Article
경북지방 아까시나무 조림지의 식생유형과 생태적 특성
송재순
1, 김학윤
1, 김준수
2, 오승환
3, 조현제
2,*
Vegetation Classification and Ecological Characteristics of Black Locust (Robinia pseudoacacia L.) Plantations in Gyeongbuk Province, Korea
Jae-Soon Song
1, Hak-Yun Kim
1, Jun-Soo Kim
2, Seung-Hwan Oh
3, Hyun-Je Cho
2,*
1Department of Global Environment, Keimyung University, Daegu 42601, Korea
2Nature and Forest Research Institute, Daegu 41476, Korea
3School of Forest Science & Landscape Architecture, Kyungpook National University, Daegu 41566, Korea
© Copyright 2023 Korean Society of Forest Science. This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received: Dec 02, 2022; Revised: Dec 26, 2022; Accepted: Dec 29, 2022
Published Online: Mar 31, 2023
요약
본 연구는 경북 일대 산지 아까시나무림의 자연성 회복을 위한 생태적 관리에 필요한 기초정보 제공을 위하여 총 200개 임분에서 수집된 식생자료를 바탕으로 이원지표종분석법(TWINSPAN)으로 식생유형을 분류하고, DCA 분석으로 환경구배에 따른 그 공간배열 상태를 파악함과 아울러 식생유형별 군락적합도(Φ) 판정으로 결정된 진단종을 기준으로 군락조성요약표를 작성하였다. 식생유형은 신갈나무-둥굴레형, 밤나무-청미래덩굴형, 철쭉-인동덩굴형, 찔레꽃-쑥형, 감태나무-댕댕이덩굴형, 참느릅나무-팽나무형, 그리고 귀룽나무-푼지나무형 등 모두 7개 유형으로 분류되었으며, 대개 해발고도, 수분 체계, 천이 단계 및 교란 체제 등 복합적 요인의 차이를 반영하고 있었다. 구성종의 평균상대중요치는 아까시나무(39.7)가 단연 높게 나타났지만, 굴참나무, 졸참나무, 신갈나무, 상수리나무, 갈참나무 등 참나무류가 상위중요치 구성종으로 나타나고 있어 잠재천이경향을 가늠할 수 있었다. 또한, 식생유형별 구성종의 총피도, 생활형 조성, 종다양성 지수, 그리고 지표종 등도 비교하였다.
Abstract
This study was established to provide basic information necessary for ecological management to restore the naturalness of black locust (Robinia pseudoacacia L.) plantations located in the mountains of Gyeongbuk, Korea. Using vegetation data collected from 200 black locust stands, vegetation types were classified using the TWINSPAN method, the spatial arrangement status according to the environmental gradient was identified through DCA analysis, and a synoptic table of communities was prepared based on the diagnostic species determined by determining community fidelity (Φ) for each vegetation type. The vegetation types were classified into seven types, namely, Quercus mongolica-Polygonatum odoratum var. pluriflorum type, Castanea crenata-Smilax china type, Clematis apiifolia-Lonicera japonica type, Rosa multiflora-Artemisia indica type, Quercus variabilis-Lindera glauca type, Ulmus parvifolia-Celtis sinensis type, and Prunus padus-Celastrus flagellaris type. These types usually reflected differences in complex factors such as altitude, moisture regime, successional stage, and disturbance regime. The mean relative importance value of the constituent species was highest for black locust (39.7), but oaks such as Quercus variabilis, Q. serrata, Q. mongolica, Q. acutissima, and Q. aliena were also identified as important constituent species with high relative importance values, indicating their potential for successional trends. In addition, the total percent cover of constituent species by vegetation type, life form composition, species diversity index, and indicator species were compared.
Keywords: black locust; TWINSPAN; DCA; vegetation types; diagnostic species
서 론
북미 원산 콩과 교목성 수종인 아까시나무(Robinia pseudoacacia L.)는 녹화, 신탄재, 용재, 밀원, 토양안정화, 산림재생, 생물치료, 탄소격리 등 다목적 활용성으로 활엽수에 한정하여 보면 전 세계적으로 유카리속(Eucalyptus spp.)나 포플러속(Populus spp.) 다음으로 많이 심어진 수종이다(Tamura et al., 2007; Nicolescu et al., 2020). 그러나 최근 들어 세계 여러 나라에서 왕성한 번식력을 가진 아까시나무가 산복, 계류, 하천, 해안, 휴‧폐경지, 저지대 등 다양한 입지에 침입 및 정착하여 다른 식물의 성장을 방해하거나 고유 식생의 구조 및 조성 그리고 동태를 변화시켜 생태계의 다양성 및 자연성 저하를 유발하는 등 부정적인 생물다양성 영향과 사회‧경제적 이익 사이의 논란이 심화하면서 그 관리가 현안으로 대두되고 있다(Tamura et al., 2007; Vilà et al., 2009; Sádlo et al., 2017). 이에 우리나라를 비롯하여 아까시나무가 널리 식재되어 있는 유럽 및 일본에서는 아까시나무림의 생태적 관리와 자연성 회복 방안을 모색하기 위하여 종조성적 식생 분류, 임분 구조 및 천이 경향 분석 등 다양한 연구를 수행하여 오고 있다. 식생 분류 연구는 식물군락을 유형화하는 식물사회학의 한 분야인 군락분류학(syntaxonomy) 측면에서 수행되어 왔는데, 주요 연구를 보면 유럽에서는 Vítková and Kolbek (2010)이 중부유럽 아까시나무림의 식생 분류를 통하여 “1개 군강(class), 2개 군목(order), 3개 군단(alliance), 6개 군집(association)”의 식생단위 체계를 보고한 바 있으며, 우리나라에서는 Cho and Kim(2005)이 우리나라 아까시나무림을 아까시나무-닭의장풀군집(전형아군집, 떡갈나무아군집, 자귀나무아군집, 소나무아군집, 굴참나무아군집, 가죽나무아군집), 아까시나무-갈대군락(전형하위군락, 띠하위군락)의 “1개 군집, 1개 군락, 6개 아군집, 2개 하위군락 등” 모두 8개 유형으로 분류하였고, 대구 및 경북일대 특정 산지의 식생 분류 연구에서 아까시나무-닭의장풀군락(Song and Kim, 1993; Choung et al., 2000)과 아까시나무-찔레꽃군락(Jegal and Kim, 2003)이 보고된 바 있다. 이외에 아까시나무림의 관리 및 자연성 복원 모델 도출을 위하여 군집 특성 및 천이 경향 분석 측면에서도 다양한 연구가 수행되어 왔는데(Lee and Han, 1998; Yun et al., 1999; Yun et al., 2001; Kang and Bang, 2001; Lee et al., 2004; Cho et al., 2009; Kim and Oh, 2018), 이 중 Lee et al.(2004)은 교란 체제와 아까시나무 발생 간에 상관관계가 있다고 보고한 바 있다.
본 연구는 상기의 점을 감안하여 우리나라에서 아까시나무 식재 면적이 가장 넓고 교란 체제(disturbance regime)도 다양하여 생태적 또는 생물다양성 측면에서 부정적 영향 심화가 우려되고 있는 대구 및 경북 일대(이하 경북) 산지 아까시나무림의 식생 분류와 군락 생태 특성을 파악하여 향후 그 자연성 회복을 위한 생태적 관리 계획 수립에 필요한 기초자료를 제공하고자 한다.
재료 및 방법
1. 조사지 개황
본 조사지는 지리적으로는 북위 35도 34분에서 37도 08분, 동경 127도 47분에서 129도 34분 사이에 위치하고, 태백산맥과 소백산맥의 크고 작은 산으로 둘러싸인 내륙지방과 해안의 영향을 받는 동해안지역, 낙동강의 영향을 받는 중앙 저지대지역으로 크게 구분되어 있다. 기후적으로 내륙성기후와 해양성 기후가 중첩되어 있으며 최근 30년간(1991|2020) 기후자료에 따르면 연평균 기온은 13.6°C, 연평균 강수량은 평균 1,123.4 mm로 전형적인 대륙성 기후환경을 나타내고 여름과 겨울의 기온 차가 매우 큰 것이 특징이다(Korea Meterological Administration, 2022). 지질적으로는 소백산지괴, 경상분지, 포항분지, 대구분지 등으로 구분되어 구성되어 있으며, 동으로 갈수록 이들의 지질연대가 새로우며, 이들 각 지층의 지반 위에 제4기층이 퇴적된 지질구조를 갖고 편마암류, 화강암류, 화산암류 등이 특징적으로 나타난다(Choi et al., 2013). 식물구계학적으로는 중부아구와 남부아구에 속한다(Lee and Yim, 2002). 대구 및 경북 일대 아까시나무림의 총면적은 9,822 ha이며 이는 우리나라 아까시나무림 전체 면적의 약 41.8%이다(Korea Forest Service, 2022).
