Article
정주지 단풍나무의 탄소저장량 추정 상대생장식
김호진
1, 백경원
1, 이지현
1, 이지현
1, 이정민
2, 손요환
2, 김춘식
1,*
Allometric Equations for Estimating the Carbon Storage of Maple Trees in an Urban Settlement Area
Hojin Kim
1, Gyeongwon Baek
1, Byeonggil Choi
1, Jihyun Lee
1, Jeongmin Lee
2, Yowhan Son
2, Choonsig Kim
1,*
1Division of Environmental and Forest Science, Gyeongsang National University, Jinju 52725, Korea
2Division of Environmental Science and Ecological Engineering, Korea University, Seoul 02841, Korea
© Copyright 2023 Korean Society of Forest Science. This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received: Jan 11, 2023; Revised: Mar 06, 2023; Accepted: Mar 08, 2023
Published Online: Mar 31, 2023
요약
본 연구는 도시 내 정주지에 조경수로 식재된 단풍나무(Acer palmatum Thunb.) 20본을 대상으로 로그(Log)와 일반화적률법(Generalized Method of Moments, GMM)에 의해 수목 부위별 탄소저장량 추정식을 개발하였다. 수목 부위별 탄소 농도는 줄기 목질부가 49.8%로 가장 높게 나타났으며, 줄기 수피는 46.5%로 가장 낮은 값을 보였다. 수목 부위별 탄소저장량을 종속변수로 하고 흉고직경(DBH), 흉고직경 제곱과 수고(DBH2×H)를 독립변수로 하는 상대생장식의 결정계수(R2)는 줄기, 뿌리, 지상부, 임목 전체의 경우 Log 식은 0.7494~0.9036, GMM 상대생장식은 0.7085~0.8847 등으로 두 방법 사이에 큰 차이가 없었다. 그러나 가지나 잎의 상대생장식은 R2=0.3027~0.6380으로 두 방법 모두 매우 낮게 나타났다. 본 연구는 정주지 수목의 탄소저장량 추정을 위한 상대생장식 개발에 있어서 표본목 수가 작거나 상대생장식의 잔차 이분산성이 문제가 되는 경우 GMM 방법의 상대생장식을 이용할 수 있음을 보여주고 있다.
Abstract
Using the logarithmic methods and the generalized method of moments (GMM), this study developed carbon storage equations for maple trees (Acer palmatum Thunb.) planted in an urban settlement area. A total of 20 maple trees of various ages and diameters were destructively harvested to determine their dry weight and carbon concentration by component. The allometric equations with DBH and DBH2×H as independent variables were developed to estimate the carbon storage for each tree component. The carbon concentration of tree components was the highest in stem wood (49.8%) and lowest in stem bark (46.5%). Allometric equations to estimate the carbon storage of tree components (stem, root, aboveground, and total) showed a similar coefficient of determinations (R2) between the allometric equations of the logarithmic method (0.7494-0.9036) and the GMM (0.7085-0.8847). However, the R2 values of the leaves and branches were in the range of 0.3027 to 0.6380, lower than those of the R2 of the other tree components. These results indicate that the carbon storage of maple trees growing in urban settlement areas can be efficiently predicted from the equations of GMM methods in the case of a small sample size or the heteroscedasticity of logarithmic equations.
Keywords: allometric equation; carbon stock; climate change; generalized method of moments; urban forest
서 론
도시지역 정주지의 수목 식재와 녹지공간의 확충은 수목 및 토양에 CO2를 저장할 수 있는 효과 외에도 도시 경관 및 열섬효과 개선, 여가 공간 제공, 미세먼지 저감, 건물의 냉난방을 위한 에너지 수요를 낮추는 등의 다양한 생태계 서비스 기능을 제공해주고 있다(Park and Kang, 2010; Roy et al., 2012; Seamans, 2013). 이러한 도시 수목이 주는 다양한 생태계 서비스 기능들은 지구 환경보전에 긍정적인 영향을 끼쳐 도시화에 따른 생태계 서비스 저하를 완화하는 역할을 할 수 있다(Roy et al., 2012).
