대표식생군락인 신갈나무군락과 도서지역의 대표식생군락인 너도밤나무군락으로 구분되었다. 신갈나무군락은 도시의 대표식생인 팥배나무군과 산악지역의 대표식생인 피나무군으로 세분되었다. 지역형에 따른 천이경향을 살펴보고자 중요치와 지표종 분석을 시행한 결과, 도시형과 도서형은 참나무군락으로 유지되며, 산악형의 경우 피나무 및 까치박달 등 활엽수종의 천이가 이루어질 것으로 판단되었다. 식생유형과 입지환경인자와의 CCA 분석결과, 동위도 상에서 종조성의 영향을 미치는 가장 큰 요인은 해발고도로 나타났다. 본 연구를 통해 한반도의 산림생태계 특성을 이해하고 이에 따른 체계적인 보전 및 복원 계획 수립 시 필요한 기초자료를 제공하고자 한다.
Abstract
The forest vegetation of the Korean Peninsula is dominated by deciduous Fagaceae forests. The study aimed to comparethe correlation between species composition and environmental factors in Namsan, Odaesan, and Ulleungdo. A vegetation survey of 75 sites was conducted from May to August 2018. Seven vegetation types were classified. The inland representative vegetation was classified as a Quercus mongolica community, and the island representative vegetation was classified as a Fagus multinervis community. The Quercus mongolica community was subdivided into the Aria alnifolia group, representative of cities, and the Tilia amurensis group, representative of mountainous regions. Analysis of important values and indicator species to examine the succession trends according to regional types showed that urban and island forestswere maintained as Fagaceae communities, and that mountainous region foreststransitioned to broadleaf species, such as Tilia amurensis and Carpinus cordata. A CCA analysis of vegetation type and site environmental factors showed that altitude had the biggest effect on species composition at the same latitude. The study results should contribute to a better understanding of the Korean Peninsula forest ecosystem characteristics and provide basic data for establishing a systematic conservation and restoration plan.
Keywords: latitude; phytosociology; Quercus mongolica; Fagus multinervis; CCA; altitude
서 론
생물군계(biome)는 전 세계를 기준으로, 유사한 기후조건에 분포한 자연 식생에 의해 특징 지워진 대규모 생태지역을 말한다(Mucina, 2018). 우리나라의 대표적인 생물군계인 온대림(온대계절림)은 위도 30∼55° 사이에 분포하며, 사계절이 뚜렷하게 구분되는 것이 특징으로, 연평균 강수량 650mm∼3,000mm의 범위를 보인다(Walter, 1975; Molles, 2013). 이러한 온대림은 자연식생 계층구조의 발달 및 물질순환, 천이 등 매우 복잡한 생태구조와 기능을 지니고 있으며(Son et al., 2016), 전체 천이 과정에 있는 복잡한 산림생태계 구조를 이해하기 위해서는 자연적으로 발생하는 임분구조 및 종간 상호작용에 대한 포괄적인 생태학적 정보가 필요하다(Chianucci et al., 2016).
한반도의 산림식생(forest vegetation)은 식생지리학적으로 낙엽참나무림으로 우점되는 대륙형의 한반도형으로 특징지어진다(Kim and Lee, 2006). 국내에서 자생하는 참나무과는 4속 15종이 분포하며, 전체 산림면적의 약 25%를 차지하고 있다(Korea Forest Service, 2018; Korea National Arboretum, 2022a). 참나무과의 분포는 낙엽활엽수림과 상록활엽수림으로 나뉘어 냉온대성 낙엽활엽수림이 한반도 전체에 걸쳐서 발달하고 있다. 특히 상록활엽수림은 남해안과 일부 도서지방을 중심으로 제한적 발달하고 있다(Kim, 1992; Oh et al., 2012). 주로 분포하는 참나무과의 낙엽활엽수종은 참나무속(Quercus), 밤나무속(Castanea), 너도밤나무속(Fagus) 3개의 속으로 나뉘며, 참나무속 6종(Q. mongolica, Q. variabilis, Q. serrata, Q. aliena, Q. acutissima, Q. dentata), 밤나무속 1종(C. crenata), 너도밤나무속 1종(F. multinervis)으로 총 8종이 분포하고 있다(Lee, 1986; Yun, 2016; Korea National Arboretum, 2022b). 이중 내륙에서 최우점하는 참나무과 낙엽활엽수종인 신갈나무는 한국을 대표하는 식생으로 중부지역의 냉온대 낙엽활엽수림대와 산악 정상 부근의 표징종이며 건조한 산악 상부에서는 기후적 극상림으로 분포하고 있다(Yim, 1977; Jang and Yim, 1985), 신갈나무군락은 국내에서 개마고원으로부터 한라산에 이르기까지 전 국토에 널리 분포하고 있는 대표적인 낙엽활엽수림으로(Lee et al., 1993; Cheon et al., 2014), 생태적 지위 폭이 넓음에 따라 현재 우리나라 산림에서 안정된 산림군락으로 인정되고 있다(Lee et al., 1994). 또한 도서지역에 우점하고 있는 낙엽활엽수종인 너도밤나무는 울릉도 지역에 국한되어 자생하며 내음성이 강한 우리나라 특산수종으로(Yun et al., 2011), 너도밤나무군락은 해발 300~400 m 이상에 분포하며 성인봉 원시림 내 한반도에서 보기 드물며 안정된 극상림을 이루고 있다(Yim et al., 1981; Han et al., 1998; Lee et al., 2007).
식생은 지표에서 생육하는 모든 식물의 집단으로 대부분 생육지의 환경에 반응하여 결정된 종들이 모인 집단이 군락의 형태로 존재한다(Lee et al., 2006b). 군락이란 동일한 장소와 시간에 살고 있는 여러 개체군의 연합체를 의미하며(Peter, 2012) 이웃하는 군락들과 종조성적 공유를 통하여 연속적 상호관계를 유지함과 동시에 서식환경의 상이성 수준에 따라 제각기 독특한 종조성을 가진다. 상관이 같아도 군락의 구성이나 생태적 특징이 다르게 나타나 그 기능과 구조에 있어서 뚜렷한 차이를 보인다(Braun-Blanquet, 1932; Kim and Lee, 2006).
본 논문은 참나무과 산림식생 우점림을 대상으로 이질적인 생태 특성에 따른 종조성 및 환경인자의 상관관계를 비교하고자 동위도상(N 37°)에 위치하는 남산, 오대산, 울릉도 지역을 대상으로 연구를 진행하였다. 이는 유사 군락의 산림생태계를 이해하는 중요한 역할을 할 수 있으며, 체계적 보전과 복원의 효율적 관리를 위한 계획 수립에 요구되는 기초자료를 제공하고자 수행하였다.
재료 및 방법
1. 연구대상지
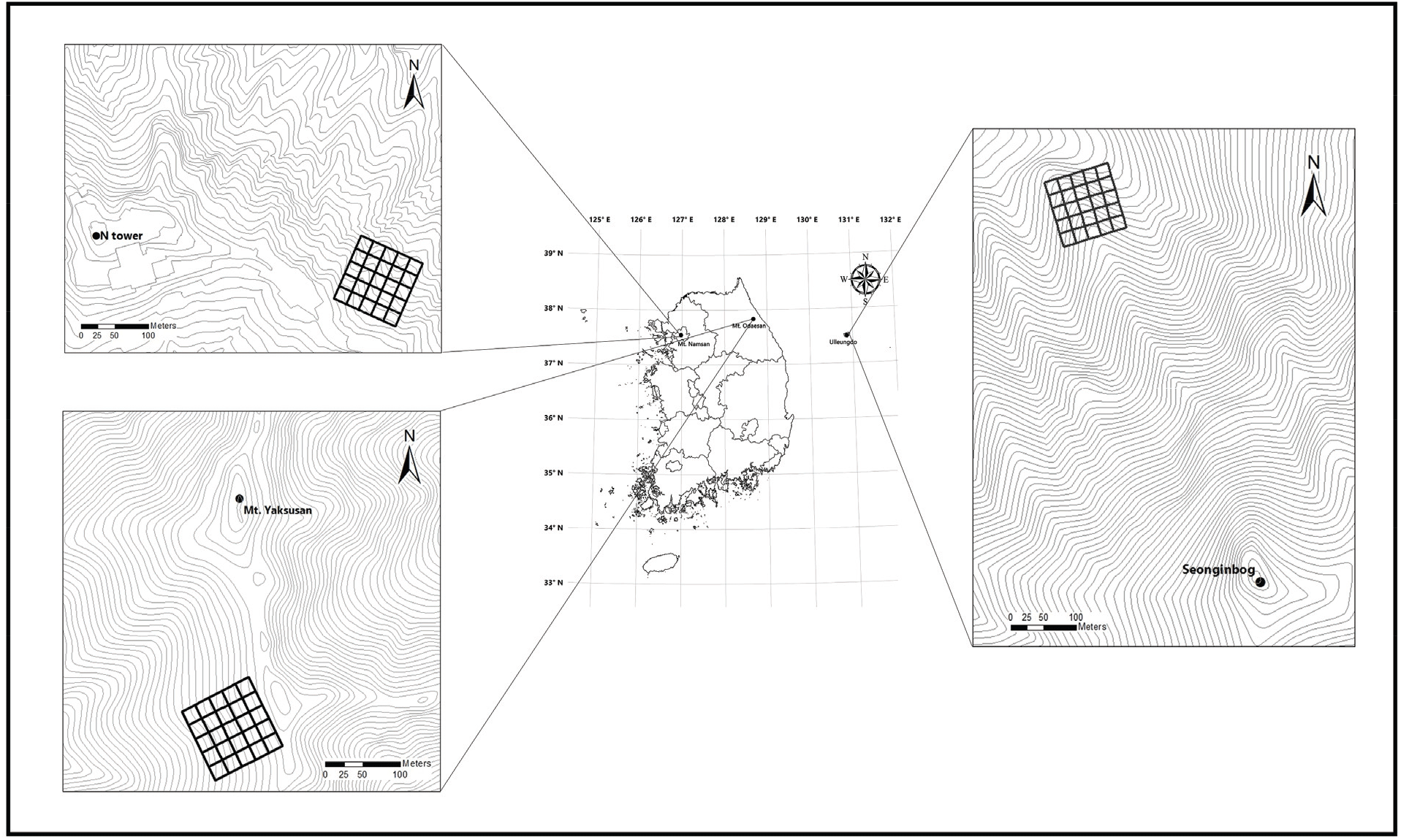
본 연구는 한반도 N 37°에 위치하는 도시림의 남산, 산악림의 오대산, 도서림의 울릉도를 연구대상지로 선정하였다(Figure 1).