2. 조사 및 분석 방법
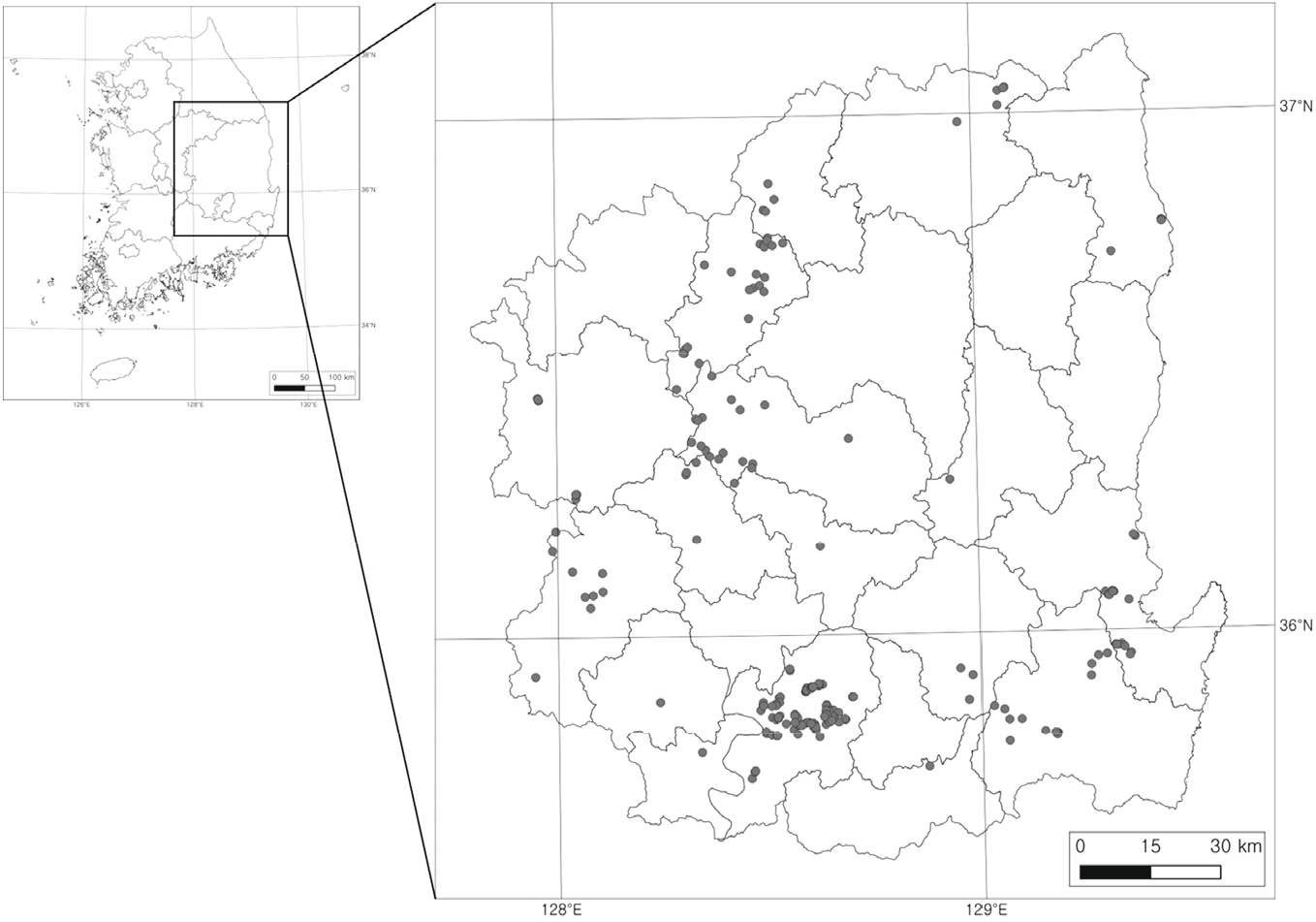
야외 조사는 2018년 6월에서 2021년 9월까지 경북 일대 산지에 분포하는 아까시나무림을 대상으로 그 공간분포, 군락상관 그리고 입지환경 등을 고려하여 총 200개 지소의 임분을 선정하고(Figure 1), 대상 임분의 상관이 가장 전형적인 곳에 10 m×10 m 또는 20 m×20 m 크기의 정방형 방형구를 설치한 후, Z-M학파의 식물사회학적 방법(Braun-Blanquet, 1965)으로 식생조사를 수행하고 그 입지환경 특성(해발, 사면경사, 사면방향, 암석노출도 등)을 파악하였다. 수집된 식생조사 자료는 MS-Excel 2016을 이용하여 소표(raw table)화 한 후, 그 소표를 근거로 하여 먼저 TWINSPAN 분석법(Hill, 1979)으로 식생유형을 분류하고 DCA 분석법(Hill and Gauch, 1980)으로 환경구배에 따른 공간배열 상태를 파악한 후, 최종적으로 식생유형별 구성종의 군락적합도(fideltiy) 판정으로 결정된 진단종(diagnostic species)을 기준(Tichý and Chytrý, 2006)으로 군락조성요약표(Synoptic table)를 작성하였다. 군락조성요약표상 식생유형 명칭은 진단종의 파이 계수(Phi coefficient, Φ), 출현빈도 및 우점도를 고려하여 상위 1|2종을 연명하여 부여하였다. 그리고 식생유형 간 생태적 특성을 구성종의 층위별 총피도(total percent cover), 중요치(importance value: IV), 평균상대중요치(mean relative importance value: MIV), 생활형 조성(life form composition), 종다양성 지수(species diversity indices), 그리고 지표종(indicator species) 등으로 비교하였다. 총피도는 식생 층위 간 피도 총합(Caratti, 2006), 중요치(IV)는 Curtis and McIntosh(1951)의 평가 기준, 평균상대중요치(MIV)는 중요치(IV)에 식생층위 간 개체 크기를 고려하여 층위별로 가중치를 부여한(교목층 IV×3+아교목층 IV×2+관목층 IV×1+초본층 IV× 0.5)/6.5의 식을 사용하여 구하였다(Kim et al., 2022). 생활형 조성은 Raunkiaer(1934)의 휴면형(dormancy form)과 Numata(1947)의 산포기관형(disseminule forms), 지하기관형(radicoid forms), 그리고 생육형(growth forms) 구분 기준을 적용하였다. 종다양성 지수는 종풍부도(species richness index: S), 종다양도(Shannon diversity index; H′), 종균재도(Pielou evenness index; J′), 그리고 종우점도(Simpson dominance index; D) 등을 적용하였다(Shanon and Weaver, 1949; Brower and Zar, 1977; Whittaker, 1972). 지표종 선정은 Dufrêne and Legendre(1997)의 IndVal (indicator value)방법을 적용하였다. 식생유형 분류와 군락적합도 분석에는 PC-ORD version 7 (McCune and Mefford, 2016) 그리고 기타 분석에는 Past version 4.06 (Hammer, 2021)을 사용하였다. 유관속식물의 동정은 원색대한식물도감(Lee, 2003)을 참조하였으며, 그 학명과 국명은 국가표준식물목록(Korea National Arboretum, 2022) 기준을 적용하였다.
결과 및 고찰
1. 식생유형
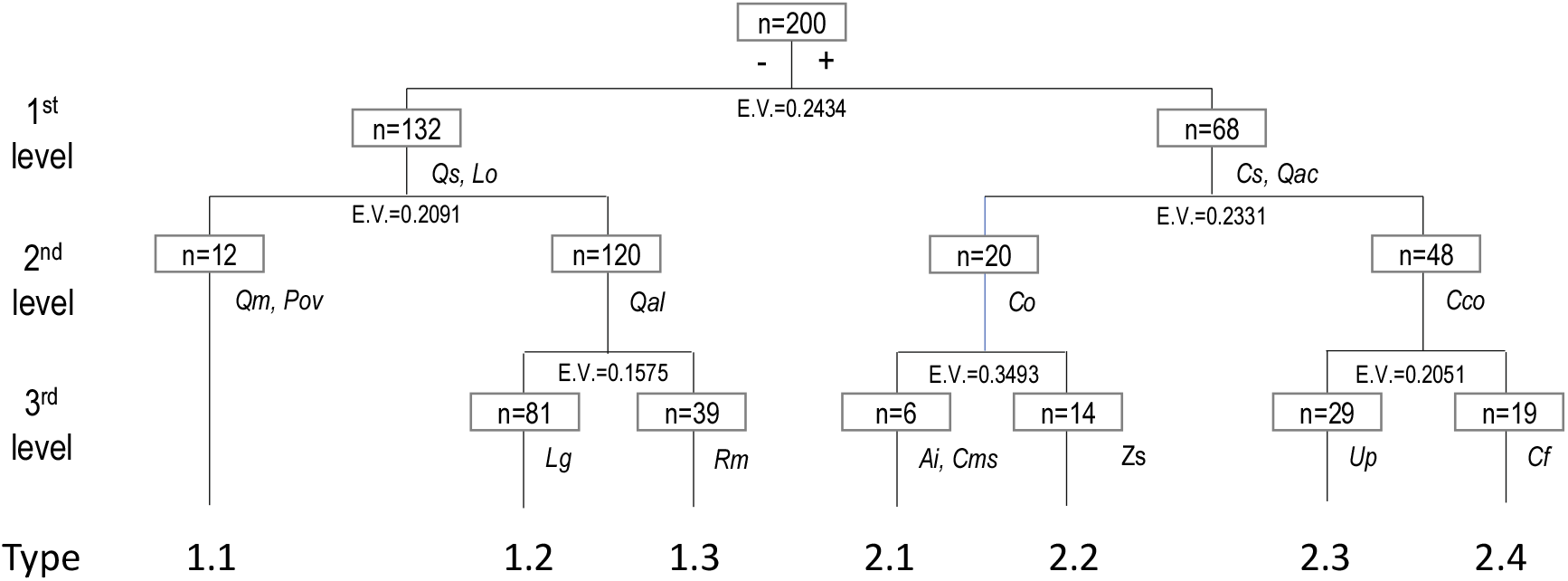
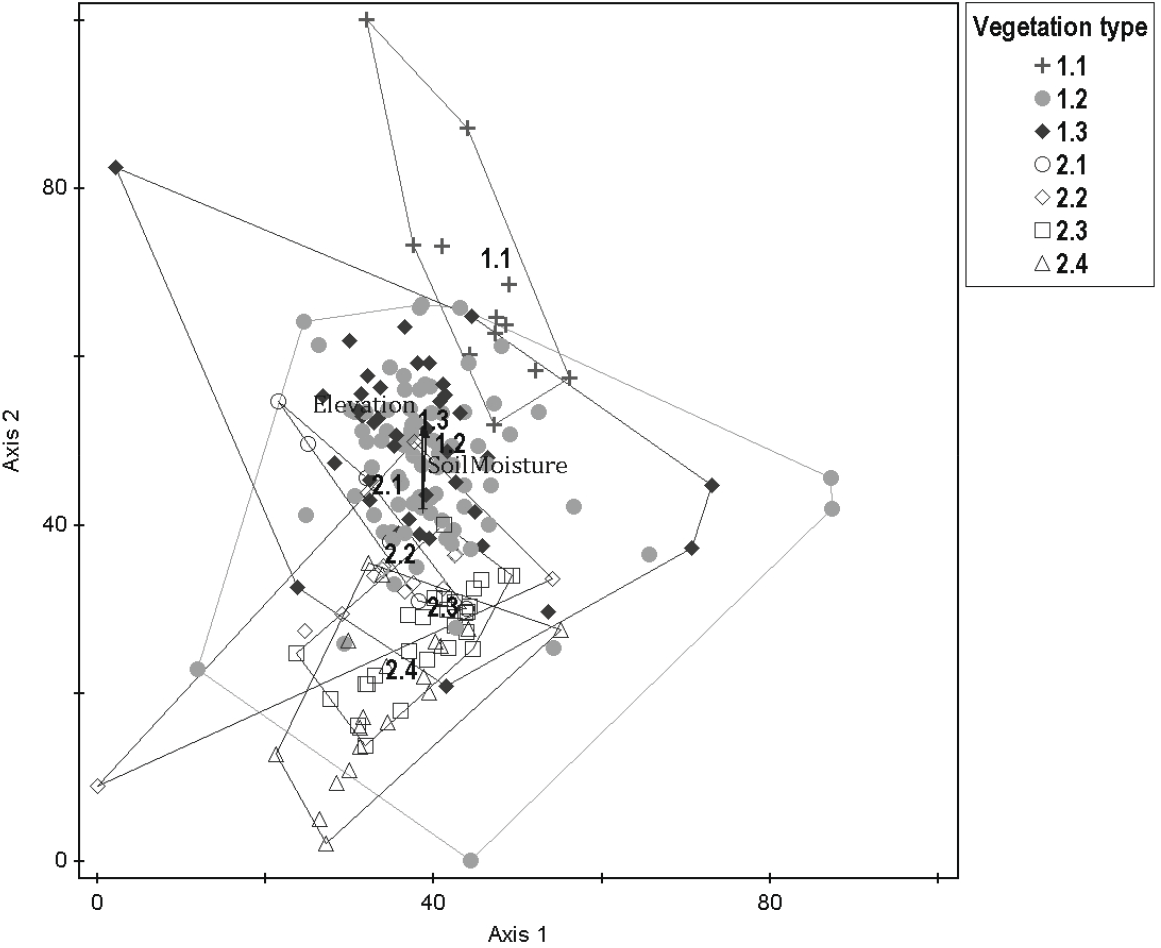
경북 일대 산지 아까시나무림 200개 임분에서 수집한 식생조사 자료를 이원지표종분석법(TWINSPAN)으로 그룹화한 결과, 모두 7개 식생유형이 구분되었다(Figure 2). 제1단계(Level 1)에서는 졸참나무(Qs)와 생강나무(Lo)가 특징짓는 왼쪽 그룹(−)과 팽나무(Cs)와 상수리나무(Qac)가 특징짓는 오른쪽 그룹(+)으로 구분되었으며, 제2단계(Level 2)에서는 왼쪽 그룹은 신갈나무(Qm)와 둥굴레(Pov)가 특징짓는 그룹과 갈참나무(Qal)가 특징짓는 그룹으로, 오른쪽 그룹은 노박덩굴(Co)이 특징짓는 그룹과 닭의장풀(Cco)이 특징짓는 그룹으로 다시 구분되었다. 제3단계(Level 3)에서는 왼쪽 그룹의 갈참나무(Qal)가 특징짓는 그룹은 감태나무(Lg)가 특징짓는 그룹과 찔레꽃(Rm)이 특징짓는 그룹으로, 오른쪽 그룹의 노박덩굴(Co)이 특징짓는 그룹은 다시 쑥(Ai)과 애기똥풀(Cms)이 특징짓는 그룹과 산초나무(Zs)가 특징짓는 그룹 그리고 닭의장풀(Cco)이 특징짓는 그룹은 참느릅나무(Up)가 특징짓는 그룹과 푼지나무(Cf)가 특징짓는 그룹으로 최종 구분되었다. 또한, DCA 기법(Hill, 1979)을 이용하여 TWINSPAN에 의해 분류된 식생유형 간 조사구의 공간배열 상태를 비교한 결과(Figure 3), DCA 제1축과 제2축의 eigenvalue가 각각 0.402, 0.344로서 3개 축 전체 합 1.033의 72.1%에 해당하여 총분산(total variance)에 대한 집중률이 높았고, Y축의 상단에서 아래로 1.1형에서 2.4형까지 7개 식생유형이 배열되어 있었는데 대개 해발고와 토양 수분체계 차이를 반영하고 있었다.
Figure 2.
Simplified TWINSPAN classification tree (dendrogram) of Robinia pseudoacacia plantations in the study area. 344 plant species × 200 relevés matrix. For each level of division number of group, numbers of relevés (in boxes) are shown. Type 1.1 to 2.4 were defined as main vegetation types of Robinia pseudoacacia plantations and each cluster is characterized by the species that are listed in its column. For full names of species (Qs, Lo, Cs, etc.) and types 1.1 to 2.4 see Table 1.
Download Original Figure
Table 1.