IPCC에서는 LULUCF (Land Use, Land Use Change and Forestry)의 토지이용 범주를 산림지, 농경지, 초지, 습지, 정주지, 기타 토지로 구분하고 이들 지역의 온실가스 배출량 및 흡수량의 국가보고서를 요구하고 있다(IPCC, 2006). 그러나 현재 정주지의 온실가스 배출량 및 흡수량 산정에 필수적인 식재 수종의 탄소저장량 평가를 위한 연구는 매우 미흡한 실정이다(Choi et al., 2020; Lee et al., 2022). 한편, 정주지에 식재된 수목은 생육환경이 다양하여 산림 수목의 탄소저장량 평가를 위해 개발된 추정식을 이들 지역의 수목에 적용하는 것은 상당한 오류를 야기할 수 있는 것으로 알려져 있다(McHale et al., 2009). 또한, 정주지에 식재되는 수목은 식재지가 분산되고 다양한 영급과 직경을 보이기 때문에 최대우도법(maximum likelihood estimation) 추정식에서 요구하는 관련 변수의 정규분포에 대한 가정의 위배나 잔차의 이분산성(heteroscedastcity)의 문제점이 발생할 수 있다. 이러한 경우 일반화적률법(generalized method of moments, GMM)에 의한 상대생장식(Bi et al., 2004; Wang et al., 2018)이 적용되기도 하여 중국의 산림지에서 조사된 잎갈나무, 소나무류, 자작나무 등의 바이오매스 추정 상대생장식은 GMM 방법이 일반 최소제곱법에 의한 Log 상대생장식에 비해 회귀식의 설명력이 높았다(Wang et al., 2018). 한편, Vargas-Larreta et al.(2017)은 멕시코 온대 산림지 수목의 탄소저장량 추정에 GMM 방법을 적용한 바 있다.
국내에서 수행된 정주지 수목의 탄소저장량 추정에는 수목 부위별 건중량을 종속변수로하고 흉고직경(DBH)이나 흉고직경 제곱에 수고를 곱한 값(DBH2×H)을 독립변수로 하는 상대생장식이 일반적으로 많이 이용되고 있다(Jo and Ahn, 2012; Son et al., 2014; Ha et al., 2022). 그러나 도시지역에 식재되는 수목은 가지치기가 빈번하고 수형 조절을 위한 인위적 훼손으로 상대생장식의 결정계수가 일반 산림지역에서 생육하는 임목에 비해 낮게 나타나고 있다(Park and Kang, 2010; Ha et al., 2022). 한편, Monteiro et al.(2016)은 도시지역 수목에 가지치기를 실시할 경우 탄소저장량에 영향을 끼칠 수 있는 수관 폭에 상당한 변이가 초래될 수 있다고 보고한 바 있다.
도시 내 정주지에 식재되는 수목의 탄소저장량에 대한 정확한 산정을 위해서는 상대생장식의 개발이 필요하나 시간이나 경비 등의 제한 요인으로 인하여 정주지 수목의 상대생장식 개발은 매우 제한적이다. 한편, 국내 정주지에 식재된 수종은 장미과 17.9%, 소나무과 7.9%, 단풍나무과 7.3% 정도로 알려져 있으나(Jung et al., 2013), 단풍나무(Acer palmatum Thunb.)와 관련한 연구는 미흡한 실정이다. 본 연구는 도시지역 정주지에 식재된 다양한 연령과 흉고직경을 가지는 단풍나무를 대상으로 탄소저장량을 추정을 위한 Log 및 GMM 상대생장식의 적용 가능성을 조사하였다.
재료 및 방법
1. 표본목 현황
본 연구는 경상국립대학교 종합농장(35°09'N, 128°04'E) 내 조경용으로 식재되고 정상적으로 생육하고 있는 단풍나무 20본을 대상으로 하였다. 선정된 표본목은 접목이 실시된 홍단풍나무와 청단풍나무가 고르게 섞여 있었으며, 가지치기나 초두부 상처 부위의 유합 상태를 고려할 때[Figure 1(a)] 최소 5년 이전에 수형조절을 실시한 것으로 사료되었다.
탄소저장량 추정 상대생장식 개발을 위한 표본목은 ‘산림 바이오매스 및 토양 탄소 조사·분석 표준’(Korea Forest Research Institute, 2007)에 의거 흉고직경이 고르게 분포하도록 하고 수형 등을 고려하여 총 20본을 선정하였다. 선정된 표본목은 잎의 생장이 완료된 2021년 5월 하순 지상부 0.2 m 높이를 기계톱을 이용하여 벌채하고 줄자를 이용하여 수고, 수관폭, 지하고를 측정하였다. 표본목의 연령은 13년에서 26년생까지 분포하였으며, 평균 연령은 17년이었다. 평균 흉고직경과 수고는 각 11.7 cm와 4.9 m였고, 평균 수관폭과 지하고는 각 3.3 m와 1.2 m 정도로 나타났다(Table 1).