남산은 서울의 중심부에 위치한 도시림으로서 행정구역상 서울특별시 중구와 용산구 2개구에 걸쳐 위치하고 있다(Seoul, 2020). 남산의 식생은 식물구계학적으로 한반도 중부아구에 속하며, 중부산야생태권역으로 구분된다(Shin and Kim, 1996; Lee and Yim, 2002). 주 우점군락은 사면에 따라 북사면에 신갈나무군락과 남사면에 소나무군락이 주로 분포하며, 부분적으로 아까시나무군락과 느티나무군락이 인위적으로 조성되어있다(Lee et al., 1998). 오대산은 백두대간의 설악산권역과 태백산권역 중간에 위치하는 산악림으로 행정구역상 강원도 평창군, 홍천군, 강릉시 등 3개 시·군에 걸쳐 있으며, 주요 봉우리로는 주봉인 비로봉(1,563 m)을 비롯하여 동대산(1,433 m), 두로봉(1,421 m), 상왕봉(1,491 m), 호령봉(1,566 m) 등이 병풍처럼 늘어서 있고 노인봉(1,338 m)이 동쪽으로 따로 떨어져 분포하고 있다(Odaesan National Park Office, 2011). 오대산의 식생은 식물구계학적으로 남산과 동일한 한반도 중부아구에 속하며, 백두대간을 따라 분류하는 산악권역으로 구분된다. 주 식생은 산지낙엽활엽수림대가 발달하여 계곡 등 일부 지역을 제외하고 주로 신갈나무가 우점하고, 사면 저지대 및 계곡부에는 물푸레나무, 들메나무, 층층나무 등이 분포한다(Lee et al., 1996). 울릉도는 신생대 대규모 화산활동으로 형성된 도서림으로 행정구역상 경상북도 울릉군 북면에 위치하고 있다. 산지 경사 대부분 30~50°로 급하며, 주봉인 성인봉(983.6 m)를 중심으로 해발 500 m이하가 전체 면적의 78%를 차지하고 있다(Lee et al., 2006b). 울릉도의 식생은 식생지리학적으로 동북아시아 식생형 중 울릉도아형으로 구분되며, 식물구계학적으로 독립된 울릉도아구에 포함된다. 산림식생형은 냉온대활엽수림으로 울릉도를 대표하는 군락인 너도밤나무-섬노루귀군락과 섬잣나무-울릉솔송나무군락이 분포한다(Kim and Lee, 2006).
2. 조사방법
해당 조사는 2018년 5월부터 2018년 8월까지 남산, 오대산, 울릉도의 장기생태모니터링 구간 100 m × 100 m 내의 소방형구 25개(20 m × 20 m)를 조사구로 선정하여 총 75개소의 방형구를 대상으로 Z-M학파의 식물사회학적 방법(Ellenberg, 1956; Braun-Blanquet, 1964)에 따른 식생조사를 실시하였다. 식생조사법에 따라 조사구 내에 출현하는 모든 식물종의 양과 생육상태에 대한 측정을 실시하였고, 표본조사구에서 출현하는 모든 종의 피도(Coverage)와 개체수(Abundance)를 조합시킨 우점도(dominance) 계급을 판정하여 기록하였으며, 생육상태는 종 개체의 집합 혹은 이산의 정도에 따른 군도(Sociability)를 판정하여 기록하였다. 구성종의 식물분류와 동정은 원색한국수목도감(Hong et al., 1987), 원색대한식물도감(Lee, 2003), 나무생태도감(Yun, 2016)을 기준으로 실시하였으며, 종명의 학명과 국명은 국가생물종지식정보시스템(Korea National Arboretum, 2022a)과 국가표준식물목록(Korea National Arboretum, 2022b)을 기준으로 작성하였다.
3. 분석방법
남산, 오대산, 울릉도 3지역에서 조사된 75개소 방형구의 대별종군과 식별종군의 용이한 파악을 위하여 PC-ORD v.7 프로그램을 통해 Hill(1979)의 TWINSPAN (Two-Way Indicator Species Analysis)과 Ellenberg(1956)의 표조작법(Tabulation method)을 이용하여 식생유형을 구분하였다. 입지환경은 해발, 방위, 경사도 등을 측정하였으며, 출현종은 각 층위별 식피율, 평균수고, 평균흉고직경, 우점종을 기록하였다. 식생유형을 구분한 후 정량적 결과산출이 불가능한 정보인 지형은 계곡 10, 사면하부 30, 사면중부 50, 사면상부 70, 능선부와 산정상부 90, 방위는 사면향이 남-남동 10, 남-남서와 남동-동 30, 남서-서와 동-북동 50, 서-북서와 북동-북 70 그리고 북-북서는 90으로 정량적인 값을 주어 분석하였다. 식생단위(Vegetation unit)의 층위별 구성종의 상대적 점유정도를 파악하기 위해 Curtis and McIntosh(1951)의 방법으로 상대빈도(RF: Relative Frequency), 상대피도(RC: Relative Coverage)를 합산한 중요치(I.V: Importance Value)를 산출하였다. 또한 구성종의 층위 간 개체 크기를 고려해 교목층 3, 아교목층 2, 관목층 1, 초본층 0.5의 가중치를 부여한 평균상대우점치(M.I.V: Mean Importance Values)를 환산 비교하였다. 지표종은 공동체 유형, 서식지 조건 또는 환경변화에 대하여 생태적 지표로 사용될 수 있는 종으로(Niemi and McDonald, 2004; De Cáceres et al., 2010), 집단의 서식지환경 또는 승계상태와 같은 생태계의 정성적 특성을 나타내기 위해 지표종분석(ISA: Indicator species analysis)을 실시하였다(Dufrêne and Legendre, 1997). 지표종 분석의 통계적 유의성 평가는 Monte Carlo method를 사용하였다(McCune and Grace, 2002). Ordination 분석 기법을 이용하여 환경요인과의 상관관계가 정량적으로 나타나고 객관적으로 판단하기 적합한 CCA(Canonical Correspondence Analysis) 분석을 실시하였다(Cut off environmental factors R2=0.2).
결 과
1. 종조성에 따른 식물사회학적 식생유형분류
남산, 오대산, 울릉도에서 조사된 75개소의 산림식생조사 자료를 바탕으로 Z-M식물사회학적 방법에 의한 식생유형분류를 수행한 결과 2개 군락(Community), 4개 군(Group), 6개 소군(Subgroup)으로 총 7개 유형이 구분되었다(Table 1). 최상위 단위인 군락 수준에서는 신갈나무군락(Quercus mongolica community)과 너도밤나무군락(Fagus multinervis community)이 분류되었으며, 군 수준에서는 신갈나무군락의 하위 단위로 팥배나무군(Aria alnifolia group)과 피나무군(Tilia amurensis group)이 세분되었고 너도밤나무군락의 하위 단위로는 황벽나무군(Phellodendron amurense group)과 울릉솔송나무군(Tsuga ulleungensis group)이 세분되었다. 소군 수준에서는 팥배나무군의 하위 단위로 진달래소군(Rhododendron mucronulatum subgroup)과 느티나무소군(Zelkova serrata subgroup)으로 세분되었으며 피나무군의 하위 단위로 모시대소군(Adenophora remotiflora subgroup)과 피나무전형소군(T. amurensis typical subgroup)으로 세분되었고 황벽나무군의 하위 단위로 섬쥐똥나무소군(Ligustrum foliosum subgroup)과 선밀나무소군(Smilax nipponica subgroup)으로 세분되었다.
Table 1.
Differentiated constancy table of forest vegetation.
The other 62 companion species omitted.
|
Vegetation Unit System |
|
Community |
A |
B |
|
Group |
a |
b |
a |
b |
|
Subgroup |
i |
ii |
i |
ii |
i |
ii |
|
|
No. of vegetation unit |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
|
Altitude(m)
|
197 |
206 |
1,255 |
1,238 |
615 |
663 |
648 |
|
Topography
|
42.5 |
52.2 |
58.6 |
59.1 |
54.4 |
50.0 |
68.2 |
|
Aspect
|
68.8 |
70.0 |
50.0 |
48.2 |
81.1 |
74.0 |
75.5 |
|
Slope degree(°)
|
7.9 |
6.0 |
15.1 |
14.1 |
33.3 |
33.0 |
33.2 |
|
Bare rock(%)
|
3.7 |
- |
19.8 |
14.1 |
6.4 |
5.0 |
26.5 |
|
Bare soil(%)
|
7.0 |
5.0 |
- |
10.6 |
16.3 |
16.0 |
26.8 |
|
Litter layer(cm)
|
3.6 |
4.8 |
6.7 |
3.6 |
11.4 |
2.1 |
0.9 |
|
Coverage(%)
|
tree layer
|
73.1 |
62.8 |
86.4 |
76.8 |
82.8 |
91.0 |
90.5 |
|
subtree layer
|
48.8 |
41.1 |
65.7 |
50.0 |
25.0 |
38.8 |
46.1 |
|
shrub layer
|
35.1 |
56.1 |
40.0 |
48.6 |
21.7 |
35.0 |
49.5 |
|
herb layer
|
23.8 |
47.6 |
85.0 |
68.2 |
81.1 |
90.0 |
71.8 |
|
Height(m)
|
tree layer
|
14.0 |
14.6 |
14.9 |
14.2 |
21.2 |
19.1 |
17.7 |
|
subtree layer
|
8.8 |
9.8 |
8.5 |
7.7 |
9.1 |
10.3 |
9.0 |
|
shrub layer
|
2.1 |
2.9 |
2.8 |
3.5 |
2.4 |
3.7 |
4.0 |
|
herb layer
|
0.4 |
0.4 |
0.6 |
0.5 |
0.7 |
0.9 |
1.5 |
|
DBH(cm)
|
tree layer
|
23.9 |
24.3 |
34.6 |
29.0 |
44.0 |
33.4 |
26.5 |
|
subtree layer
|
11.9 |
12.4 |
8.9 |
12.7 |
12.4 |
13.7 |
12.7 |
|
shrub layer
|
2.5 |
2.6 |
3.2 |
5.2 |
3.8 |
4.7 |
5.4 |
|
Species diversity
|
1.901±0.2360 |
2.088±0.195 |
2.864±0.128 |
2.563±0.331 |
2.428±0.183 |
2.257±0.223 |
2.040±0.289 |
|
The number of present species
|
21 |
24 |
44 |
39 |
32 |
24 |
20 |
|
Relevé
|
16
|
9
|
14
|
11
|
9
|
5
|
11
|
|
1. Character species and differential species of Quercus mongolica community ;
|
|
Quercus mongolica
|
V24 |
V24 |
V45 |
V24 |
|
|
|
|
Acer pseudosieboldianum
|
V+4 |
V23 |
V13 |
V+3 |
|
|
|
|
Athyrium niponicum
|
III+1 |
III+1 |
V12 |
V13 |
|
|
I++ |
|
Carex siderosticta
|
III+2 |
II+1 |
V13 |
V+3 |
|
|
|
|
Disporum smilacinum
|
V+3 |
V13 |
IV+1 |
I++ |
II++ |
|
|
|
Symplocos sawafutagi
|
III+1 |
II11
|
IV+1
|
V+1 |
|
|
|
|
2. Character species and differential species of Aria alnifolia group ;
|
|
Aria alnifolia
|
V+2 |
V+2 |
I++ |
|
|