Synoptic table of Black Locust (Robinia pseudoacaia L.) plantations with percentage constancy and median cover of species. Species are sorted by their life form (LF; trees = 3, shrubs = 2, herbs = 1) and within them by decreasing constancy. Dark grey and light grey indicate Φ > 0.40 and Φ > 0.20, respectively. Only species with a constancy >15% are listed; different thresholds are due to differences in species richness among groups. Transgressive diagnostic species are those that have both their diagnostic value in more than one group and a constancy >20% in all groups for which the species is diagnostic. Forest vegetation types: 1.1 = Quercus mongolica-Polygonatum odoratum var. pluriflorum type; 1.2 = Castanea crenata-Smilax china type; 1.3 = Clematis apiifolia-Lonicera japonica type; 2.1 = Rosa multiflora-Artemisia indica type; 2.2 = Quercus variabilis-Lindera glauca type; 2.3 = Ulmus parvifolia-Celtis sinensis type; 2.4 = Prunus padus-Celastrus flagellaris type.
|
Forest vegetation type |
|
C1 |
C2 |
|
|
|
1.1 |
1.2 |
1.3 |
2.1 |
2.2 |
2.3 |
2.4 |
|
Number of relevés |
LF
|
12 |
81 |
39 |
6 |
14 |
28 |
19 |
|
Diagnostic species
|
|
|
|
|
|
|
|
|
|
Quercus mongolica (Qm)
|
3 |
92 2
|
16 +
|
23 +
|
. |
. |
. |
. |
|
Fraxinus sieboldiana
|
3 |
33 +
|
1 r
|
. |
. |
. |
. |
. |
|
Fraxinus rhynchophylla
|
3 |
25 +
|
. |
3 +
|
. |
7 +
|
. |
. |
|
Rhododendron mucronulatum
|
2 |
33 +
|
9 +
|
15 +
|
. |
. |
. |
. |
|
Lonicera praeflorens
|
2 |
25 +
|
. |
. |
. |
. |
. |
. |
|
Polygonatum odoratum var. pluriflorum (Pov)
|
1 |
75 1
|
5 +
|
10 +
|
. |
. |
4 r
|
. |
|
Carex humilis var. nana (Chv)
|
1 |
58 +
|
19 +
|
3 r
|
. |
7 r
|
. |
. |
|
Athyrium yokoscense
|
1 |
58 1
|
14 +
|
18 +
|
. |
. |
. |
. |
|
Prunus serrulata var. pubescens (Psv)
|
1 |
50 +
|
15 +
|
8 +
|
17 r
|
29 1
|
7 r
|
. |
|
Carex fililpes var. oligostachys
|
1 |
25 r
|
. |
. |
. |
. |
. |
. |
|
Syneilesis palmata
|
1 |
25 +
|
. |
. |
. |
. |
. |
. |
|
Persicaria nepalensis
|
1 |
25 +
|
. |
. |
. |
. |
. |
. |
|
Hosta longipes
|
1 |
25 r
|
. |
. |
. |
. |
. |
. |
|
Artemisia keiskeana
|
1 |
25 +
|
1 r
|
5 r
|
. |
. |
. |
. |
|
Castanea crenata (Cc)
|
3 |
17 +
|
56 1
|
36 1
|
33 +
|
. |
. |
5 r
|
|
Styrax japonicus
|
3 |
. |
30 +
|
5 +
|
. |
21 +
|
11 r
|
5 +
|
|
Smilax china (Sc)
|
2 |
25 +
|
56 +
|
31 +
|
. |
57 +
|
11 +
|
. |
|
Lonicera japonica
|
2 |
17 +
|
12 +
|
54 +
|
. |
14 r
|
11 +
|
5 r
|
|
Clematis apiifolia (Ca)
|
2 |
17 +
|
31 +
|
67 1
|
50 2
|
36 +
|
21 +
|
21 +
|
|
Quercus acutissima (Qac)
|
3 |
. |
371 |
23 +
|
83 2
|
50 1
|
36 +
|
53 1
|
|
Alnus japonica
|
3 |
. |
. |
. |
33 +
|
. |
. |
. |
|
Rubus parvifolius
|
2 |
. |
. |
3 +
|
33 +
|
14 r
|
14 r
|
5 +
|
|
Spiraea prunifolia f. simpliciflora
|
2 |
. |
5 +
|
10 +
|
33 +
|
. |
4 r
|
. |
|
Artemisia indica (Ai)
|
1 |
17 r
|
4 r
|
21 +
|
67 1
|
7 r
|
. |
. |
|
Erigeron annuus
|
1 |
. |
2 r
|
8 +
|
50 +
|
14 r
|
. |
5 r
|
|
Stellaria aquatica
|
1 |
. |
1 r
|
10 +
|
50 1
|
7 r
|
4 r
|
11 +
|
|
Aconitum jaluense
|
1 |
. |
. |
. |
33 +
|
. |
. |
. |
|
Bromus remotiflorus
|
1 |
. |
. |
. |
33 +
|
. |
. |
. |
|
Cyrtomium fortunei
|
1 |
. |
. |
. |
33 r
|
. |
. |
. |
|
Cornus walteri
|
3 |
. |
1 r
|
. |
. |
21 +
|
7 +
|
. |
|
Pyrus ussuriensis
|
3 |
. |
. |
3 +
|
. |
21 +
|
. |
. |
|
Lindera glauca
|
2 |
. |
47 +
|
3 r
|
17 +
|
86 2
|
21 +
|
11 +
|
|
Zanthoxylum schinifolium (Zs)
|
2 |
33 +
|
35 +
|
26 +
|
17 +
|
64 +
|
. |
. |
|
Securinega suffruticosa
|
2 |
. |
10 +
|
8 r
|
. |
36 +
|
a |
. |
|
Cardamine flexuosa (Cf)
|
1 |
. |
17 +
|
21 +
|
17 1
|
57 1
|
7 +
|
21 r
|
|
Boehmeria platanifolia
|
1 |
. |
1 r
|
5 +
|
. |
21 1
|
. |
5 r
|
|
Galium maximowiczii
|
1 |
. |
2 r
|
8 +
|
. |
21 r
|
. |
. |
|
Ulmus parvifolia (Up)
|
3 |
. |
. |
. |
17 +
|
21 +
|
71 +
|
16 r
|
|
Prunus padus
|
3 |
. |
2 r
|
3 +
|
. |
. |
4 +
|
45 1
|
|