Table 1.
General characteristics of the sampled Acer palmatum
|
Tree age (yr) |
DBH (cm) |
Height (m) |
Crown width (m) |
Clear bole length (m) |
|
17±0.9/13~26*
|
11.7±0.8/6.1~18.6 |
4.9±0.2/2.7~7.4 |
3.3±0.3/1.8~6.5 |
1.2±0.1/0.6~1.8 |
Download Excel Table
2. 표본목 조사
벌도된 표본목의 줄기로부터 가지를 분리하고 줄기의 0.2 m에서 1.2 m까지는 1 m, 1.2 m 이후부터는 2 m 간격(3.2 m, 5.2 m, 7.2 m …)으로 절단하였다. 줄기의 생중량은 10 g 단위까지 측정이 가능한 전자저울을 이용하였으며, 건중량과 탄소 농도 측정을 위해 각 절단부의 원구로부터 5 cm 두께의 원판 시료를 채취하였다. 가지 부위는 가지와 잎으로 분리하고[Figure 2(b)] 5 g 단위까지 측정이 가능한 전자저울로 생중량을 측정한 후 건중량 환산과 탄소 농도 분석을 위하여 일부 시료를 채취하였다. 뿌리의 굴취는 굴삭기를 이용하였고 뿌리에 있는 흙을 최대한 제거한 다음 전자저울을 이용하여 10 g 단위까지 측정하였다. 뿌리 분석 시료는 굵은 뿌리와 잔뿌리의 비율을 고려하여 고루 섞어 채취하였다.
실험실로 운반된 원판 시료는 85°C 온풍 건조기에서 항량에 도달할 때까지 건조하였다. 건조한 원판 시료는 수피 칼을 이용하여 목질부와 수피로 분리하고 건중량을 환산하였다. 부위별 건중량 시료는 분쇄기로 곱게 갈은 후 40메쉬(0.420 mm) 체를 이용하여 탄소 농도 분석용 시료를 조제하였다. 부위별 탄소 농도 측정은 원소분석기(Vario Macro Cube, Langensebold, Germany)를 이용하였고, 탄소 농도 값에 해당 부위의 건중량을 곱하여 각 부위의 탄소저장량으로 환산하였다.
3. 토양분석
단풍나무 식재지 주위의 토양특성 조사를 위해 400 cm3 원통형 채토기를 이용하여 0~10 cm 깊이의 토양 10점을 채취하였다. 채취한 토양 시료는 65°C의 온풍건조기에서 항량에 도달할 때까지 건조 후 2 mm 체를 이용하여 토양 물리·화학적 특성 분석용 시료를 조제하였다. 토양 입경 분포는 비중계법, 토양 탄소 농도는 원소분석기(Vario Macro Cube, Langensebold, Germany)를 이용하였다. 유효인산, 칼륨, 칼슘, 마그네슘은 토양 2.5 g을 암모늄아세테이트 용액 55 ml를 첨가하고 16시간 동안 진공 추출 후 ICP (Perkin Elmer Optima 5300, USA)로 분석하였다(Kalra and Maynard, 1991). 토양 pH와 EC는 토양시료 5 g과 증류수 25 ml의 혼합용액을 이온전극(Istec 735-P, Korea)과 EC 메터(Orion 3 star conductivity, Singapore)로 측정하였다.
4. 자료 분석
본 연구에서는 DBH와 DBH2×H 를 독립변수(X)로 하고, 줄기 목질부(stem wood), 줄기 수피(stem bark), 가지(branch), 잎(leaf), 뿌리(root), 지상부+지하부(aboveground+ root) 탄소저장량을 종속변수(Y)로 하는 상대생장식(log10Y=a+b×log10X)을 개발하였다. 한편, 본 연구에서 조사된 표본목은 연령과 식재지가 다양하고, 표본목 수가 20본 정도로 적어 추정 변수에 대한 정규분포의 가정과 상대생장식의 잔차 이분산성의 문제점이 발생할 수 있다(Bi et al., 2004; Wang et al., 2018). 이러한 문제점을 해결하기 위해 SAS의 Proc Model Procedure(SAS Institute Inc., 2003)에 도구변수(instrumental variable)로 표본목의 연령과 수관폭을 포함하여 GMM 방법에 의한 상대생장식을 개발하였다. 또한, GMM 상대생장식과 비교를 위해 탄소저장량 추정에 일반적으로 사용하는 Log 상대생장식을 SAS의 Proc Reg Procedure(SAS Institute Inc., 2003)로 계산하였고, Log 변환 상대생장식에서 나타날 수 있는 편향(bias)에 대한 보정 인자(correction factors, CF)도 제시하였다(Sprugel, 1983). 표본목의 부위별 탄소농도는 연령을 공변이로 하여 SAS의 Proc GLM Procedure로 유의성 검정 후(P<0.05), Least square means에 의해 평균 탄소 농도의 차이를 비교하였다.