|
|
|
Styrax japonicus
|
V+3 |
V+3 |
|
|
|
|
|
|
Parthenocissus tricuspidata
|
V+1 |
V+2 |
|
|
|
|
|
|
Stephanandra incisa
|
V+2 |
V+3 |
II++ |
I++ |
|
|
|
|
Viburnum erosum
|
V+2 |
V+2 |
|
|
|
|
|
|
Callicarpa japonica
|
V+1 |
III++ |
|
|
IV+1 |
|
|
|
Prunus spp. |
IV+2 |
IV+1 |
|
|
|
|
|
|
Euonymus oxyphyllus
|
IV+3 |
IV+3 |
|
|
|
|
|
|
Smilax sieboldii
|
V+1 |
IV+1 |
|
|
|
|
|
|
Dryopteris chinensis
|
IV+1 |
IV+1 |
|
I++ |
II++ |
|
I++ |
|
Lespedeza maximowiczii
|
III+1 |
IV+2 |
|
|
|
|
|
|
Quercus serrata
|
II+4 |
IV+2 |
|
|
|
|
|
|
Alnus incana subsp. hirsuta
|
III+3 |
III+1 |
|
|
|
|
|
|
Toxicodendron trichocarpum
|
II+1 |
III++ |
|
|
|
|
|
|
Zanthoxylum schinifolium
|
II++ |
II++ |
|
|
|
|
|
|
3. Differential species of Rhododendron mucronulatum subgroup ;
|
|
Rhododendron mucronulatum
|
IV+2 |
I++ |
|
|
|
|
|
|
Liriope muscari
|
II++ |
I++ |
|
|
|
|
|
|
4. Differential species of Zelkova serrata subgroup ;
|
|
Zelkova serrata
|
|
III++ |
|
|
|
|
|
|
Vitis amurensis
|
I++ |
III+1 |
|
I++ |
|
|
|
|
Ageratina altissima
|
I++ |
III++ |
|
|
|
|
|
|
Robinia pseudoacacia
|
I++ |
III+1 |
|
|
|
|
|
|
Magnolia kobus
|
|
II++ |
|
|
|
|
|
|
Ligustrum obtusifolium
|
|
II++ |
|
|
|
|
|
|
Magnolia obovata
|
I++ |
II++ |
|
|
|
|
|
|
Maackia amurensis
|
I++ |
II++ |
|
|
I++ |
|
|
|
5. Character species and differential species of Tilia amurensis group ;
|
|
Tilia amurensis
|
|
|
V+5 |
V14 |
|
|
|
|
Calamagrostis arundinacea
|
I11 |
I++ |
V23 |
V24 |
|
|
|
|
Rhododendron schlippenbachii
|
|
II+1 |
V13 |
V23 |
|
|
|
|
Tripterygium regelii
|
|
|
V+3 |
V+2 |
|
|
|
|
Ainsliaea acerifolia
|
II++ |
I11 |
V12 |
V14 |
|
|
|
|
Schisandra chinensis
|
|
|
V12 |
V+3 |
|
|
|
|
Hepatica asiatica
|
|
|
V12 |
V+3 |
|
|
|
|
Carpinus cordata
|
|
|
V+2 |
V+4 |
|
|
|
|
Pseudostellaria palibiniana
|
|
|
V+2 |
V+2 |
|
|
|
|
Streptopus ovalis
|
|
|
V+2 |
V+1 |
|
|
|
|
Pinus koraiensis
|
I11 |
|
V+2 |
V+1 |
|
|
|
|
Convallaria keiskei
|
|
|
V+2 |
II+2 |
|
|
|
|
Galium maximowiczii
|
|
|
V+1 |
V+1 |
|
|
|
|
Pimpinella brachycarpa
|
|
|
V+1 |
IV+2 |
|
|
|
|
Astilbe chinensis
|
|
|
V+1 |
IV+1 |
|
|
|
|
Veratrum maackii var. japonicum
|
|
|
V+1 |
IV+1 |
|
|
|
|
Solidago virgaurea subsp. asiatica
|
|
|
V+1 |
II++ |
|
|
|
|
Aster scaber
|
|
|
IV12 |
IV+1 |
|
|
|
|
Weigela spp. |
I++ |
I++ |
IV+2 |
IV+1 |
|
|
|
|
Isodon inflexus
|
|
|
IV+2 |
V+2 |
|
|
|
|
Ligularia fischeri
|
|
|
IV+2 |
III+1 |
|
|
|
|
Saussurea grandifolia
|
|
|
IV+2 |
III+1 |
|
|
|
|
Acer komarovii
|
|
|
IV+2 |
II+2 |
|
|
|
|
Magnolia sieboldii
|
|
|
IV+1 |
IV+2 |
|
|
|
|
Philadelphus schrenkii
|
|
|
IV+1 |
IV++ |
|
|
|
|
Saussurea seoulensis
|
|
|
IV+1 |
III+1 |
|
|
|
|
Angelica decursiva
|
|
|
IV+1 |
III++ |
|
|
|
|
Artemisia stolonifera
|
|
|
IV+1 |
III+2 |
|
|
|
|
Rubia chinensis
|
|
|
IV+1 |
III++ |
|
|
|
|
Asarum sieboldii
|
I++ |
|
IV++ |
IV++ |
|
|
|
|
Angelica gigas
|
|
|
III+1 |
III+1 |
|
|
|
|
Viola collina
|
|
|
III+1 |
III++ |
|
|
|
|
Vicia unijuga
|
|
|
II+1 |
III++ |
|
|
|
|
Corylus sieboldiana var. mandshurica
|
|
|
II+1 |
III+1 |
|
|
|
|
Euonymus sachalinensis
|
|
|
II+1 |
III++ |
|
|
|
|
Clematis urticifolia
|
|
|
II+1 |
II11 |
|
|
|
|
6. Differential species of Adenophora remotifloras subgroup ;
|
|
Adenophora remotiflora
|
|
|
V12 |
I+1 |
|
|
|
|
Viola rossii
|
|
|
II+1 |
|
|
|
|
|
Meehania urticifolia
|
|
|
III+1 |
|
|
|
|
|
Viola orientalis
|
|
|
III+1 |
I++ |
|
|
|
|
Synurus deltoides
|
|
|
III+1 |
I++ |
|
|
|
|
Viola albida
|
|
|
II+1 |
I++ |
|
|
|
|
Paris verticillata
|
|
|
IIr1 |
I++ |
|
|
|
|
Paeonia japonica
|
|
|
II++ |
I++ |
|
|
|
|
Viburnum wrightii
|
|
|
II++ |
|
|
|
|
|
Actinidia polygama
|
|
|
II++ |
I++ |
|
|
|
|
Acer mandshuricum
|
|
|
II++ |
I11 |
|
|
|
|
Maianthemum japonicum
|
|
|
II++ |
I11 |
|
|
|
|
Viola diamantiaca
|
|
|
II+1 |
|
|
|
|
|
7. Character species and differential species of Fagus multinervis community ;
|
|
Fagus multinervis
|
|
|
|
|
V25 |
V15 |
V55 |
|
Dryopteris crassirhizoma
|
|
|
I++ |
II+2 |
V13 |
V+5 |
III+2 |
|
Hydrangea petiolaris
|
|
|
|
|
V12 |
IV+3 |
V+5 |
|
Hepatica maxima
|
|
|
|
|
V12 |
V+2 |
III++ |
|
Arachniodes standishii
|
|
|
|
|
V14 |
IV24 |
III+4 |
|
Schizophragma hydrangeoides
|
|
|
|
|
V13 |
V+4 |
V+4 |
|
Allium microdictyon
|
|
|
|
|
V12 |
IV13 |
V14 |
|
Disporum sessile
|
|
|
|
|
V+2 |
V++ |
II++ |
|
Sorbus commixta
|
|
|
|
|
IV+1 |
IV+3 |
V+4 |
|
Polystichum ovatopaleaceum var. coraiense
|
|
|
|
|
IV+3 |
IV+2 |
IV+1 |
|
Styrax obassia
|
|
|
|
|
IV+1 |
I11 |
V+1 |
|
Prunus takesimensis
|
|
|
|
|
III+2 |
III12 |
IV14 |
|
Maianthemum dilatatum
|
|
|
|
|
III+4 |
II12 |
IV+4 |
|
Tilia insularis
|
|
|
|
|
II+1 |
V+3 |
V+2 |
|
Acer takesimense
|
|
|
|
|
II11 |
II12 |
V+4 |
|
Adiantum pedatum
|
|
|
|
|
II+1 |
II12 |
IIr+ |
|
8. Character species and differential species of Phellodendron amurense group ;
|
|
Phellodendron amurense
|
|
|
|
|
IV+4 |
IV14 |
|
|
Phryma leptostachya var. oblongifolia
|
|
|
|
I11 |
IV+1 |
V+2 |
I11 |
|
Sambucus racemosa subsp. pendula
|
|
|
|
|
IV+1 |
IV++ |
I++ |
|
Actinidia arguta
|
|
I11 |
I11 |
I11 |
IV+2 |
III++ |
|
|
Celtis jessoensis
|
|
|
|
IV+2 |
III+4 |
|
|
|
Trillium tschonoskii
|
|
|
|
IV+1 |
II+1 |
|
|
|
Galium odoratum
|
|
|
|
|
III+1 |
III+2 |
|
|
Ulmus laciniata
|
|
|
|
|
II+1 |
II22 |
|
|
9. Differential species of Ligustrum foliosum subgroup ;
|
|
Ligustrum foliosum
|
|
|
|
|
V+1 |
|
I++ |
|
Polystichum tripteron
|
|
|
I11 |
I++ |
V12 |
|
I++ |
|
Smilax riparia
|
|
|
|
|
IV+1 |
I++ |
I++ |
|
Hylodesmum podocarpum subsp. oxyphyllum
|
|
|
|
|
IV+1 |
|
|
|
Arisaema takesimense
|
|
|
|
|
IV+1 |
|
|
|
Gynostemma pentaphyllum
|
|
|
|
|
III++ |
|
|
|
Leptogramma pozoi subsp. mollisima
|
|
|
|
|
IV+2 |
|
|
|
Hedera rhombea
|
|
|
|
|
III+1 |
|
|
|
Athyrium acutipinnulum
|
|
|
|
|
III12 |
I++ |
I11 |
|
Ampelopsis glandulosa var. heterophylla
|
|
|
I++ |
|
III+1 |
|
I++ |
|