Celastrus flagellaris (Gf)
|
2 |
. |
1 r
|
3 r
|
. |
14 +
|
11 r
|
63 2
|
|
Galium spurium (Gs)
|
1 |
. |
4 +
|
3 r
|
. |
. |
21 +
|
58 1
|
|
Fallopia dumetorum
|
1 |
. |
. |
3 r
|
. |
7 r
|
11 r
|
21 +
|
|
Forest vegetation type |
C1 |
C2 |
|
|
LF
|
1.1 |
1.2 |
1.3 |
2.1 |
2.2 |
2.3 |
2.4 |
|
Transgressive diagnostic species
|
|
|
|
|
|
|
|
|
|
Lindera obtusiloba (Lo)
|
2 |
92 2
|
51 1
|
31 +
|
17 +
|
7 r
|
7 +
|
5 +
|
|
Rhododendron schlippenbachii
|
2 |
25 +
|
5 +
|
23 +
|
. |
. |
. |
. |
|
Athyrium vidalii
|
1 |
8 +
|
9 +
|
28 +
|
33 +
|
. |
. |
5r |
|
Conyza canadensis
|
1 |
. |
. |
21 +
|
33 +
|
. |
. |
. |
|
Rosa multiflora (Rm)
|
2 |
8+ |
20 +
|
74 1
|
100 3 |
43 +
|
36 +
|
63 1
|
|
Celastrus orbiculatus (Go)
|
2 |
17 +
|
6 +
|
18 +
|
50 +
|
36 +
|
14 +
|
16 +
|
|
Chelidonium majus subsp. asiaticum (Cms)
|
1 |
17 +
|
5 +
|
33 +
|
67 +
|
. |
11 +
|
11 +
|
|
Paederia foetida
|
1 |
. |
6+ |
3 r
|
33 +
|
. |
. |
21+ |
|
Ailanthus altissima
|
3 |
. |
4 +
|
3 +
|
17 +
|
36 +
|
4 +
|
26 +
|
|
Diospyros lotus (Dl)
|
3 |
. |
16 +
|
5 +
|
50 +
|
50 +
|
32 +
|
21 +
|
|
Cocculus orbiculatus
|
2 |
8+ |
12 r
|
10 r
|
17 r
|
64 r
|
46 r
|
5 r
|
|
Celtis sinensis (Cs)
|
3 |
. |
7 +
|
3 +
|
. |
57 +
|
71 1
|
74 2
|
|
Quercus variabilis (Qv)
|
3 |
. |
44 1
|
21 +
|
17 +
|
79 2
|
93 2
|
37 1
|
|
Chenopodium album var. centrorubrum
|
1 |
8 r
|
. |
. |
. |
. |
32 +
|
21 r
|
|
Liriope muscari
|
1 |
. |
2 r
|
5 r
|
. |
7 r
|
25 +
|
42 +
|
|
Commelina communis (Cco)
|
1 |
33 +
|
16 +
|
28 +
|
33 +
|
7 r
|
57 +
|
68 r
|
|
Constant & companion species
|
|
|
|
|
|
|
|
|
|
Robinia pseudoacacia (Rp)
|
3 |
100 4
|
100 5
|
100 5
|
100 5
|
100 5
|
100 5
|
100 5
|
|
Oplismenus undulatifolius (Ou)
|
1 |
50 1
|
85 2
|
67 1
|
17 +
|
71 +
|
79 +
|
74 +
|
|
Quercus serrata (Qs)
|
3 |
67 2
|
65 2
|
62 2
|
33 +
|
7 r
|
25 +
|
21 +
|
|
Ligustrum obtusifolium (Lob)
|
2 |
17 +
|
35 +
|
54 1
|
17 1
|
29 +
|
39 +
|
32 +
|
|
Quercus aliena (Qal)
|
3 |
17 +
|
46 1
|
38 1
|
17 +
|
29 +
|
. |
16 +
|
Download Excel Table
Figure 3.
DCA ordination diagram with axis 1 and 2 of relevés of Robinia pseudoacacia plantations in the study area. For full names of types 1.1 to 2.4 see Table 1.
Download Original Figure
Table 1은 TWINSPAN으로 구분된 7개 식생유형별 전체 구성종에 대한 군락적합도(fidelity) 평가로 결정된 진단종(diagnostic species)과 그들의 수도(Φ 계수, 출현 빈도, 우점도)를 중심으로 단위 식생유형의 명칭이 부여된(Type 1.1: 신갈나무-둥굴레형, Type 1.2: 밤나무-청미래덩굴형, Type 1.3: 철쭉-인동덩굴형, Type 2.1: 찔레꽃_쑥형, Type 2.2: 감태나무-댕댕이덩굴형, Type 2.3: 참느릅나무-팽나무형, 그리고 Type 2.4: 귀룽나무-푼지나무형) 경북 일대 산지 아까시나무림의 군락조성요약표(synoptic table)이다. 군락조성요약표상 각 구성종에 표기된 아라비아숫자와 윗첨자(5, 4, 3, 2, 1, +, r)는 각각 해당종의 단위 식생유형내 출현 빈도와 우점도(Braun-Blanquet, 1956)를 나타내며 그리고 어두운 회색 음영과 밝은 회색 음영은 각각 군락적합도 척도인 파이 계수(Φ)가 >0.40, >0.20인 진단종(daignostic species)을 나타낸다. 신갈나무-둥굴레형(Quercus mongolica-Polygonatum odoratum var. pluriflorum type)은 지리적으로 영주, 상주, 예천, 봉화 등 경북 북부 및 서북부 산지의 해발고도 200 m 이상의 사면부 적윤지, 밤나무-청미래덩굴형(Castanea crenata-Smilax china type)은 지리적으로 대구, 경주, 예천, 포항, 군위 등의 해발고도 100|200 m 내외의 북향 또는 북서향의 사면 중ㆍ하부 급경사 약건지, 사위질빵-인동덩굴형(Clematis apiifolia-Lonicera japonica type)은 대구를 제외한 경북 일대의 해발고도 50|350 m 내외인 북향의 사면 하부 적윤지 또는 약건지, 찔레꽃-쑥형(Rosa multiflora-Artemisia indica type)은 지리적으로 김천, 영천, 예천, 군위 등의 해발고도 200 m 이하의 북향 또는 남향인 사면 하부 완경사 적윤지, 굴참나무-감태나무형(Quercus variabilis-Lindera glauca type)은 지리적으로 대구의 해발고도 100|150 m 내외의 동향의 사면 중부 건성지, 참느릅나무-팽나무형(Ulmus parvifolia-Celtis sinensis type)은 지리적으로 대구의 해발고도 50|150 m 내외의 북향 사면을 제외한 모든 사면 중부 건성지, 그리고 귀룽나무-푼지나무형(Prunus padus-Celastrus flagellaris type)은 지리적으로 대구와 인접 경북 칠곡의 해발고도 100 m 이하의 남향사면을 제외한 모든 사면 중부 완경사 약건지에 주로 분포하였다.