결과 및 고찰
1. 토양특성
조사지의 용적밀도는 0.81 g cm−3(Table 2)로 본 연구 조사지와 인접한 조경용 느티나무 식재지의 용적밀도 0.87 g cm−3(Ha et al., 2022)과 유사하였다. 토성은 사양토, 석력 함량, 토양 pH, 전기전도도는 각각 0.24 g g−1, 6.10, 311 ㎲ cm−1로 나타났다(Table 2). 유기탄소 농도는 1.23%로 진주시 4개의 도시공원 토양 0~10 cm 깊이에 유기탄소 농도 1.12%(Ahn et al., 2022)와 큰 차이는 없었다. 교환성 칼륨, 칼슘, 마그네슘은 각각 0.24, 5.81, 0.75 cmolc kg−1로 타 연구에서 보고된 단풍나무 식재지 토양(Jo and Ahn, 2012)의 0.23, 1.19, 0.60 cmolc kg−1에 비해 칼슘 함량은 높으나 칼륨과 마그네슘 함량은 큰 차이가 없었다.
Table 2.
Soil physical and chemical properties of the sampled Acer palmatum sites.
|
Bulk density (g cm−3) |
Coarse fragment (g g−1) |
Soil particle distribution (%) |
Soil pH |
Soil EC (us cm−1) |
Organic carbon (%) |
Extractable P (mg kg−1) |
Exchangeable (cmolc kg−1) |
|
Sand |
Silt |
Clay |
K+
|
Ca2+
|
Mg2+
|
|
0.81±0.05*
|
0.24±0.03 |
62±1.3 |
30±0.8 |
8±0.6 |
6.1±0.3 |
311±45 |
1.23±0.2 |
11.3±1.2 |
0.24±0.05 |
5.81±1.18 |
0.75±0.07 |
Download Excel Table
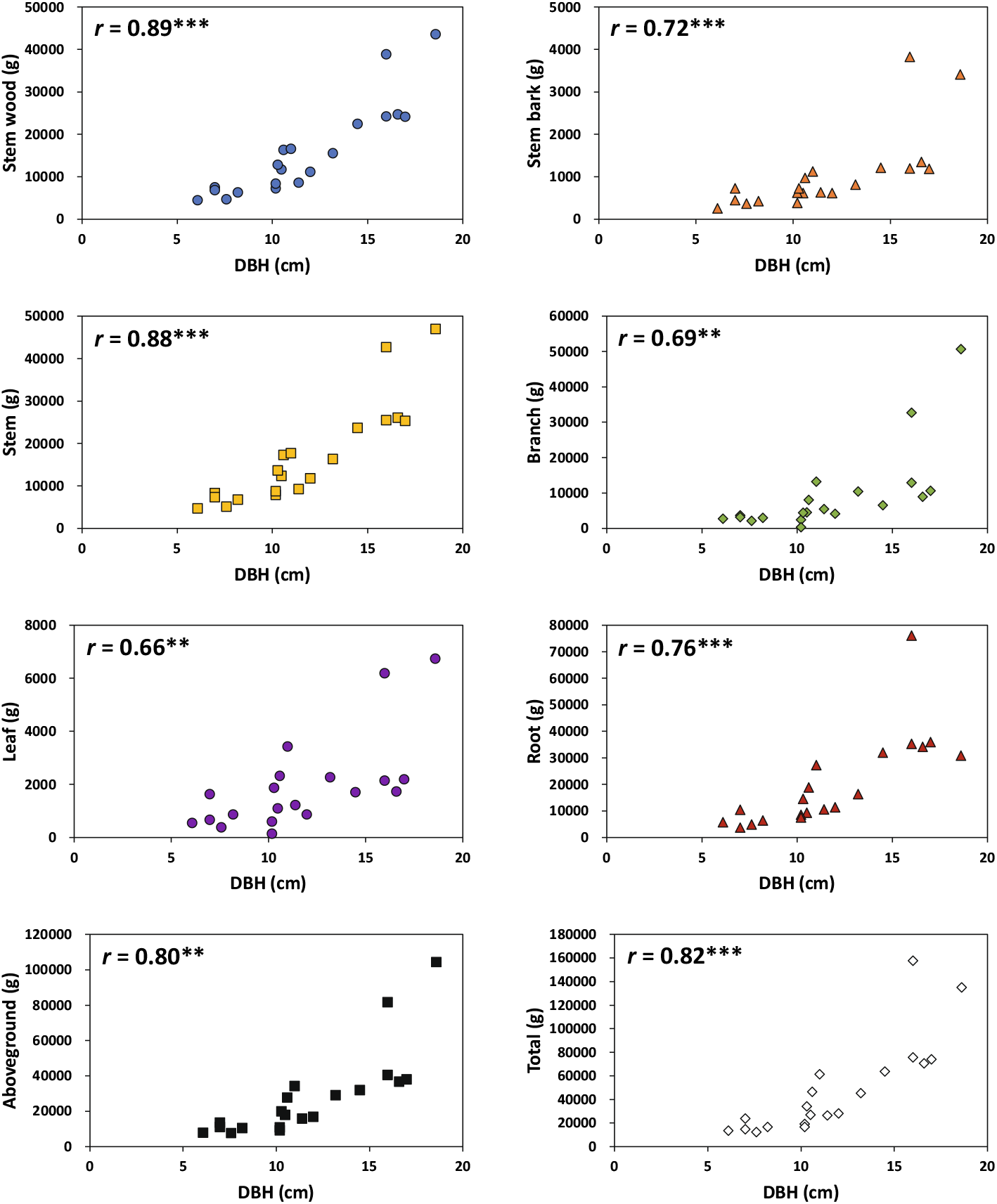
2. 표본목의 흉고직경과 부위별 건중량의 관계