Pentarhizidium orientale
|
|
|
|
|
III11 |
|
I++ |
|
Alangium platanifolium var. trilobum
|
|
|
|
III++ |
I++ |
|
|
|
Polystichum polyblepharum
|
|
|
|
|
II12 |
|
|
|
Boehmeria tricuspis
|
|
|
|
|
II++ |
|
|
|
Galearis camtschatica
|
|
|
|
|
II++ |
|
|
|
Viola keiskei
|
|
|
I++ |
|
II++ |
I++ |
|
|
10. Differential species of Smilax nipponica subgroup ;
|
|
Smilax nipponica
|
I++ |
|
|
|
|
IV++ |
I++ |
|
Arisaema serratum
|
|
|
|
|
|
IV++ |
|
|
Hylodesmum podocarpum var. mandshuricum
|
|
|
|
|
|
III+1 |
I++ |
|
Aruncus dioicus
|
|
|
|
|
|
II++ |
I++ |
|
Arisaema heterophyllum
|
|
|
|
|
I++ |
II++ |
|
|
Viburnum carlesii
|
|
|
|
|
|
II11 |
I+1 |
|
11. Character species and differential species of Tsuga ulleungensis group ;
|
|
Tsuga ulleungensis
|
|
|
|
|
|
|
V+3 |
|
Taxus cuspidata
|
|
|
|
|
II+1 |
|
IV+2 |
|
Rhododendron brachycarpum
|
|
|
|
|
|
|
V+3 |
|
Mitchella undulata
|
|
|
|
|
I11 |
|
IV+1 |
|
Viburnum furcatum
|
|
|
|
|
I++ |
|
III+1 |
|
Alnus alnobetula subsp. fruticosa
|
|
|
|
|
|
|
II12 |
|
Pyrola renifolia
|
|
|
|
|
|
|
II++ |
|
Campanula takesimana
|
|
|
|
|
|
|
II++ |
|
12. Companion species group;
|
|
Kalopanax septemlobus
|
II+1 |
IV+1 |
III++ |
II++ |
I++ |
|
III+2 |
|
Polygonatum odoratum var. pluriflorum
|
III+1 |
II++ |
II++ |
II++ |
I++ |
|
|
|
Aralia elata
|
II+1 |
IV++ |
III++ |
II++ |
|
|
|
|
Fraxinus rhynchophylla
|
II+1 |
I++ |
II++ |
I++ |
|
|
|
|
Sambucus williamsii var. coreana
|
I++ |
III+1 |
II++ |
III++ |
|
|
|
|
Athyrium yokoscense
|
I+1 |
III++ |
II+1 |
|
|
|
|
|
Carex humilis var. nana
|
III+1 |
II++ |
|
II+2 |
|
|
|
|
Acer pictum var. mono
|
Irr |
|
IV+1 |
III+2 |
V13 |
V+5 |
III13 |
|
Cornus controversa
|
|
III+1 |
I+1 |
I12 |
V12 |
III+2 |
II12 |
|
Osmunda japonica
|
I++ |
I++ |
|
|
|
III24 |
II+3 |
|
Morus bombycis
|
|
II++ |
|
|
II++ |
|
|
|
Carex lanceolata
|
|
II+1 |
II11 |
|
|
|
|
|
Angelica anomala
|
|
|
|
II++ |
|
|
|
|
Adenophora grandiflora
|
|
|
|
II++ |
|
|
|
|
Actaea heracleifolia
|
|
|
I+1 |
II+1 |
|
|
|
|
Arisaema amurensef.serratum
|
|
|
|
II++ |
|
|
|
|
Euonymus fortunei var. radicans
|
|
I++ |
|
|
I++ |
I++ |
|
|
Lactuca triangulata
|
|
|
I++ |
|
I++ |
|
|
|
Sanicula chinensis
|
|
|
|
I++ |
|
I++ |
|
|
Rhododendron yedoensef.poukhanense
|
I+1 |
I++ |
|
|
|
|
|
|
Lespedeza bicolor
|
I+1 |
I++ |
|
|
|
|
|
|
Celastrus orbiculatus
|
I++ |
I++ |
|
|
|
|
|
|
Zanthoxylum piperitum
|
I11 |
I++ |
|
|
|
|
|
|
Abies nephrolepis
|
|
|
I+1 |
I11 |
|
|
|
|
Lilium distichum
|
|
|
I++ |
I++ |
|
|
|
|
Deparia japonica
|
|
|
|
I11 |
I11 |
|
|
|
Deutzia glabrata
|
|
|
I11 |
I12 |
|
|
|
|
Rubia argyi
|
I++ |
|
|
I++ |
|
|
|
|
Deparia conilii
|
I++ |
|
|
I++ |
|
|
|
|
Aconitum pseudolaeve
|
|
|
I+1 |
I++ |
|
|
|
|
Pimpinella komarovii
|
|
|
I++ |
I++ |
|
|
|
|
Adenophora triphylla var. japonica
|
|
|
Irr |
I++ |
|
|
|
|
Rhus chinensis
|
I++ |
I++ |
|
|
|
|
|
|
Abies holophylla
|
|
|
I++ |
I++ |
|
|
|
|
Thalictrum actaefolium var. brevistylum
|
|
|
I++ |
I++ |
|
|
|
|
Betula ermanii
|
|
|
I11 |
I11 |
|
|
|
Download Excel Table
1) 도시형
① 식생단위 1(신갈나무군락-팥배나무군-진달래소군)
본 식생단위는 신갈나무군락에서 종군 2의 팥배나무, 때죽나무, 담쟁이덩굴, 국수나무, 덜꿩나무 등 총 15종의 출현에 의하여 팥배나무군으로 분류되었으며, 팥배나무군의 하위 식생단위로 종군 3의 진달래, 맥문동의 식별종에 의해 진달래소군으로 세분되었다. 식물의 우점도가 3이상인 종은 신갈나무, 당단풍나무, 애기나리, 때죽나무, 참회나무 등 총 7종이 나타났고 상재도가 IV이상의 광역분포특징을 보이는 종은 신갈나무, 당단풍나무, 애기나리, 팥배나무, 때죽나무, 덜꿩나무 등 총 14종이 나타났다. 입지환경요인은 평균해발고 197 m, 지형은 주로 계곡과 사면중부에 분포하였다. 사면방위는 북동사면에 위치하였으며, 평균사면경사도는 7.9°, 평균암석노출도는 3.7%로 전체 식생단위 중 가장 낮은 해발고도가 나타났다. Lee (1998)의 연구결과에 따르면 남산 신갈나무군락의 상재도 III이상인 종으로 신갈나무, 진달래, 덜꿩나무, 애기나리 등이 분포하였으며 이는 본 연구와 일치하였다. 또한 팥배나무와 때죽나무는 도시환경에 적응성이 강한 수종(Lee et al, 2006a)으로 특히 팥배나무는 서울에 분포하는 도시림 중 도심 도시림에서 면적비율이 가장 높게 나타나(Kwak, 2011) 도시림을 대표하는 수종으로 사료되었다.
② 식생단위 2(신갈나무군락-팥배나무군-느티나무소군)
본 식생단위는 팥배나무군의 하위 식생단위로 종군 4의 느티나무, 왕머루, 서양등골나물, 아까시나무, 일본목련 등 총 8종의 식별종 출현에 의해 느티나무소군으로 세분되었다. 식생단위에 출현하는 식물의 우점도가 3이상인 종은 신갈나무, 당단풍나무, 애기나리, 때죽나무, 국수나무, 참회나무로 총 6종이 나타났고 상재도가 IV이상의 광역분포특징을 보이는 종은 신갈나무, 당단풍나무, 애기나리, 팥배나무, 때죽나무, 담쟁이덩굴 등 총 16종이 나타났다. 입지환경요인은 평균해발고 206 m, 지형은 주로 계곡, 사면중부에 분포하였다. 사면방위는 북동사면에 위치하였으며, 평균사면경사도는 6.0°, 평균암석노출도는 나타나지 않았다. 식별종인 느티나무는 우리나라 계곡부 식생을 대표하는 수종(Yun et al., 2011; Byeon and Yun, 2017)으로 해당 지역은 계곡과 사면중부가 맞닿는 비교적 습한 지역으로 인해 느티나무소군으로 구분되었다. 또한 도시림 특성상 과도한 인간간섭으로 인하여 자연환경의 변화가 심한 지역인 남산은(Kil et al., 2004) 본 식생단위에서 귀화식물인 서양등골나물, 아까시나무가 상재도 Ⅲ으로 출현하여 이입된 귀화식물의 생태적인 특성을 파악한 체계적인 관리방안이 요구된다. 또한, 최근 도시온도 상승에 의해 확산되고 있는 수종인 일본목련(Kwak, 2011)의 지속적인 모니터링이 필요할 것으로 판단된다.
2) 산악형
① 식생단위 3(신갈나무군락-피나무군-모시대소군)
본 식생단위는 신갈나무군락에서 종군 5의 피나무, 실새풀, 철쭉, 미역줄나무, 단풍취, 오미자, 노루귀, 까치박달, 큰개별꽃, 금강애기나리 등 총 35종의 출현에 의하여 피나무군으로 분류되었으며, 피나무군의 하위 식생단위로 종군 6의 모시대, 고깔제비꽃, 벌깨덩굴, 노랑제비꽃, 수리취, 태백제비꽃, 백작약, 금강제비꽃 등 총 13종의 식별종에 의해 모시대소군으로 세분되었다. 식물의 우점도가 3이상인 종은 신갈나무, 당단풍나무, 대사초, 피나무, 실새풀, 철쭉, 미역줄나무로 총 7종이 나타났고 상재도가 IV이상의 광역분포특징을 보이는 종은 신갈나무, 당단풍나무, 개고사리, 대사초, 애기나리, 노린재나무, 피나무, 참나물, 노루오줌, 바디나물 등 총 38종이 나타났다. 입지환경요인은 평균해발고 1,255 m, 지형은 주로 사면중부와 상부에 분포하였다. 사면방위는 남서사면에 위치하였으며, 평균사면경사도는 15.1°, 평균암석노출도 19.8%로 나타났다. 강원도 점봉산 단목령 지역의 모시대 군락에 대한 기존의 문헌을 따르면 바디나물, 노루오줌, 나비나물, 참나물 등이 수반종으로 나타났으며 본 연구 결과와 유사한 결과를 나타냈다(Choo et al., 2008). 또한 본 식생단위에서 금강애기나리, 태백제비꽃, 백작약, 금강제비꽃 등 희귀식물의 출현으로 보아 보전가치가 높을 것으로 사료되며 오대산의 동대산 식물군락 연구에서 피나무의 피도가 가장 높게 나타난 군락의 초본층에 금강제비꽃이 출현하는 연구결과와 일치하였다(Lee, 2000).
② 식생단위 4(신갈나무군락-피나무군-피나무전형소군)
본 식생단위는 피나무군의 하위 식생단위로 종군 5의 식별종이 출현하지 않아 피나무전형소군으로 구분되었다. 식생단위에 출현하는 식물의 우점도가 3이상인 종은 신갈나무, 당단풍나무, 개고사리, 대사초, 피나무 등 총 11종이 나타났고 상재도가 IV이상의 광역분포특징을 보이는 종은 신갈나무, 당단풍나무, 개고사리, 대사초, 노린재나무, 피나무 등 총 26종이 나타났다. 입지환경요인은 평균해발고 1,238 m, 지형은 주로 사면중부와 상부에 분포하였다. 사면방위는 서사면에 위치하였으며, 평균사면경사도는 14.1°, 평균암석노출도 14.5%로 나타났다. 강원지역 백두대간 천연림 연구에 따르면 강원지역 백두대간 6개 지역인 향로봉, 오대산, 석병산, 두타산, 덕항산 내에 신갈나무가 우점하는 것으로 확인되었고, 그 중 향로봉, 오대산, 석병산, 함백산에 신갈나무-피나무군집이 나타났으며(Hwang et al., 2012), 오대산 기존 연구에서는 오대산의 서쪽지역인 동대산, 비로봉지역에 주로 신갈나무-피나무군집이 나타나(Lee et al., 1996a; Lee et al., 1996b; Han et al., 2015) 본 연구와 유사한 결과를 나타냈다.