2. 식생구조
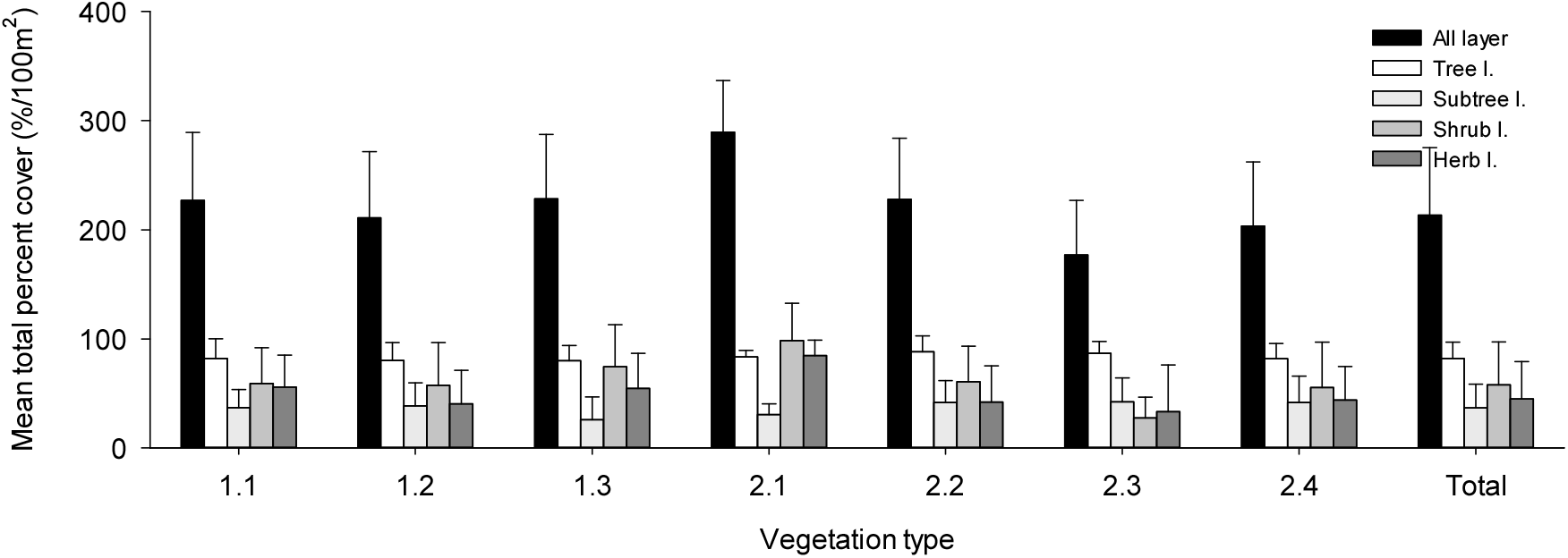
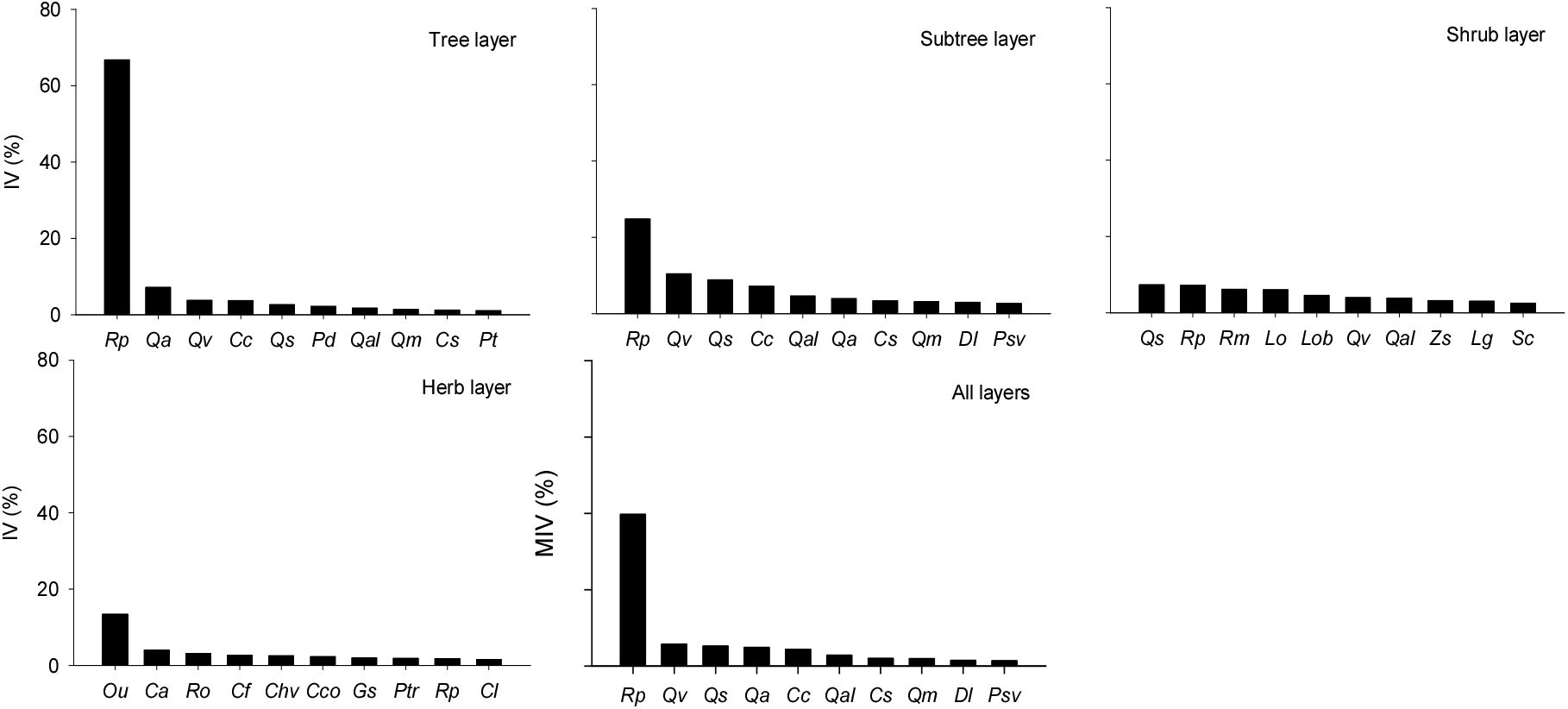
경북 일대 산지 아까시나무림의 식생구조 특성을 전체 임분 및 식생유형 간 구성종의 단위 면적당(/100 m2) 총피도, 중요치, 생활형 조성 그리고 종다양성 지수 등으로 파악하였다. 총피도는 성엽기에 단위 면적당 구성종의 총생체량(total biomass)을 간접적으로 가늠하는 기준으로 식생층위 간 구성종의 피도 중복성을 모두 반영하여 나타내므로 그 값이 100%를 초과하는 경우도 있다(Caratti, 2006). 총피도(Figure 4)는 식생유형 간 다소 차이가 있지만, 전체 평균 213±82%이었으며, 식생층위 간에는 교목층(82%)이 가장 높았고, 다음으로 관목층(58%), 초본층(45%), 그리고 아교목층(37%) 순이었으며 교목층이 관목층과 초본층에 비해 거의 두 배정도 높은 경향이었다. 식생유형 간에는 사면하부 완경사 적윤지를 특징짓는 찔레꽃-쑥형(Type 2.1)이 290±83%로 가장 높게 나타났고, 해발 100 m이하 저지대 사면중부 건성지를 특징짓는 참느릅나무-팽나무형(Type 2.4)이 177±87%로 가장 낮게 나타났는데, 주로 관목층과 초본층의 발달 정도 차이에 따른 것으로 판단되었다. 구성종의 생태적 우세정도를 가늠하는 중요치(Figure 5)는 전층 기준 평균상대중요치(MIV)에서는 아까시나무(39.7)가 단연 높게 나타났고, 다음으로 굴참나무(5.6), 졸참나무(5.2), 상수리나무(4.9), 밤나무(4.3), 갈참나무(2.8), 팽나무(1.9), 신갈나무(1.9), 고욤나무(1.4), 잔털벚나무(1.3) 순으로 나타났으며, 중요치 상위 10종 내에 잠재자연식생인 졸참나무와 신갈나무를 비롯한 5종의 참나무류가 자리하고 있어 경북 일대 산지 아까시나무림의 잠재 천이경향을 가늠할 수 있었다. 식생층위 간 중요치(IV)에서는 교목층은 우점종인 아까시나무(66.8)가 단연 높게 나타났고, 다음으로 상수리나무(7.1), 굴참나무(3.7), 밤나무(3.7), 졸참나무(2.6) 순, 아교목층은 아까시나무(24.8)가 가장 높게 나타났고, 다음으로 굴참나무(10.4), 졸참나무(8.9), 밤나무(7.1), 갈참나무(4.6) 순, 관목층은 졸참나무(7.3)와 아까시나무(7.2)가 가장 높게 나타났고, 다음으로 찔레꽃(6.1), 생강나무(6.0), 쥐똥나무(4.6) 등 관목성 식물 순, 그리고 초본층은 동물산포형 식물인 주름조개풀(13.3)이 단연 높게 나타났고, 다음으로 사위질빵(4.0), 줄딸기(3.1), 황새냉이(2.7), 그늘사초류(2.5), 닭의장풀(2.3), 갈퀴덩굴(2.0), 담쟁이덩굴(1.9) 순이었다.
Figure 4.
Mean total percent cover (/100 m2) by vegetation layers of constituent species for the seven vegetation types classified in the study area.
Download Original Figure
Figure 5.
Importance value (MIV, RIV) of the top 10 most important species by vegetation layers in Robinia pseudoacacia plantations. Among the abbreviations of species names (Rp, Qv, Qs, … etc.), Ro and Ptr are Rubus oldhamii and Parthenocissus tricuspidata, respectively, and the rest are presented in Table 1.
Download Original Figure
구성종의 생활형 조성 체계를 평균상대중요치(MIV)와 종수 구성비(PS) 기준에서 분석한 결과(Table 2), 식생유형간 다소 차이가 있지만, 전체적으로 보면 평균상대중요치 기준에서는 휴면형은 대형지상식물(MM), 지하기관형은 단립식물(R5), 산포기관형은 중력산포형(D4), 그리고 생육형은 직립형(e)이 중심 유형인 ‘MM-R5-D1-e’체계, 종수 구성비 기준에서는 휴면형은 반지중식물(H), 지하기관형은 단립식물(R5), 산포기관형은 중력산포형(D4), 그리고 생육형은 직립형(e)이 중심 유형인 ‘H-R5-D4-e’의 체계로 나타났다.