표본목의 목질부, 뿌리, 지상부+지하부 총건중량은 흉고직경의 증가와 함께 강한 양의 상관관계를 보이고 있으나, 줄기 수피, 가지, 소지, 잎 등은 표본목 사이에 변이 폭이 크게 나타나 상관관계가 비교적 약하게 나타났다(Figure 2). 이는 가지치기나 초두부 절단 같은 수관부의 훼손에 의해 이들 부위의 건중량이 흉고직경 생장과 밀접한 관계가 나타나지 않은 것으로 사료된다. 한편, 본 연구에서 조사된 단풍나무 표본목 당 바이오매스 총량은 12 kg~158 kg로, 뿌리 바이오매스가 높게 나타난 일부 표본목을 제외하고 Jo and Ahn(2012)이 보고한 흉고직경 4.9~19.6 cm 범위의 조경용 단풍나무 단목 당 바이오매스 5~133 kg에 속하고 있다.
3. 표본목의 바이오매스 분포 비
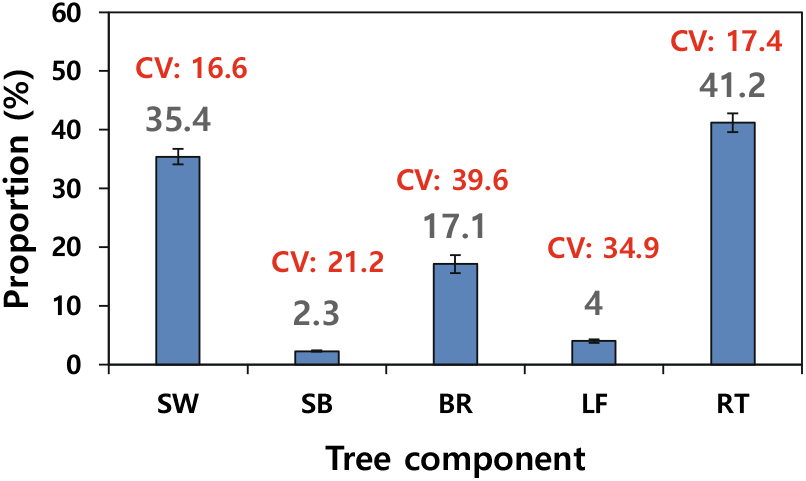
표본목의 부위별 바이오매스 분포 비율은 뿌리(41.2%), 줄기 목질부(35.4%), 가지(17.1%), 잎(4.0%), 줄기 수피(2.3%) 순이었다(Figure 2). 타 연구에서 보고된 조경용 단풍나무, 느티나무, 왕벚나무의 평균 바이오매스 분포 비율(Jo and Ahn, 2012)은 줄기(41~44%), 뿌리(26~30%), 가지(21~26%), 잎(4~7%)으로 본 연구 결과와 비교할 때 뿌리의 비율은 높고, 줄기 목질부와 가지 바이오매스는 낮게 나타났다. 본 연구에서 줄기 비율이 낮은 것은 수관을 이루는 줄기, 가지, 잎 등의 일부가 제거되어 뿌리 바이오매스 분포비가 상대적으로 높게 나타났거나, 접목에 따른 지상부와 지하부 바이오매스 축적량의 차이로 사료된다. 한편, 잎 바이오매스는 총 바이오매스의 4%로 정주지에 식재된 느티나무 4.2%, 은행나무 4.4%(Ha et al., 2022)와 큰 차이는 없었다. 도시에 식재된 수목의 바이오매스 분포비율은 생육지의 입지환경, 수형 조절을 위한 가지치기 강도(pruning intensity) 및 초두부 제거 작업 같은 관리방법에 따라 차이가 나타날 수 있는 것으로 알려져 있다(Park and Kang, 2010; Ha et al., 2022). 표본목의 바이오매스 분포에 대한 변이계수는 가지>잎>줄기 수피>뿌리>줄기 목질부 순으로(Figure 3) 수관부를 구성하는 가지와 잎의 변이 폭이 크게 나타났다.