3) 도서형
① 식생단위 5(너도밤나무군락-황벽나무군-섬쥐똥나무소군)
본 식생단위는 너도밤나무군락에서 종군 8의 황벽나무, 파리풀, 말오줌나무, 다래, 풍게나무, 큰연영초, 선갈퀴, 난티나무 등 8종의 식별종 출현에 의해 황벽나무군으로 분류되었으며 황벽나무군의 하위 식생단위로 종군 9의 섬쥐똥나무, 십자고사리, 밀나물, 도둑놈의갈고리, 섬남성, 돌외, 진퍼리고사리 등 총 16종의 식별종에 의해 섬쥐똥나무소군으로 세분되었다. 식물의 우점도가 3이상인 종은 너도밤나무, 관중, 일색고사리, 바위수국, 참나도히초미, 큰두루미꽃, 황벽나무, 고로쇠나무로 총 8종이 나타났고 상재도가 IV이상의 광역분포특징을 보이는 종은 너도밤나무, 관중, 등수국, 섬노루귀, 일색고사리, 바위수국, 산마늘, 윤판나물아재비, 마가목, 참나도히초미, 황벽나무, 십자고사리, 진퍼리고사리 등 총 26종이 나타났다. 입지환경요인은 평균해발고 615 m, 지형은 주로 계곡과 사면상부에 분포하였다. 사면방위는 북서, 북동사면에 위치하였으며, 평균사면경사도는 31.3°, 평균암석노출도 6.4%로 나타났다. 본 식생단위의 식별종인 황벽나무는 비교적 습하고 비옥한 산골짜기나 개울가에서 자생하며(Lee et al., 2014) 수광량이 적은 그늘에서 적응력이 높은 큰두루미꽃, 윤판나물아재비(Jung et al., 2010)와 관중, 일색고사리, 십자고사리, 진퍼리고사리, 섬고사리, 개면마 등의 양치식물이 상재도 III이상으로 나타나 비교적 도서형이 다른 식생단위에 비해 습한 곳으로 사료되었다. 또한 북부 독일의 너도밤나무림의 식별종으로 다수의 고사리류(Dryopteris dillatata, Dryopteris fillx-mas, Athyrium fillx-femina, Polystichum aculeatum etc.)가 출현한 선행연구와 일치하였다(Ellenberg, 1996; Lee et al., 2000).
② 식생단위 6(너도밤나무군락-황벽나무군-선밀나물소군)
본 식생단위는 황벽나무군의 하위 식생단위로 종군 10의 선밀나물, 점박이천남성, 애기도둑놈의갈고리, 두루미천남성, 눈개승마, 분꽃나무 등 6종의 식별종 출현에 의해 선밀나물소군으로 세분되었다. 식생단위에 출현하는 식물의 우점도가 3이상인 종은 너도밤나무, 관중, 등수국, 일색고사리, 마가목, 섬피나무, 황벽나무 등 총 12종이 나타났고 상재도가 IV이상의 광역분포특징을 보이는 종은 너도밤나무, 관중, 등수국, 섬노루귀, 일색고사리, 바위수국, 산마늘, 윤판나물아재비, 마가목, 섬피나무, 황벽나무 등 총 17종이 나타났다. 입지환경요인은 평균해발고 663 m, 지형은 주로 사면상부에 분포하였다. 사면방위는 북서사면에 위치하였으며, 평균사면경사도는 33.0°, 평균암석노출도 5.0%로 전체 식생단위 중 가장 높은 해발고도를 나타냈다. 울릉도 성인봉일대 산림군락에는 주로 나타나는 우점종으로 너도밤나무, 마가목, 등수국, 섬피나무 등(Cho et al., 1993; Song et al., 2000)이 출현하였으며, 본 식생단위에서 우점도 3이상으로 나타나 선행연구와 일치하는 경향을 나타냈다.
③ 식생단위 7(너도밤나무군락-울릉솔송나무군)
본 식생단위는 너도밤나무군락에서 종군 11의 울릉솔송나무, 주목, 만병초, 호자덩굴, 분단나무, 덤불오리나무, 콩팥노루발, 섬초롱꽃 등 8종의 식별종 출현에 의해 울릉솔송나무군으로 분류되었다. 식물의 우점도가 3이상인 종은 너도밤나무, 등수국, 일색고사리, 바위수국, 산마늘, 마가목 등 총 13종이 나타났고 상재도가 IV이상의 광역분포특징을 보이는 종은 너도밤나무, 등수국, 바위수국, 산마늘, 마가목, 참나도히초미, 쪽동백나무 등 총 15종이 나타났다. 입지환경요인은 평균해발고 648 m, 지형은 주로 능선과 사면상부에 분포하였다. 사면방위는 북서사면에 위치하였으며, 평균사면경사도는 33.2°, 평균암석노출도 26.5%로 전체 식생단위 중 가장 높은 사면경사도와 암석노출도를 보였다. 울릉도 성인봉일대 원시림의 산림군집구조를 살펴본 Cho et al.(1993)의 연구에서는 너도밤나무-큰두루미꽃군락-섬단풍-산마늘군-회솔나무-울릉솔송나무소군에 상재도 III이상인 너도밤나무, 마가목, 큰두루미꽃, 쪽동백나무, 섬피나무, 섬벚나무, 섬단풍나무, 만병초, 울릉솔송나무 등이 나타났으며, 본 연구에서도 상재도 IV이상으로 유사한 종조성을 나타내었다. 본 식생단위의 식별종인 울릉솔송나무는 울릉도에 자생하는 수종으로(Korea National Arboretum, 2022a) 울릉도 내 분포지로는 서사면 산록 및 산복부와 동사면 산복부의 절벽주변에 나타나는 것으로 보아(Jeong et al., 2010) 울릉도형에 나타나는 식생단위 중 경사가 급하며 암석노출도가 가장 높은 식생단위 7에서 나타난 것으로 사료된다.
2. 식생단위별 중요치
식생단위로 분류된 구성종의 중요도를 파악하기 위해 전체 산림식생의 중요치와 층위별 중요치를 산출하였다(Table 2).
Table 2.
Importance value of major species in each vegetation units.
|
Unit |
Scientific name |
Layer |
MIV |
|
T1 |
T1 |
S |
H |
|
1 |
Quercus mongolica
|
59.3 |
27.8 |
4.1 |
6.7 |
37.1 |
|
Acer pseudosieboldianum
|
- |
35.3 |
14.5 |
3.7 |
13.4 |
|
Aria alnifolia
|
10.5 |
11.2 |
5.7 |
4.2 |
9.5 |
|
Styrax japonicus
|
5.3 |
6.4 |
11.8 |
1.0 |
6.3 |
|
Alnus sibirica
|
8.2 |
2.3 |
2.9 |
0.8 |
5.0 |
|
Quercus serrata
|
9.8 |
1.2 |
- |
0.8 |
5.0 |
|
Prunus spp. |
2.1 |
7.0 |
3.5 |
1.2 |
3.8 |
|
Euonymus oxyphyllus
|
- |
4.6 |
10.5 |
3.4 |
3.3 |
|
Disporum smilacinum
|
- |
- |
- |
28.4 |
2.2 |
|
Stephanandra incisa
|
- |
- |
12.2 |
3.7 |
2.2 |
|
Viburnum erosum
|
- |
- |
9.7 |
2.6 |
1.7 |
|
Kalopanax septemlobus
|
1.6 |
1.2 |
0.6 |
0.6 |
1.2 |
|
Parthenocissus tricuspidata
|
- |
1.8 |
0.6 |
5.0 |
1.0 |
|
Rhododendron mucronulatum
|
- |
- |
5.5 |
0.6 |
0.9 |
|
Pinus densiflora
|
1.6 |
- |
- |
0.4 |
0.8 |
|
Fraxinus sieboldiana
|
1.6 |
- |
- |
0.2 |
0.7 |
|
Callicarpa japonica
|
- |
- |
3.6 |
1.9 |
0.7 |
|
Symplocos sawafutagi
|
- |
- |
3.3 |
0.4 |
0.5 |
|
Others(45 Species) |
- |
1.2 |
11.5 |
34.3 |
4.7 |
|
Total(63 Species) |
100.0 |
100.0 |
100.0 |
100.0 |
100.0 |
|
2 |
Quercus mongolica
|
58.7 |
14.0 |
4.1 |
2.8 |
32.3 |
|
Acer pseudosieboldianum
|
- |
43.0 |
14.9 |
5.3 |
15.9 |
|
Aria alnifolia
|
17.5 |
13.3 |
2.5 |
1.4 |
12.7 |
|
Quercus serrata
|
15.6 |
- |
- |
1.8 |
7.3 |
|
Styrax japonicus
|
2.7 |
5.6 |
12.3 |
1.8 |
5.0 |
|
Kalopanax septemlobus
|
2.7 |
5.7 |
1.7 |
1.8 |
3.4 |
|
Alnus incana subsp. hirsuta
|
2.7 |
4.2 |
2.5 |
0.4 |
3.0 |
|
Stephanandra incisa
|
- |
- |
14.3 |
3.9 |
2.5 |
|
Disporum smilacinum
|
- |
- |
- |
29.9 |
2.3 |
|
Prunus spp. |
- |
4.2 |
2.2 |
1.1 |
1.7 |
|
Cornus controversa
|
- |
4.2 |
2.2 |
0.4 |
1.7 |
|
Robinia pseudoacacia
|
- |
4.2 |
2.1 |
- |
1.6 |
|
Viburnum erosum
|
- |
- |
7.9 |
1.8 |
1.3 |
|
Euonymus oxyphyllus
|
- |
- |
6.3 |
3.9 |
1.3 |
|
Parthenocissus tricuspidata
|
- |
- |
0.9 |
8.9 |
0.8 |
|
Lespedeza maximowiczii
|
- |
- |
3.4 |
1.8 |
0.7 |
|
Clerodendrum trichotomum
|
- |
1.5 |
0.4 |
- |
0.5 |
|
Vitis amurensis
|
- |
- |
2.5 |
1.1 |
0.5 |
|
Others(40 Species) |
- |
- |
19.7 |
32.5 |
5.5 |
|
Total(58 Species) |
100.0 |
100.0 |
100.0 |
100.0 |
100.0 |
|
3 |
Quercus mongolica
|
56.8 |
6.4 |
1.6 |
1.0 |
28.5 |
|
Tilia amurensis
|
26.5 |
16.9 |
7.9 |
1.1 |
18.8 |
|
Acer pseudosieboldianum
|
- |
31.6 |
20.2 |
3.0 |
13.1 |
|
Carpinus cordata
|
2.0 |
31.0 |
11.9 |
1.9 |
12.5 |
|
Pinus koraiensis
|
8.3 |
- |
5.8 |
1.2 |
4.8 |
|
Rhododendron schlippenbachii
|
- |
- |
22.1 |
2.4 |
3.6 |
|
Cornus controversa
|
1.6 |
4.7 |
- |
0.1 |
2.2 |
|
Acer pictum subsp. mono
|
3.2 |
- |
1.5 |
0.8 |
1.7 |
|
Tripterygium regelii
|
- |
2.1 |
0.8 |
4.8 |
1.2 |
|
Magnolia sieboldii
|
- |
- |
6.9 |
0.7 |
1.1 |
|
Symplocos sawafutagi
|
- |
- |
6.4 |
0.9 |
1.0 |
|
Acer komarovii
|
- |
- |
5.1 |
1.8 |
0.9 |
|
Betula ermanii
|
1.6 |
- |
- |
- |
0.7 |
|
Calamagrostis arundinacea
|
- |
- |
- |
9.2 |
0.7 |
|
Euonymus sachalinensis
|
- |
1.1 |
1.8 |
0.2 |
0.6 |
|
Others(86 Species) |
- |
6.4 |
8 |
70.9 |
8.4 |
|
Total(101 Species) |
100.0 |
100.0 |
100.0 |
100.0 |
100.0 |
|
4 |
Quercus mongolica
|
46.2 |
12.1 |
0.5 |
1.1 |
25.2 |
|
Tilia amurensis
|
31.5 |
19.3 |
7.1 |
1.1 |
21.6 |
|
Acer pseudosieboldianum