Table 2.
Mean relative importance values (MIV) and percentage of species (PS) for each component of the four life-forms (dormancy, radicoid, disseminule, growth) of all vascular plants that occurred for the seven vegetation types classified in the study area. MM: megaphanerophytes, M: microphanerophtes, N: nanophanerophtes, H: hemicryptophytes, Odo: other dormancy forms, R1: widest extent of rhizomatous growth, R2: moderate extent of rhizomatous growth, R3: narrowest extent of rhizomatous growth, R5: non-clonal growth (monophyte), R1-2 or R2-3: plant with rhizomatous mutation of R1 and R2 or R2 and R3, Ora: other radicoid forms, D1: disseminated widely by wind or water, D2: disseminated attaching with or eaten by animals and man, D4: having no special modification for dissemination, D2,4: plant with D2 and D4, b: branched form, e: erect form, p: procumbent form, ps: pseudo-rosette form, r: rosette form, t: tussock form, l: liane form, Ogr: other growth forms.
|
Life forms |
C1 |
C2 |
All |
|
1.1 |
1.2 |
1.3 |
2.1 |
2.2 |
2.3 |
2.4 |
|
MIV |
PS |
MIV |
PS |
MIV |
PS |
MIV |
PS |
MIV |
PS |
MIV |
PS |
MIV |
PS |
MIV |
PS |
|
Dormancy |
MM |
43.3
|
20.8
|
48.6
|
20.4
|
42.0
|
17.9
|
35.6
|
21.4
|
49.4
|
26.2
|
59.1
|
23.3
|
54.6
|
30.0
|
47.4
|
17.6 |
|
M |
11.4 |
13.9 |
9.1 |
12.7 |
10.2 |
9.4 |
6.0 |
8.6 |
11.2 |
14.3 |
6.0 |
14.4 |
10.1 |
10.0 |
9.5 |
10.6 |
|
N |
17.1 |
18.8 |
18.6 |
15.8 |
19.8 |
17.9 |
24.7 |
21.4
|
19.6 |
22.6 |
13.4 |
21.1 |
10.3 |
18.8 |
18.2 |
14.5 |
|
Ch |
1.7 |
5.0 |
1.6 |
5.0 |
2.6 |
5.2 |
5.8 |
5.7 |
2.9 |
6.0 |
0.1 |
1.1 |
1.8 |
5.0 |
2.1 |
5.2 |
|
H |
11.1 |
16.8 |
12.7 |
18.6 |
11.5 |
21.2
|
7.3 |
15.7 |
8.4 |
11.9 |
5.8 |
7.8 |
6.2 |
11.3 |
10.5 |
21.5
|
|
G |
8.8 |
14.9 |
5.0 |
16.3 |
5.7 |
12.3 |
4.5 |
7.1 |
3.0 |
8.3 |
4.8 |
13.3 |
3.4 |
6.3 |
5.2 |
15.5 |
|
Th |
4.1 |
8.9 |
3.5 |
10.0 |
7.7 |
14.6 |
16.1 |
20.0 |
5.2 |
9.5 |
10.8 |
18.9 |
13.5 |
18.8 |
6.5 |
13.0 |
|
HH |
2.5 |
1.0 |
0.9 |
0.9 |
0.3 |
0.9 |
- |
- |
0.2 |
1.2 |
- |
- |
- |
- |
0.6 |
0.9 |
|
HH(Th) |
- |
- |
0.1 |
0.5 |
0.2 |
0.5 |
- |
- |
- |
- |
- |
- |
- |
- |
0.2 |
1.2 |
|
Radicoid |
R1-2
|
- |
- |
0.2 |
0.9 |
0.1 |
0.5 |
- |
- |
- |
- |
- |
- |
- |
- |
0.1 |
0.6 |
|
R2-3
|
3.0 |
7.9 |
1.5 |
7.2 |
2.7 |
8.1 |
4.4 |
4.3 |
3.1 |
4.8 |
1.7 |
1.1 |
0.7 |
2.5 |
2.1 |
8.2 |
|
R3
|
17.0 |
22.8 |
11.2 |
20.4 |
13.9 |
22.7 |
10.5 |
17.1 |
8.8 |
17.9 |
4.0 |
11.1 |
3.2 |
12.5 |
10.5 |
20.9 |
|
R3(b)
|
- |
- |
0.1 |
0.5 |
0.1 |
0.5 |
- |
- |
- |
- |
0.4 |
1.1 |
- |
- |
0.1 |
0.6 |
|
R3(o)
|
- |
- |
- |
- |
0.1 |
0.5 |
0.5 |
1.4 |
- |
- |
- |
- |
- |
- |
0.0 |
0.3 |
|
R3(s)
|
2.4 |
2.0 |
3.6 |
3.6 |
3.1 |
2.8 |
1.3 |
1.4 |
3.6 |
4.8 |
2.1 |
5.6 |
3.3 |
5.0 |
3.1 |
2.4 |
|
R3(v)
|
0.5 |
2.0 |
0.3 |
2.3 |
0.5 |
2.8 |
0.5 |
1.4 |
0.2 |
1.2 |
0.1 |
1.1 |
- |
- |
0.3 |
3.0 |
|
R4
|
3.7 |
3.0 |
6.1 |
1.8 |
3.7 |
2.4 |
1.1 |
2.9 |
2.7 |
1.2 |
4.2 |
1.1 |
3.1 |
1.3 |
4.5 |
2.7 |
|
R5
|
73.0
|
60.4
|
76.4
|
60.6
|
75.3
|
56.9
|
81.3
|
70.0
|
81.1
|
69.0
|
87.4
|
78.9
|
89.4
|
76.3
|
78.8
|
58.5
|
|
R5(s)
|
0.2 |
1.0 |
0.5 |
1.8 |
0.3 |
1.4 |
0.5 |
1.4 |
0.4 |
1.2 |
- |
- |
0.4 |
2.5 |
0.4 |
1.5 |
|
R5(t)
|
0.2 |
1.0 |
0.1 |
0.9 |
0.1 |
0.5 |
- |
- |
- |
- |
- |
- |
- |
- |
0.1 |
0.6 |
|
R5(v)
|
- |
- |
- |
- |
0.1 |
0.5 |
- |
- |
- |
- |
- |
- |
- |
- |
0.0 |
0.3 |
|
R5(c)
|
- |
- |
- |
- |
0.1 |
0.5 |
- |
- |
- |
- |
- |
- |
- |
- |
0.0 |
0.3 |
|
Disseminule |
D1
|
13.6 |
19.8 |
9.5 |
24.0 |
11.8 |
19.4 |
15.5 |
24.3 |
10.5 |
16.7 |
10.2 |