Figure 3.
Proportion of biomass distribution in tree components in Acer palmatum. Vertical bars indicate standard errors. CV: coefficient variation.
Download Original Figure
4. 표본목의 탄소 농도
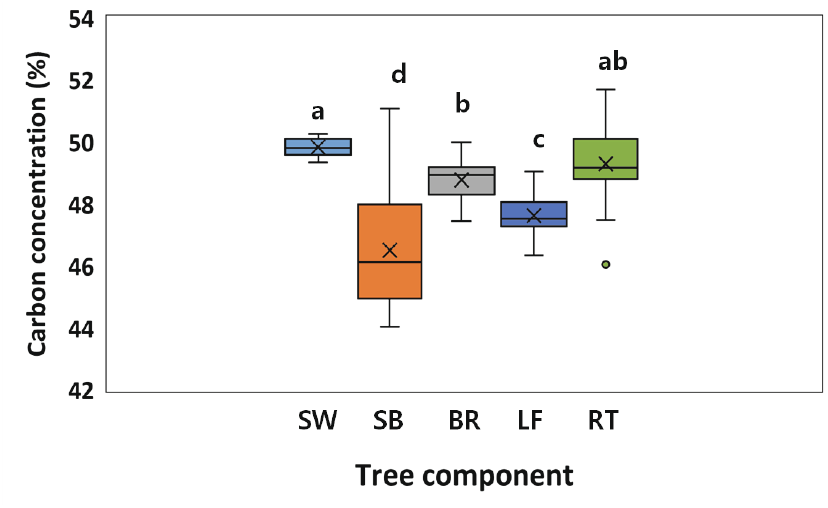
단풍나무 목질부의 탄소농도는 49.8%로 잎 47.2%, 가지 48.8%, 줄기 수피 46.5%에 비해 높은 값을 보였으나, 뿌리의 탄소 농도 49.3%와는 유의적 차이가 없었다(Figure 4). 수목의 부위별 탄소 농도는 리그닌 함량 등과 함께 생리적·화학적 요인에 따라 차이가 있는 것으로 알려져 있다(Herrero de Aza et al., 2011; Thomas and Martin, 2012; Morhart et al., 2016). 본 연구에서 나타난 목질부의 높은 탄소 농도는 이 부위의 리그닌 함량이 타 수목 부위에 비해 높기 때문일 수 있다(Novaes et al., 2010). 그러나 잎이나 수피의 탄소 농도는 타 수목 부위에 비해 낮은 값을 보이고 있으며, 이는 잎이나 형성층을 포함하는 수피는 주로 대사작용에 관여하는 기관으로 조직 내 양분 함량이 타 수목 부위에 비해 높게 나타나기 때문으로 사료된다. 예를 들면, 수목의 조직 내 양분 함량과 탄소 농도 사이는 부의 상관이 보고된 바 있다(Kim, 2019). 한편, 본 연구에서 조사된 단풍나무의 평균 탄소 농도는 48.4%로 IPCC에서 보고된 온대지역 낙엽활엽수의 평균 탄소농도 48% (IPCC, 2006)와는 차이가 없었다.