|
- |
31.2 |
17.2 |
1.6 |
12.4 |
|
Carpinus cordata
|
- |
26.7 |
16.0 |
1.1 |
10.8 |
|
Pinus koraiensis
|
7.6 |
1.3 |
2.4 |
0.7 |
4.4 |
|
Cornus controversa
|
4.5 |
4.0 |
0.8 |
- |
3.4 |
|
Rhododendron schlippenbachii
|
- |
- |
20.4 |
1.7 |
3.3 |
|
Betula ermanii
|
1.9 |
2.7 |
- |
- |
1.7 |
|
Acer pictum var. mono
|
2.6 |
- |
1.9 |
0.4 |
1.5 |
|
Tripterygium regelii
|
- |
1.3 |
3.3 |
3.3 |
1.2 |
|
Calamagrostis arundinacea
|
- |
- |
- |
13.7 |
1.1 |
|
Actinidia arguta
|
1.9 |
- |
0.8 |
- |
1.0 |
|
Acer mandshuricum
|
1.9 |
- |
0.8 |
- |
1.0 |
|
Magnolia sieboldii
|
- |
- |
6.1 |
0.8 |
1.0 |
|
Symplocos sawafutagi
|
- |
- |
5.9 |
0.9 |
1.0 |
|
Abies nephrolepis
|
1.9 |
- |
- |
- |
0.9 |
|
Acer komarovii
|
- |
1.3 |
2.3 |
0.4 |
0.8 |
|
Ainsliaea acerifolia
|
- |
- |
- |
9.9 |
0.8 |
|
Carex siderosticta
|
- |
- |
- |
7.0 |
0.5 |
|
Others(77 Species) |
- |
- |
14.6 |
56.5 |
6.6 |
|
Total(96 Species) |
100.0 |
100.0 |
100.0 |
100.0 |
100.0 |
|
5 |
Fagus multinervis
|
37.4 |
38.6 |
38.5 |
3.0 |
35.3 |
|
Acer pictum var. mono
|
14.4 |
7.6 |
13.0 |
1.8 |
11.1 |
|
Celtis jessoensis
|
7.9 |
14.1 |
8.3 |
0.4 |
9.3 |
|
Phellodendron amurense
|
16.1 |
- |
- |
0.2 |
7.4 |
|
Cornus controversa
|
12.5 |
2.5 |
1.2 |
0.8 |
6.8 |
|
Hydrangea petiolaris
|
- |
14.6 |
- |
4.3 |
4.8 |
|
Schizophragma hydrangeoides
|
1.6 |
10.1 |
2.3 |
7.9 |
4.8 |
|
Sorbus commixta
|
4.7 |
5.0 |
6.0 |
0.4 |
4.7 |
|
Prunus takesimensis
|
5.4 |
2.5 |
1.2 |
0.4 |
3.5 |
|
Acer takesimense
|
- |
5.0 |
- |
- |
1.6 |
|
Tilia insularis
|
- |
- |
5.9 |
0.4 |
0.9 |
|
Styrax obassia
|
- |
- |
4.8 |
1.2 |
0.8 |
|
Callicarpa japonica
|
- |
- |
4.8 |
0.6 |
0.8 |
|
Ulmus laciniata
|
- |
- |
4.7 |
0.6 |
0.8 |
|
Arachniodes standishii
|
- |
- |
- |
9.7 |
0.7 |
|
Taxus cuspidata
|
- |
- |
3.5 |
0.2 |
0.6 |
|
Actinidia arguta
|
- |
- |
2.3 |
2.4 |
0.5 |
|
Ligustrum foliosum
|
- |
- |
2.3 |
1.8 |
0.5 |
|
Others(46 Species) |
- |
- |
1.2 |
63.7 |
5.1 |
|
Total(64 Species) |
100.0 |
100.0 |
100.0 |
100.0 |
100.0 |
|
6 |
Fagus multinervis
|
20.4 |
39.9 |
32.9 |
3.6 |
27.0 |
|
Sorbus commixta
|
11.1 |
17.3 |
6.6 |
1.1 |
11.5 |
|
Tilia insularis
|
5.7 |
17.9 |
11.8 |
0.5 |
10.0 |
|
Acer pictum var. mono
|
17.4 |
- |
10.3 |
1.1 |
9.7 |
|
Phellodendron amurense
|
15.7 |
- |
- |
- |
7.2 |
|
Prunus takesimensis
|
5.7 |
6.5 |
9.9 |
0.5 |
6.2 |
|
Celtis jessoensis
|
8.7 |
4.6 |
1.8 |
- |
5.7 |
|
Hydrangea petiolaris
|
- |
13.7 |
1.8 |
5.8 |
4.9 |
|
Cornus controversa
|
6.5 |
- |
1.8 |
- |
3.3 |
|
Ulmus laciniata
|
6.5 |
- |
- |
- |
3.0 |
|
Acer takesimense
|
2.4 |
- |
9.9 |
0.5 |
2.7 |
|
Schizophragma hydrangeoides
|
- |
- |
1.8 |
12.0 |
1.2 |
|
Viburnum carlesii
|
- |
- |
6.6 |
1.4 |
1.1 |
|
Dryopteris crassirhizoma
|
- |
- |
- |
13.4 |
1.0 |
|
Arachniodes standishii
|
- |
- |
- |
8.1 |
0.6 |
|
Osmunda japonica
|
- |
- |
- |
7.7 |
0.6 |
|
Styrax obassia
|
- |
- |
3.3 |
- |
0.5 |
|
Others(16 Species) |
- |
- |
1.8 |
44.3 |
3.6 |
|
Total(40 Species) |
100.0 |
100.0 |
100.0 |
100.0 |
100.0 |
|
7 |
Fagus multinervis
|
41.4 |
37.1 |
31.9 |
4.1 |
35.7 |
|
Acer takesimense
|
4.9 |
23.4 |
9.9 |
3.3 |
11.3 |
|
Sorbus commixta
|
10.9 |
8.2 |
13.2 |
2.9 |
9.8 |
|
Tsuga ulleungensis
|
1.5 |
7.1 |
21.2 |
2.8 |
6.4 |
|
Prunus takesimensis
|
9.8 |
4.7 |
0.9 |
0.3 |
6.1 |
|
Acer pictum var. mono
|
9.4 |
2.3 |
1.7 |
- |
5.3 |
|
Tilia insularis
|
6.5 |
4.7 |
4.7 |
1.7 |
5.3 |
|
Styrax obassia
|
3.4 |
2.3 |
5.5 |
2.1 |
3.3 |
|
Alnus alnobetula subsp. fruticosai |
4.9 |
- |
1.2 |
- |
2.5 |
|
Cornus controversa
|
2.7 |
3.6 |
- |
- |
2.3 |
|
Kalopanax septemlobus
|
4.6 |
- |
- |
0.6 |
2.2 |
|
Taxus cuspidata
|
- |
2.3 |
7.2 |
1.8 |
2.0 |
|
Schizophragma hydrangeoides
|
- |
1.8 |
0.9 |
13.4 |
1.7 |
|
Hydrangea petiolaris
|
- |
2.3 |
- |
9.6 |
1.5 |
|
Allium microdictyon
|
- |
- |
- |
11.4 |
0.9 |
|
Maianthemum dilatatum
|
- |
- |
- |
10.7 |
0.8 |
|
Others(31 Species) |
- |
- |
1.8 |
35.5 |
3 |
|
Total(47 Species) |
100.0 |
100.0 |
100.0 |
100.0 |
100.0 |
Download Excel Table
1) 도시형
① 식생단위 1(신갈나무군락-팥배나무군-진달래소군)
평균상대우점치는 신갈나무(37.1%), 당단풍나무(13.4%), 팥배나무(9.5%)의 순으로 나타났으며 교목층 중요치는 신갈나무(59.3%)로 가장 높게 나타났고 팥배나무(10.5%), 졸참나무(9.8%) 등의 순으로 나타났다. 아교목층 중요치는 당단풍나무(35.3%)가 가장 높게 나타났고 신갈나무(27.8%), 팥배나무(11.2%), 때죽나무(6.4%) 등의 순으로 나타났다. 관목층 중요치는 당단풍나무(14.5%), 국수나무(12.2%), 때죽나무(11.8%), 참회나무(10.5%) 등의 순으로 나타났다. 초본층 중요치는 애기나리(28.4%)가 가장 높게 나타났으며 신갈나무(6.7%), 담쟁이덩굴(5.0%) 등의 순으로 나타났다. 교목층의 경우 신갈나무의 중요치가 가장 높게 나타났으며 두 번째로 높게 나타난 팥배나무 중요치 값과 높은 차이를 보여 신갈나무가 우세할 것으로 예상된다. 또한 초본층에 나타나는 신갈나무의 중요치 값으로 보아 당분간 신갈나무의 세력이 유지 될 것으로 판단되었다.
② 식생단위 2(신갈나무군락-팥배나무군-느티나무소군)
평균상대우점치는 신갈나무(32.3%), 당단풍나무(15.9%), 팥배나무(12.7%), 졸참나무(7.3%) 순으로 나타났으며 교목층 중요치는 신갈나무(58.7%)가 가장 높게 나타났고 팥배나무(17.5%), 졸참나무(15.6%) 등의 순으로 나타났다. 아교목층 중요치는 당단풍나무(43.0%)가 가장 높게 나타났으며 신갈나무(14.0%), 팥배나무(13.3%) 등의 순으로 나타났다. 관목층 중요치는 당단풍나무(14.9%)가 가장 높게 나타났으며 국수나무(14.3%), 때죽나무(12.3%) 등의 순으로 나타났다. 초본층 중요치는 애기나리(29.9%)가 가장 높게 나타났으며 담쟁이덩굴(8.9%), 당단풍나무(5.3%) 등의 순으로 나타났다. 남산의 신갈나무군락은 교목층과 아교목층에서 신갈나무와 당단풍나무가 우점하며 아교목층과 관목층에 팥배나무, 당단풍나무, 때죽나무 등이 혼생하는 기존 선행연구(Lee et al., 1998a; Seol, 2008)와 비슷한 결과가 나타났으며 지위경쟁 대상의 교목성 수종이 나타나지 않아 신갈나무 군락의 세력이 유지 될 것으로 판단되었다.
2) 산악형
① 식생단위 3(신갈나무군락-피나무군-모시대소군)
평균상대우점치는 신갈나무(28.5%), 피나무(18.8%), 당단풍나무(13.1%) 순으로 나타났으며 교목층 중요치는 신갈나무(56.8%)가 가장 높게 나타났고 피나무(26.5%), 잣나무(8.3%) 등의 순으로 나타났다. 아교목층 중요치는 당단풍나무(31.6%)가 가장 높게 나타났으며 까치박달(31.0%), 피나무(16.9%) 등의 순으로 나타났다. 관목층 중요치는 철쭉(22.1%)이 가장 높게 나타났으며 당단풍나무(20.2%), 까치박달(11.9%) 등의 순으로 나타났다. 초본층 중요치는 실새풀(9.2%)이 가장 높게 나타났으며 대사초(5.0%), 미역줄나무(4.8%) 등의 순으로 나타났다. 오대산지역 평균해발고 700 m 이상의 대표적인 수종인 신갈나무와 피나무(Lee et al., 1996a; Han et al., 2015)가 교목층에 가장 높은 중요치를 나타내며 아교목층에도 높은 중요치로 출현하고 있다. 아교목층에 가장 높은 중요치를 나타내는 당단풍나무는 아교목성의 수종으로 세력을 키우기에 다소 어려움이 있으나, 교목성 수종인 까치박달과 피나무가 차우점종으로 나타나 신갈나무와 지위경쟁이 있을 것으로 사료된다.