16.7 |
13.0 |
25.0 |
10.9 |
22.4 |
|
D1,2
|
- |
- |
0.0 |
0.5 |
0.1 |
0.5 |
- |
- |
- |
- |
- |
- |
- |
- |
0.0 |
0.6 |
|
D1,3
|
- |
- |
0.0 |
0.5 |
0.1 |
0.9 |
0.5 |
1.4 |
- |
- |
- |
- |
- |
- |
0.1 |
1.2 |
|
D1,4
|
0.2 |
1.0 |
0.1 |
0.5 |
0.5 |
1.4 |
0.4 |
1.4 |
- |
- |
- |
- |
- |
- |
0.2 |
1.2 |
|
D2
|
16.7 |
15.8 |
22.6 |
19.9 |
21.5 |
18.0 |
24.0 |
24.3 |
21.6 |
25.0 |
21.5 |
30.0 |
24.1 |
22.5 |
21.9 |
18.5 |
|
D2,4
|
11.1 |
11.9 |
6.8 |
8.1 |
7.0 |
8.1 |
4.2 |
5.7 |
9.2 |
11.9 |
5.3 |
15.6 |
8.7 |
11.3 |
7.2 |
7.3 |
|
D3
|
21.2 |
5.9 |
24.7 |
5.9 |
23.3 |
7.6 |
21.3 |
5.7 |
30.4
|
6.0 |
34.7
|
3.3 |
31.5
|
5.0 |
26.2 |
7.3 |
|
D4
|
35.6
|
42.6
|
35.3
|
38.0
|
33.6
|
41.7
|
31.4
|
35.7
|
27.5
|
38.1
|
27.7
|
32.2
|
21.6
|
32.5
|
32.2
|
39.1
|
|
D4,1
|
0.8 |
2.0 |
0.1 |
0.9 |
0.6 |
1.4 |
- |
- |
- |
- |
- |
- |
- |
- |
0.3 |
1.2 |
|
D4,2
|
0.7 |
1.0 |
0.6 |
1.4 |
1.2 |
0.5 |
2.6 |
1.4 |
0.2 |
1.2 |
0.6 |
1.1 |
0.7 |
2.5 |
0.8 |
0.9 |
|
D5
|
- |
- |
0.1 |
0.5 |
0.2 |
0.5 |
- |
- |
0.6 |
1.2 |
0.1 |
1.1 |
0.5 |
1.3 |
0.2 |
0.3 |
|
Growth |
b |
0.4 |
1.0 |
0.2 |
1.4 |
0.9 |
1.9 |
3.2 |
2.9 |
0.2 |
1.2 |
0.1 |
1.1 |
0.6 |
1.3 |
0.5 |
1.8 |
|
b-l |
- |
- |
0.7 |
1.8 |
0.8 |
0.9 |
1.8 |
2.9 |
0.2 |
1.2 |
0.6 |
3.3 |
0.2 |
1.3 |
0.6 |
1.5 |
|
b-p |
1.5 |
2.0 |
0.7 |
1.8 |
1.5 |
1.9 |
1.4 |
1.4 |
0.2 |
1.2 |
2.9 |
1.1 |
2.4 |
1.3 |
1.3 |
1.8 |
|
b-pr |
0.2 |
1.0 |
- |
- |
0.1 |
0.5 |
0.5 |
1.4 |
0.2 |
1.2 |
0.7 |
1.1 |
0.5 |
1.3 |
0.2 |
0.3 |
|
b-ps |
0.5 |
2.0 |
0.5 |
1.4 |
0.6 |
1.9 |
0.5 |
1.4 |
0.6 |
2.4 |
- |
- |
- |
- |
0.4 |
1.2 |
|
e |
72.6
|
59.4
|
71.6
|
57.0
|
68.5
|
55.5
|
60.4
|
48.6
|
69.8
|
54.8
|
74.8
|
56.7
|
71.1
|
52.5
|
70.8
|
52.1
|
|
e,b |
0.8 |
1.0 |
1.3 |
1.8 |
1.6 |
3.8 |
1.6 |
1.4 |
- |
- |
0.3 |
2.2 |
0.2 |
1.3 |
1.1 |
3.0 |
|
l |
7.0 |
9.9 |
7.9 |
10.4 |
8.4 |
8.1 |
8.8 |
8.6 |
14.1 |
15.5 |
8.4 |
14.4 |
13.0 |
16.3 |
9.0 |
8.8 |
|
l-b |
0.9 |
1.0 |
1.1 |
1.8 |
1.8 |
1.9 |
1.8 |
2.9 |
0.8 |
2.4 |
0.5 |
1.1 |
1.3 |
3.8 |
1.2 |
1.5 |
|
p |
2.6 |
1.0 |
6.0 |
0.9 |
3.1 |
0.5 |
0.7 |
1.4 |
2.7 |
1.2 |
4.2 |
1.1 |
3.1 |
1.3 |
4.2 |
0.6 |
|
p-l |
0.2 |
1.0 |
1.4 |
0.9 |
2.5 |
1.4 |
2.3 |
2.9 |
1.7 |
2.4 |
1.6 |
2.2 |
0.6 |
2.5 |
1.6 |
0.9 |
|
pr |
1.5 |
3.0 |
0.9 |
3.6 |
2.0 |
3.8 |
6.0 |
4.3 |
0.8 |
3.6 |
0.8 |
1.1 |
0.2 |
1.3 |
1.3 |
3.6 |
|
ps |
1.4 |
3.0 |
1.3 |
4.1 |
1.1 |
2.4 |
2.6 |
4.3 |
3.8 |
3.6 |
1.5 |
5.6 |
2.7 |
6.3 |
1.7 |
5.5 |
|
r |
1.6 |
5.9 |
0.5 |
4.5 |
0.7 |
3.8 |
0.9 |
2.9 |
0.2 |
1.2 |
1.7 |
4.4 |
2.2 |
1.3 |
0.9 |
6.1 |
|
t |
8.8 |
8.9 |
5.9 |
8.6 |
6.0 |
10.9 |
6.9 |
11.4 |
4.8 |
8.3 |
1.8 |
4.4 |
2.0 |
8.8 |
5.3 |
10.3 |
|
t-p |
- |
- |
- |
- |
0.1 |
0.9 |
0.7 |
1.4 |
- |
- |
- |
- |
- |
- |
0.1 |
0.9 |
Download Excel Table
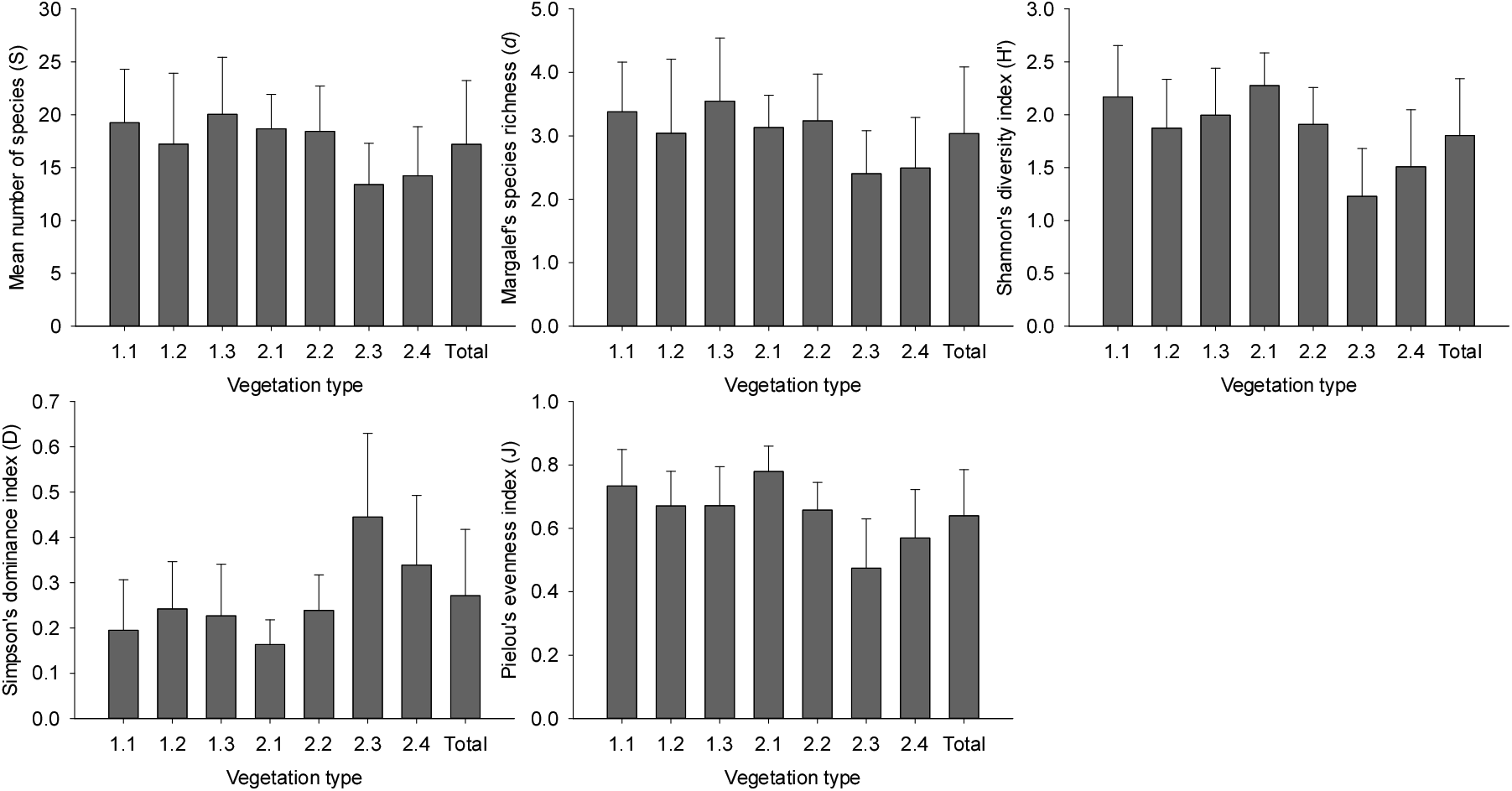
한편, 종다양성 경향을 단위 면적당(/100 m2) 종풍부도(S), 종균재도(J′), 종다양도(H′) 그리고 종우점도(D)의 4가지 지수로 파악한 결과(Figure 6), 특정 임분의 생물다양성을 가장 간략하게 평가하는 지수인 종풍부도는 전체 평균 17종이었고 북향 사면하부 적윤지를 특징짓는 사위질빵-인동덩굴형(Type 1.3)이 평균 20종으로 가장 풍부하였고, 동향 사면중부 건성지를 특징짓는 참느릅나무-팽나무형(Type 2.3)이 평균 13종으로 가장 적게 나타났다. 종다양도는 전체 평균 1.804이었고 찔레꽃_쑥형(Type 2.1)이 2.276으로 가장 높았고, 참느릅나무-팽나무형(Type 2.3)이 1.228로 가장 낮았으며, 종균재도는 전체 평균 0.640이었고 찔레꽃-쑥형(Type 2.1)이 0.779로 가장 높았고, 참느릅나무-팽나무형(Type 2.3)이 0.474로 가장 낮게 나타났는데 대개 해발고도가 높은 유형일수록 종다양성과 종균재도가 모두 높은 경향이었다. 종우점도는 전체 평균 0.271로 다수의 종이 우점하고 있었고 참느릅나무-팽나무형(Type 2.3)이 0.445로 가장 높았고, 찔레꽃-쑥형(Type 2.1)이 0.163으로 가장 낮았다.
Figure 6.
Alpha diversity indices (mean±SD) for the seven vegetation types classified in Robinia pseudoacacia plantations.