Figure 4.
Carbon concentration (%) of tree components (SW: stem wood; SB: stem bark; BR: branch; LF: leaf; RT: root) in Acer palmatum. The box represents the median and the 25th and 75th percentiles. × represents the arithmetic mean. The solid lines extend to 1.5 of the interquartile range and the values outside this range are indicated by circle. Vertical bars indicate standard error. Different letters on the bar of each tree component represent a significant difference at P<0.05.
Download Original Figure
5. 탄소저장량 추정 상대생장식
수목 부위별 탄소저장량(줄기 목질부, 줄기 수피, 가지, 잎, 뿌리, 지상부, 지상부+지하부 총량)은 DBH를 독립변수로 하는 Log 상대생장식의 결정계수가 0.3858~0.8450, GMM 상대생장식은 0.3027~0.8087로 두 방법 사이에 큰 차이는 없었으나 Log 상대생장식의 결정계수 값이 약간 높았다(Table 3). DBH2×H를 독립변수로 하는 Log 상대생장식의 결정계수는 0.5619~0.9069, GMM 상대생장식의 결정계수는 0.5055~0.8837로 DBH2×H가 DBH를 독립변수로하는 상대생장식에 비해 결정계수 값이 약간 높게 나타났다. 유사한 결과로 Wang et al.(2018)은 중국 갈참나무(Quercus aliena Bl. var. acuteserrata) 바이오매스 추정을 위한 Log 상대생장식의 결정계수는 0.9516, GMM 상대생장식은 0.9414로 두 방법 사이에 큰 차이 없음을 보고한 바 있다. 본 연구 결과는 탄소저장량 추정 상대생장식에 있어서 표본목 수가 적거나, 탄소저장량 추정 관련 변수들이 정규분포하지 않은 경우, Log 상대생장식으로부터 발생할 수 있는 잔차의 이분산성의 해결을 위해 도구변수로 표본목의 연령과 수관폭을 사용한 GMM 상대생장식의 이용이 가능함을 시사한다.
Table 3.
Allometric equations (log10Y=a+b×log10X) to estimate carbon storage (g) of tree components in Acer palmatum.
|
Variables (X) |
Tree component (Y) |
Methods |
Regression coefficient |
R
2
|
RMSE |
P-value |
CF |
|
|
|
|
a |
b |
|
|
|
|
|
DBH(cm) |
Stem wood |
Log |
1.8234 |
1.8892 |
0.8450 |
0.1180 |
<.0001 |
1.0162 |
|
GMM |
1.4097 |
2.2779 |
0.8087 |
0.1311 |
-*
|
1.0200 |
|
Stem bark |
Log |
0.8025 |
1.6926 |
0.6376 |
0.1861 |
<.0001 |
1.0407 |
|
GMM |
0.3977 |
2.0558 |
0.6012 |
0.1952 |
- |
1.0448 |
|
Stem |
Log |
1.8596 |
1.8791 |
0.8378 |
0.1206 |
<.0001 |
1.0169 |
|
GMM |
1.4435 |
2.2691 |
0.8009 |
0.1336 |
- |
1.0208 |
|
Branch |
Log |
1.1364 |
2.1944 |
0.4436 |
0.3584 |
0.0013 |
1.1594 |
|
GMM |
0.9040 |
2.4992 |
0.3989 |
0.3725 |
- |
1.1732 |
|
Leaf |
Log |
0.9446 |
1.7751 |
0.3858 |
0.3266 |
0.0035 |
1.1307 |
|
GMM |
0.2661 |
2.4798 |
0.3027 |
0.3480 |
- |
1.1496 |
|
Aboveground |
Log |
1.9580 |
1.9680 |
0.7494 |
0.1660 |
<.0001 |
1.0322 |
|
GMM |
1.4848 |
2.4258 |