② 식생단위 4(신갈나무군락-피나무군-피나무전형소군)
평균상대우점치는 신갈나무(25.2%), 피나무(21.6%), 당단풍나무(12.4%)의 순으로 나타났으며 교목층 중요치는 신갈나무(46.2%)가 가장 높게 나타났고 피나무(31.5%), 잣나무(7.6%) 등의 순으로 나타났다. 아교목층 중요치는 당단풍나무(31.2%)가 가장 높게 나타났으며 까치박달(26.7%), 피나무(19.3%) 등의 순으로 나타났다. 관목층 중요치는 철쭉(20.4%)이 가장 높게 나타났으며 당단풍나무(17.2%), 까치박달(16.0%) 등의 순으로 나타났다. 초본층 중요치는 실새풀(13.7%)이 가장 높게 나타났으며 단풍취(9.9%), 대사초(7.9%) 등의 순으로 나타났다. 신갈나무군락 내 공통적으로 출현하는 식별종인 아교목층의 당단풍나무, 관목층의 철쭉, 초본층의 실새풀, 단풍취, 대사초(Chung et al., 2000; Kim et al., 2009; Song et al., 2019)가 높은 중요치 값을 나타났다. 특히 교목층의 경우 신갈나무의 중요치가 가장 높게 나타났지만 내음성이 강한 피나무의 중요치가 교목층을 제외한 모든 층위에서 신갈나무보다 높은 중요치 값을 보였으며, 아교목층 및 관목층에 분포하는 교목성 까치박달의 중요치 값이 가장 높게 나타나는 점을 비교하여 이후 군락 내 신갈나무의 구성 비율이 줄어들고 피나무 및 까치박달 등 활엽수종들의 천이가 이루어질 것으로 판단된다.
3) 도서형
① 식생단위 5(너도밤나무군락-황벽나무군-섬쥐똥나무소군)
평균상대우점치는 너도밤나무(35.3%), 고로쇠나무(11.1%), 풍게나무(9.3%)의 순으로 나타났으며 교목층 중요치는 너도밤나무(37.4%)로 가장 높게 나타났고 황벽나무(16.1%), 고로쇠나무(14.4%) 등의 순으로 나타났다. 아교목층 중요치는 너도밤나무(38.6%)가 가장 높게 나타났으며 등수국(14.6%), 풍게나무(14.1%) 등의 순으로 나타났다. 관목층 중요치는 너도밤나무(38.5%)가 가장 높게 나타났으며 고로쇠나무(13.0%), 풍게나무(8.3%) 등의 순으로 나타났다. 초본층 중요치는 일색고사리(9.3%)가 가장 높게 나타났으며 바위수국(7.9%), 산마늘(5.1%) 등의 순으로 나타났다. 초본층을 제외한 전 층위에서 너도밤나무의 중요치가 가장 높게 나타나 당분간은 너도밤나무군락으로 유지될 것으로 판단된다.
② 식생단위 6(너도밤나무군락-황벽나무군-선밀나물소군)
평균상대우점치는 너도밤나무(27.0%), 마가목(11.1%), 섬피나무(10.0%)의 순으로 나타났으며 교목층 중요치는 너도밤나무(20.4%)가 가장 높게 나타났고 고로쇠나무(17.4%), 황벽나무(15.7%) 등의 순으로 나타났다. 아교목층 중요치는 너도밤나무(39.9%)가 가장 높게 나타났으며 섬피나무(17.9%), 마가목(17.3%) 등의 순으로 나타났다. 관목층 중요치는 너도밤나무(32.9%)가 가장 높게 나타났으며 섬피나무(11.8%), 고로쇠나무(10.3%) 등의 순으로 나타났다. 초본층 중요치는 관중(13.4%)이 가장 높게 나타났으며 바위수국(12.0%), 일색고사리(8.1%) 등의 순으로 나타났다. 교목층과 아교목층에서 우위를 나타내는 너도밤나무와 두 번째로 높게 출현한 고로쇠나무의 중요치 값이 큰 차이가 나지 않았다. 이는 울릉도에서 고로쇠나무의 생장과 확산이 빠르게 나타나며 일부 군락에서는 우점하는 경향을 나타내므로(Han et al., 1998; Lee et al., 2007; Jeong et al., 2010), 너도밤나무와 고로쇠나무의 지위경쟁이 있을 것으로 사료되었다. 하지만 하층의 치수 비교 시 고로쇠나무에 비해 너도밤나무가 우세함으로 너도밤나무의 세력이 유지될 것으로 판단되었다.
③ 식생단위 7(너도밤나무군락-울릉솔송나무군)
평균상대우점치는 너도밤나무(35.7%), 섬단풍나무(11.3%), 마가목(9.8%)의 순으로 나타났으며 교목층 중요치는 너도밤나무(41.4%)가 가장 높게 나타났고 마가목(10.9%), 섬벚나무(9.8%) 등의 순으로 나타났다. 아교목층 중요치는 너도밤나무(37.1%)가 가장 높게 나타났으며 섬단풍나무(23.4%), 마가목(8.2%) 등의 순으로 나타났다. 관목층 중요치는 너도밤나무(31.9%)가 가장 높게 나타났으며 울릉솔송나무(21.2%), 마가목(13.2%) 등의 순으로 나타났다. 초본층 중요치는 바위수국(13.4%)이 가장 높게 나타났으며 산마늘(11.4%), 큰두루미꽃(10.7%) 등의 순으로 나타났다. 교목층에서 중요치가 가장 높은 수종인 너도밤나무가 아교목층과 관목층에도 가장 높은 중요치를 나타내어 당분간 너도밤나무의 세력이 유지 될 것으로 사료되었다.
3. 지표종분석
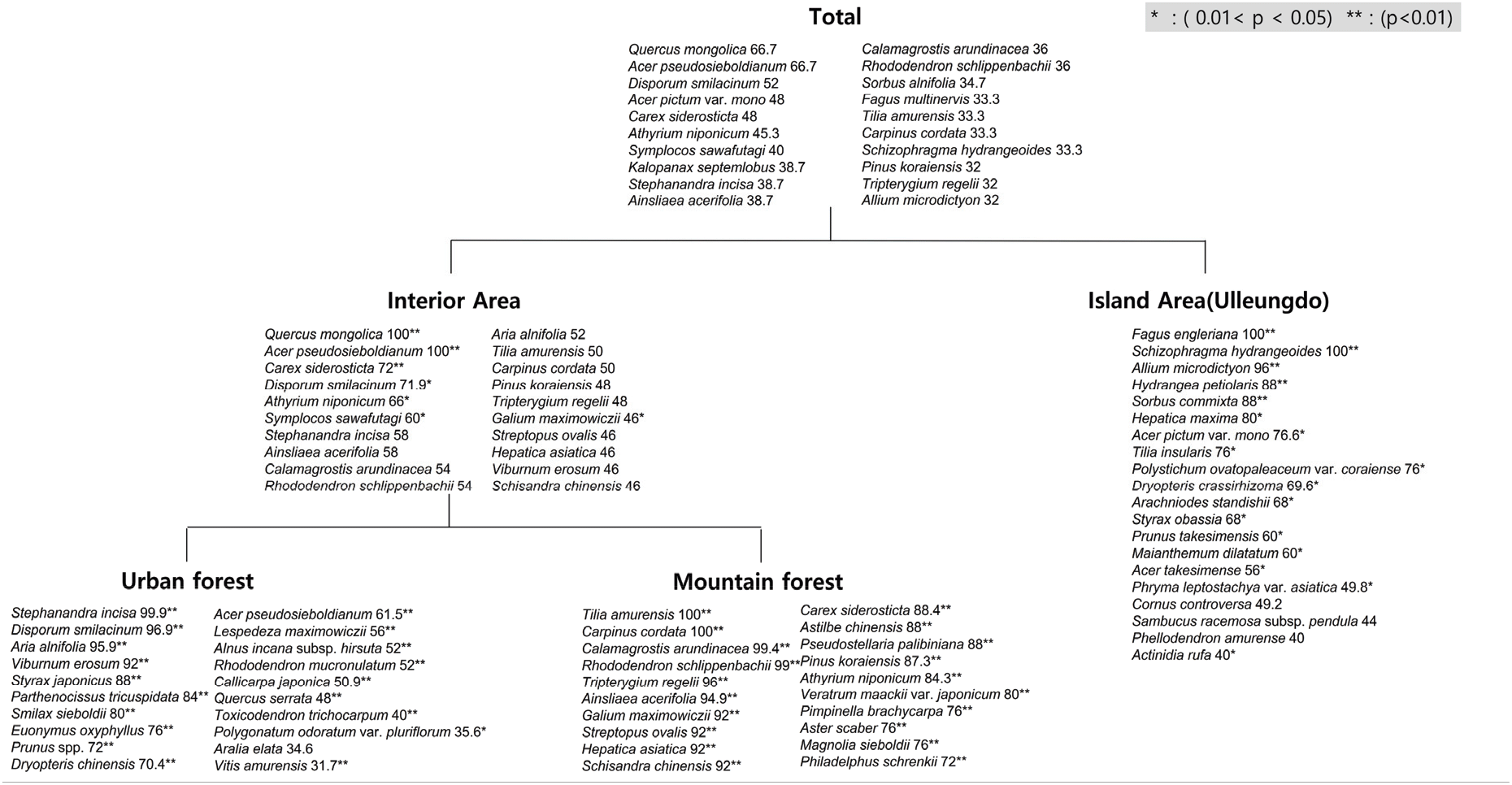
조사지점 내 출현종을 대상으로 지표종 분석을 실시한 결과, 참나무과 산림식생은 내륙지역(Interior Area)과 도서지역(Island Area)으로 구분되었다(Figure 2). 내륙지역의 산림식생은 신갈나무가 우점하는 군락으로 신갈나무(100.0), 당단풍나무(100.0), 대사초(72.0) 3종이, 도서지역의 산림식생은 너도밤나무가 우점하는 군락으로 너도밤나무(100.0), 바위수국(100.0), 산마늘(96.0), 등수국(88.0), 마가목(88.0) 4종이 유의한 지표종으로 나타났다(p<0.01). 내륙지역은 다시 도시형(Urban forest)과 산악형(Mountain forest)으로 나뉘었으며, 도시형의 산림식생은 국수나무(99.9), 애기나리(96.9), 팥배나무(95.9), 덜꿩나무(92.0), 때죽나무(88.0), 담쟁이덩굴(84.0), 청가시덩굴(80.0), 참회나무(76.0), 벚나무류(72.0), 가는잎족제비고사리(70.4), 당단풍나무(61.5), 조록싸리(56.0), 물오리나무(52.0), 진달래(52.0), 작살나무(50.9), 졸참나무(48.0), 개옻나무(40.0), 왕머루(31.7) 순으로 18종이, 산악형의 산림식생은 피나무(100.0), 까치박달(100.0), 실새풀(99.4), 철쭉(99.0), 미역줄나무(96.0), 단풍취(94.9), 개갈퀴(92.0), 금강애기나리(92.0), 노루귀(92.0), 오미자(92.0), 대사초(88.4), 노루오줌(88.0), 큰개별꽃(88.0), 잣나무(87.3), 개고사리(84.3), 여로(80.0), 참나물(76.0), 참취(76.0), 함박꽃나무(76.0), 고광나무(72.0)순으로 20종이 유의한 지표종으로 나타났다(p<0.01).