Download Original Figure
3. 지표종
경북 일대 산지 아까시나무림에서 구분된 7개 식생유형 간 지표종(indicator species)을 30% 이상의 IndVal(P < 0.05)을 갖는 종을 기준으로 식별한 결과(Table 3), 모두 55종의 지표종이 식별되었다. 식생유형 간에는 신갈나무-둥굴레형(Type 1.1)은 신갈나무, 둥굴레, 쇠물푸레나무 등 모두 15종으로 7개 식생유형 중 지표종 수가 가장 많았으며, 대개 풍수산포형인 대형지상식물(MM)과 반지중식물(H) 요소가 특징짓고 있었다. 밤나무-청미래덩굴형(Type 1.2)은 대형지상식물인 밤나무와 미소지상식물(N)인 청미래덩굴의 2종으로 7개 식생유형 중 지표종 수가 가장 적었다. 사위질빵-인동덩굴형(Type 1.3)은 인동덩굴과 사위질빵 등 대개 풍수산포형 만목성 식물이 특징짓고 있었다. 찔레꽃-쑥형(Type 2.1)은 쑥, 쇠별꽃, 개망초, 애기똥풀, 찔레꽃 등 모두 14종의 다양한 지표종이 출현하였고, 대개 적윤한 임연부에 출현하는 초본성 식물들이 상위 지표값을 특징짓고 있었다. 굴참나무-감태나무형(Type 2.2)은 감태나무, 댕댕이덩굴, 가죽나무 등 모두 11종이 지표종으로 출현하였고, 대개 목본성 식물이 특징짓고 있었다. 참느릅나무-팽나무형(Type 2.3)은 참느릅나무, 팽나무, 굴참나무 등 모두 4종의 지표종이 출현하였고, 대개 대형지상식물이 특징짓고 있었다. 귀룽나무-푼지나무형(Type 2.4)은 갈퀴덩굴, 푼지나무, 귀룽나무 등 모두 6종의 지표종이 출현하였고, 일년생식물과 동물산포형 식물이 특징짓고 있었다.
Table 3.
List of indicator species with their IndVal (Indicator Value) percentage and p value for the seven vegetation types of Robinia pseudoacacia plantations.
|
Species |
Observed indicator value |
IV from randomized groups |
P
|
Species |
Observed indicator value |
IV from randomized groups |
P
|
|
IndVal(%) |
Mean |
S.Dev |
|
IndVal(%) |
Mean |
S.Dev |
|
|
Quercus mongolica-Polygonatum odoratum var. pluriflorum type
|
Alnus japonica
|
38.3 |
16.2 |
6.6 |
* |
|
Quercus mongolica
|
66.3 |
13.9 |
7.0 |
*** |
Rubus parvifolius
|
37.0 |
14.3 |
7.1 |
* |
|
Polygonatum odoratum var. pluriflorum
|
69.1 |
13.7 |
7.1 |
*** |
Bromus remotiflorus
|
38.3 |
16.2 |
6.7 |
* |
|
Fraxinus sieboldiana
|
51.6 |
15.0 |
7.4 |
*** |
Cyrtomium fortunei
|
38.3 |
16.1 |
6.8 |
* |
|
Lindera obtusiloba
|
43.4 |
13.4 |
6.6 |
** |
Spiraea prunifolia f. simpliciflora
|
35.5 |
14.2 |
7.2 |
* |
|
Carex humilis var. nana
|
46.9 |
13.9 |
7.4 |
** |
Celastrus orbiculatus
|
32.8 |
13.8 |
7.0 |
* |
|
Carex fililpes var. oligostachys
|
38.3 |
16.1 |
7.2 |
** |
Paederia foetida
|
34.1 |
14.5 |
7.1 |
* |
|
Athyrium yokoscense
|
45.8 |
13.8 |
7.4 |
** |
Quercus variabilis-Lindera glauca type
|
|
|
|
|
|
Syneilesis palmata
|
38.3 |
15.9 |
7.1 |
** |
Lindera glauca
|
44.0 |
13.5 |
6.8 |
*** |
|
Persicaria nepalensis
|
38.3 |
16.1 |
7.1 |
** |
Cocculus orbiculatus
|
39.9 |
13.9 |
7.1 |
** |
|
Hosta longipes
|
38.3 |
16.0 |
7.0 |
** |
Quercus variabilis
|
34.4 |
13.8 |
7.0 |
** |
|
Lonicera praeflorens
|
38.3 |
16.0 |
7.3 |
** |
Ailanthus altissima
|
34.4 |
13.8 |
7.0 |
** |
|
Prunus serrulata var. pubescens
|
35.4 |
13.7 |
7.2 |
** |
Securinega suffruticosa
|
34.4 |
13.9 |
7.1 |
* |
|
Fraxinus rhynchophylla
|
40.4 |
15.0 |
7.4 |
** |
Cardamine flexuosa
|
35.3 |
13.7 |
6.8 |
* |
|
Rhododendron mucronulatum
|
30.1 |
13.7 |
7.2 |
* |
Cornus walteri
|
33.6 |
14.8 |
7.3 |
* |
|
Artemisia keiskeana
|
30.0 |
14.9 |
7.3 |
* |
Diospyros lotus
|
33.3 |
13.9 |
7.0 |
* |
|
Castanea crenata-Smilax china type
|
|
|
|
Pyrus ussuriensis |
30.8 |
15.3 |
7.6 |
* |
|
Castanea crenata
|
33.5 |
13.6 |
6.8 |
* |
Boehmeria platanifolia
|
31.2 |
14.7 |
7.5 |
* |
|
Smilax china
|
30.3 |
13.5 |
6.6 |
* |
Zanthoxylum schinifolium
|
31.6 |
13.6 |
7.0 |
* |
|
Clematis apiifolia-Lonicera japonica type
|
|
|
|
Ulmus parvifolia-Celtis sinensis type
|
|
|
|
|
|
Lonicera japonica
|
31.0 |
13.7 |
7.0 |
* |
Ulmus parvifolia
|
46.0 |
13.9 |
7.4 |
** |
|
Conyza canadensis
|
30.8 |
14.7 |
7.3 |
* |
Celtis sinensis
|
44.2 |
13.5 |
7.0 |
** |
|
Clematis apiifolia
|
30.1 |
13.8 |
7.4 |
* |
Quercus variabilis
|
39.4 |
13.2 |
6.3 |
** |
|
Rosa multiflora-Artemisia indica type
|
|
|
|
Chenopodium album var. centrorubrum |
30.2 |
14.4 |
7.2 |
* |
|
Artemisia indica
|
57.1 |
13.8 |
7.2 |
*** |
Prunus padus-Celastrus flagellaris type
|
|
|
|
|
Stellaria aquatica
|
50.4 |
14.3 |
7.2 |
** |
Celastrus flagellaris
|
63.0 |
13.8 |
7.3 |
*** |
|
Erigeron annuus
|
51.9 |
14.2 |
7.1 |
** |
Galium spurium
|
54.4 |
13.9 |
7.6 |
*** |
|
Chelidonium majus subsp. asiaticum
|
47.3 |
13.7 |
7.0 |
** |
Prunus padus
|
41.9 |
14.3 |
7.2 |
** |
|
Rosa multiflora
|
43.2 |
13.6 |
6.9 |
*** |
Liriope muscari
|
38.5 |
13.6 |
7.2 |
* |
|
Quercus acutissima
|
34.9 |
13.4 |
6.5 |
* |
Commelina communis
|
31.7 |
13.7 |
6.8 |
* |
|
Aconitum jaluense
|
38.3 |
16.0 |
6.5 |
* |
Fallopia dumetorum
|
30.3 |
14.3 |
7.0 |
* |
Download Excel Table
결 론
본 연구는 경북 일대 산지에 분포하는 아까시나무림의 자연성 회복을 위한 생태적 관리에 필요한 기초자료를 제공할 목적으로 총 200개 지소의 임분에서 수집한 식생자료를 이용하여 종조성적 측면에서 식생유형을 분류하고 DCA 분석으로 환경구배에 따른 그 공간배열 상태를 파악함과 아울러 군락적합도 판정에 의해 결정된 진단종을 기준으로 군락조성요약표를 작성하였다. 경북 일대 산지 아까시나무림의 종조성적 식생유형은 신갈나무-둥굴레형, 밤나무-청미래덩굴형, 철쭉-인동덩굴형, 찔레꽃-쑥형, 감태나무-댕댕이덩굴형, 참느릅나무-팽나무형, 그리고 귀룽나무-푼지나무형 등 모두 7개 유형으로 분류되었으며, 대개 해발고도, 수분체계, 천이단계 및 교란체제 등 복합적 요인의 차이에 의해 구분되는 경향이었다. 또한 찔레꽃-쑥형은 Jegal and Kim(2003)이 경북 일원 화성암산지와 퇴적암산지의 산림식생 식생비교 연구에서 분류한 아까시나무-찔레꽃군락 그리고 참느릅나무-팽나무형과 귀룽나무-푼지나무형은 Cho and Kim(2005)이 우리나라 아까시나무림의 군락분류 연구에서 기재한 아까시나무-닭의장풀군집의 하위단위인 전형아군집과 유사한 종조성을 나타내고 있음을 알 수 있었다. 구성종의 생태적 우세정도를 가늠하는 평균상대중요치(MIV)는 전체적으로 아까시나무(39.7)가 단연 높게 나타났지만 굴참나무, 졸참나무, 신갈나무, 상수리나무, 갈참나무 등 참나무류가 상위중요치 구성종으로 나타나고 있어 잠재천이경향을 가늠할 수 있었다. 생활형 조성 체계(휴면형-지하기관형-산포기관형-생육형)는 평균상대중요치(MIV) 기준에서는 ‘MM-R5-D1-e’형, 종수 구성비(PS) 기준에서는 ‘H-R5-D4-e’형으로 나타났다. 종다양도(H′)와 종균재도(J′)는 해발고도가 높은 유형일수록 모두 높은 경향이었다. 특정 임분의 입지환경 상태를 반영하는 척도인 지표종(indictor species)은 모두 55종이 식별되었으며 식생유형 간 지표종 구성은 대개 휴면형 차이로 나타났다. 끝으로 본 연구에서 도출된 식생유형과 관련 생태정보는 향후 우리나라 산지 아까시나무림의 자연성 회복을 위한 생태적 관리 방안을 마련하는데 있어 하나의 기초자료로 활용 될 수 있지만, 종조성적 식생유형에 대한 산림분야에서의 범용적 적용을 위해서는 생육지 토양 특성, 수분 체계 등 입지환경과의 관계 및 천이 경향 등을 포함한 보다 넓고 장기적인 추가연구가 수행되어야 할 것으로 생각되었다.