Figure 2.
Indicator value of major species in Fagaceae forest. All species with an indicator value > 30% are mentioned for each site hierarchy where they are founded.
Download Original Figure
4. CCA 분석
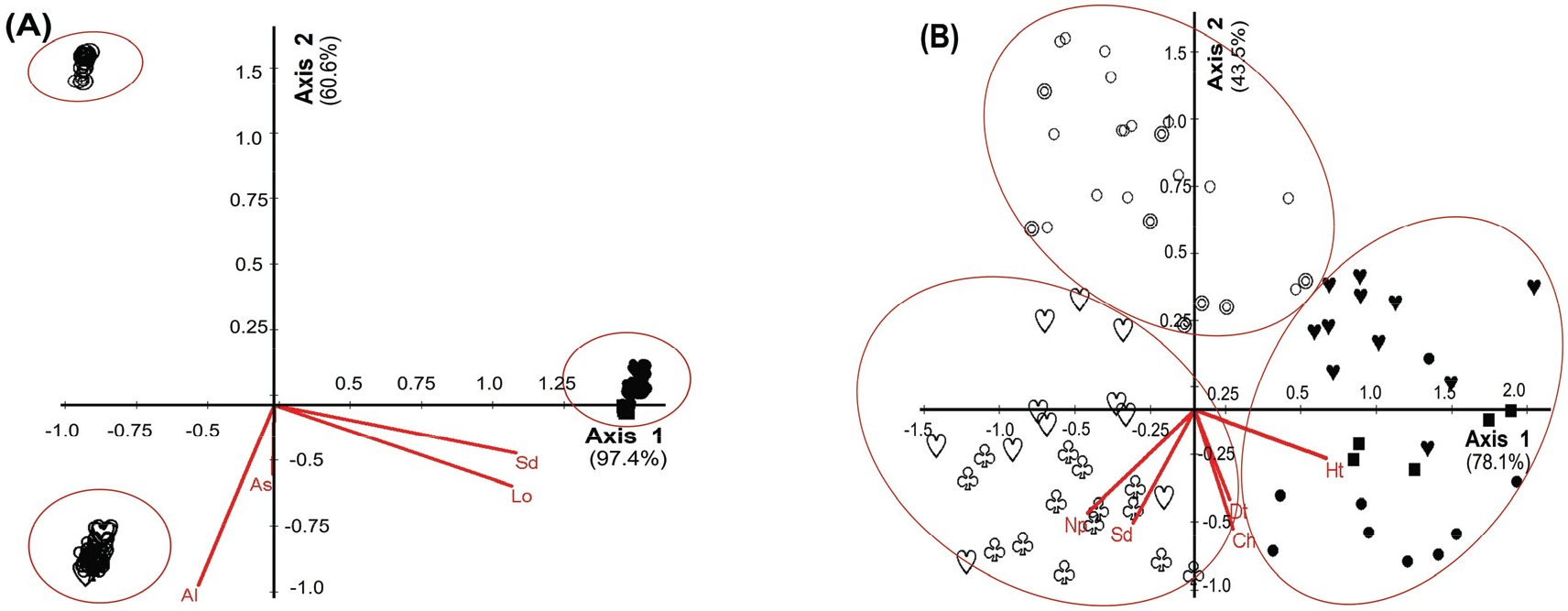
본 연구에서 참나무과 조사지의 식생단위의 종과 환경과의 상관관계를 정량적인 방법으로 알아보기 위해 정준대응분석 방법을 이용하여 파악해보았다(Figure 3, Table 3, 4). 그 결과 무생물적 환경인자(Abiotic Environmental)는 Axis 1 97.4%, Axis 2 60.6%로 나타내었으며, 식생단위 간 상관관계를 살펴보면 도시형(Vegetation Unit 1, 2)은 경사 및 경도와 부의 상관관계를 가지는 것으로 나타났고, 산악형(Vegetation Unit 3, 4)은 해발고도와 정의 상관관계를 가지는 것으로 나타났으며 도서형(Vegetation Unit 5, 6, 7)은 경사 및 경도와 정의 상관관계를 가지는 것으로 나타났다. 생물적 환경인자(Biotic Environmental)는 Axis 1 78.1%, Axis 2 43.5%로 나타났다. 식생단위 간 상관관계는 도시형(Vegetation Unit 1, 2)은 초본층 식피율과 교목층 DBH와 부의 상관관계를 가지는 것으로 나타났고, 오대산형(Vegetation Unit 3, 4)은 출현종수, 종다양도와 정의 상관관계를 가지는 것으로 나타났으며 울릉도형(Vegetation Unit 5, 6, 7)은 교목층 수고와 정의 상관관계를 가지는 것으로 나타났다. 따라서 식생유형과 입지환경인자와의 상관관계 결과 세 지역형을 나누는 환경인자는 해발고도이며(p<0.05), 비생물적 환경인자 중 해발고도가 가장 중요한 인자라는 기존의 연구결과와 일치하였다(Yun and Hong, 2000; Lee et al., 2001; Song et al., 2019).
Figure 3.
Canonical correspondence analysis(CCA) ordination diagram showing 7 vegetation units and environmental variables(A; abiotic environmental - Altitude: Al, Slope degree: Sd, Longitude: Lo, Aspect: As, B; biotic environmental - Height of tree layer: Ht, DBH of tree layer: Dt, Coverage of herb layer: Ch, Species diversity: Sd, No. of present species: Np) against axis 1 and axis 2(Cutoff R2; A: 0.200, B: 0.430, ○: VU1, ◎: VU2, ♧: VU3, ♡: VU4, ●: VU5, ■: VU6, ♥: VU7).
Download Original Figure
Table 3.
Summary Statistic of canonical correspondence analysis(CCA) with environment.
|
|
Abiotic Environment |
Biotic Environment |
|
Axis1 |
Axis2 |
Axis3 |
Axis1 |
Axis2 |
Axis3 |
|
Eigenvalue |
0.974 |
0.606 |
0.139 |
0.781 |
0.435 |
0.162 |
|
Variance in species data |
% of variance explained |
15.8 |
9.8 |
2.2 |
12.7 |
7.0 |
2.6 |
|
Cumulative % explained |
15.8 |
25.6 |
27.9 |
12.7 |
19.7 |
22.4 |
|
Pearson Correlation, Species-Environmental*
|
0.999 |
0.987 |
0.701 |
0.910 |
0.863 |
0.756 |
|
Kendall(Rank) Corr., Species-Environmental |
0.576 |
0.720 |
0.530 |
0.714 |
0.661 |
0.527 |
Download Excel Table
Table 4.
Canonical coefficients and inter-set correlations between ordination axes and environment factors.
|
|
Variables |
Canonical coefficients |
Correlation coefficients |
|
Axis1 |
Axis2 |
Axis3 |
Axis1 |
Axis2 |
Axis3 |
|
Abiotic Environment |
Altitude |
−0.439 |
−0.834*
|
0.092 |
−0.288 |
−0.956 |
−0.039 |
|
Aspect |
−0.003 |
0.011 |
0.018 |
−0.006 |
−0.372 |
0.132 |
|
Slope degree |
−0.028 |
−0.103 |
0.088 |
0.917 |
−0.258 |
0.026 |
|
Topography |
0.005 |
0.035 |
−0.377 |
0.173 |
−0.238 |
−0.663 |
|
Bare rock |
0.002 |
0.013 |
−0.471 |
0.270 |
−0.200 |
−0.767 |
|
Longitude |
0.988 |
−0.172 |
0.154 |
0.902 |
−0.431 |
−0.011 |
|
Biotic Environment |
Height of tree layer |
0.544 |
−0.272 |
0.086 |
0.805 |
−0.345 |
0.421 |
|
DBH of tree layer |
−0.045 |
0.039 |
0.564 |
0.212 |
−0.640 |
0.676 |
|
Coverage of herb layer |
0.324 |
−0.391 |
−0.548 |
0.233 |
−0.858 |
−0.375 |
|
Species diversity |
0.149 |
−0.168 |
0.026 |
−0.377 |
−0.815 |
0.087 |
|
No. of present species |
−0.752 |
−0.364 |
0.078 |
−0.654 |
−0.743 |
0.120 |
Download Excel Table
고 찰
낙엽참나무류는 위도구간 37°와 35.5°에서 출현빈도가 가장 높게 나타나며, 특히 신갈나무는 위도 37° 부근에서 가장 높은 출현빈도를 유지한다고 알려져 있다(Kim and Kim, 2017). 본 논문은 온대중부권역의 대표 극상 수종으로 거론되는 참나무과 낙엽활엽수종 우점림을 대상으로 동위도상 지리적 위치 및 생태특성이 이질적인 지역을 선정하여 환경에 따른 군락의 구조, 천이 경향 등을 파악하고자 시행하였다. 산림식생에 대한 군집구조 비교를 위해 생활형이 다른 남산, 오대산, 울릉도에서 조사된 75개소의 산림식생조사 자료를 바탕으로 Z-M학파의 식물사회학적 방법에 의한 식생유형 분류를 수행한 결과 2개 군락, 4개 군, 6개 소군으로 총 7개 유형이 구분되었다. 7개로 나뉜 식생유형은 다시 식생단위 1, 2가 도시형, 식생단위 3, 4가 산악형, 식생단위 5, 6, 7이 도서형으로 구분되었다.
식생 유형별 종구성과 환경인자과의 상관관계를 정량적인 방법으로 파악해보고자 CCA를 분석해본 결과, 세 지역형의 주요 종구성을 나누는 요인은 해발고도로 나타났으며, 이는 식생분포의 가장 큰 요인은 해발고도라는 기존 연구결과와 일치하였다(Yun and Hong, 2000; Lee et al., 2001; Song et al., 2019).
또한 각 유형별 생태적 천이특성을 규명하고자 중요치 분석, 지표종 분석을 시행한 결과 도시형과 도서형은 우점종인 참나무과 수종으로 유지될 것으로 보인 반면, 산악형의 경우 피나무, 까치박달 등 활엽수종이 우점하는 군락으로의 천이가 이루어질 것으로 판단되었다.
낙엽참나무류는 우리나라의 대표 극상 수종으로 거론되는 종으로서, 이들 참나무림의 발달, 확장, 쇠퇴에 대해 보다 구체적 검증을 위해서는 시간적으로 개체군 구조 변화에 대한 연구가 필요하다. 또한 지속적 이용을 위한 생태적 관리를 위해서는 각 식생단위의 종구성적 특성, 해발고도, 경도, 지형 등에 따른 개체군 및 군락수준의 특성을 고려한 관리방안이 필요할 것으로 사료된다.