0.7085 |
0.1790 |
- |
1.0376 |
|
Root |
Log |
1.6315 |
2.1289 |
0.7500 |
0.1792 |
<.0001 |
1.0377 |
|
GMM |
1.2098 |
2.5024 |
0.7188 |
0.1901 |
- |
1.0425 |
|
Total |
Log |
2.1056 |
2.0502 |
0.7786 |
0.1595 |
<.0001 |
1.0297 |
|
GMM |
1.5873 |
2.5355 |
0.7339 |
0.1748 |
- |
1.0358 |
|
DBH2xH(cm2.m) |
Stem wood |
Log |
1.4918 |
0.8317 |
0.9069 |
0.0914 |
<.0001 |
1.0097 |
|
GMM |
1.1308 |
0.9620 |
0.8847 |
0.1018 |
- |
1.0120 |
|
Stem bark |
Log |
0.4024 |
0.7822 |
0.7542 |
0.1533 |
<.0001 |
1.0274 |
|
GMM |
−0.0587 |
0.9414 |
0.7186 |
0.1640 |
- |
1.0314 |
|
Stem |
Log |
1.5241 |
0.8293 |
0.9036 |
0.0930 |
<.0001 |
1.0100 |
|
GMM |
1.1482 |
0.9649 |
0.8794 |
0.1040 |
- |
1.0125 |
|
Branch |
Log |
0.3285 |
1.1183 |
0.6380 |
0.2891 |
<.0001 |
1.1010 |
|
GMM |
0.2816 |
1.1630 |
0.6083 |
0.3007 |
- |
1.1097 |
|
Leaf |
Log |
0.2754 |
0.9103 |
0.5619 |
0.2759 |
0.0001 |
1.0916 |
|
GMM |
−0.3953 |
1.1672 |
0.5055 |
0.2931 |
- |
1.1040 |
|
Aboveground |
Log |
1.5228 |
0.8987 |
0.8654 |
0.1216 |
<.0001 |
1.0172 |
|
GMM |
0.9340 |
1.1130 |
0.8158 |
0.1423 |
- |
1.0236 |
|
Root |
Log |
1.2221 |
0.9501 |
0.8272 |
0.1490 |
<.0001 |
1.0259 |
|
GMM |
0.8977 |
1.0613 |
0.8138 |
0.1547 |
- |
1.0279 |
|
Total |
Log |
1.6756 |
0.9278 |
0.8830 |
0.1159 |
<.0001 |
1.0156 |
|
GMM |
1.1116 |
1.1292 |
0.8412 |
0.1350 |
- |
1.0212 |
Download Excel Table
한편, 본 연구의 상대생장식 결정계수는 중부지역 조경수 재배지의 단풍나무(Jo and Ahn, 2012)나, 본 연구 조사지와 인접지에 조경수로 식재된 느티나무, 은행나무, 왕벚나무(Ha et al., 2022)의 DBH 또는 DBH2×H 독립변수의 탄소저장량 추정 상대생장식의 결정계수 0.97~0.99에 비해 낮은 값을 보였다. 이는 타 연구의 경우 표본목의 수가 5~11본으로 표본목 수가 적어 회귀식의 설명력이 높게 나타났거나(Chen et al., 2021), 표본목의 흉고직경 사이 범위가 넓어 직경 생장에 따른 임목 부위별 탄소저장량의 상대생장 관계가 분명하였기 때문으로 사료된다. 그러나 DBH 또는 DBH2×H를 독립변수로 한 Log나 GMM 상대생장식 모두 가지나 잎의 결정계수는 0.3027~0.6380으로 타 임목 부위에 비해 매우 낮은 값을 보였다. 이는 도시에서 생육하는 수목은 수형조절을 위한 가지치기와 초두부 제거 작업과 같은 인위적인 관리로 인하여 수관을 구성하는 잎이나 가지의 변이 폭이 크게 나타나 해당 부위의 상대생장식 결정계수 값이 낮게 나타나는 결과로 해석된 바 있다(Ha et al., 2022). 본 연구에서도 가지와 잎의 변이계수는 34% 이상으로 타 수목부위 21% 이하에 비해 높은 값을 보였다(Figure 3). 한편, Log 변환 상대생장식에 대한 보정 인자(CF)의 경우 줄기 목질부, 줄기 수피, 뿌리, 총 탄소저장량 등은 1.05보다 작은 값을 보여 탄소저장량 추정 시 발생할 수 있는 변이(variation)가 작지만, 가지나 잎 등은 1.10 이상으로 크게 나타나 이 부위의 탄소저장량 추정 상대생장식에 CF의 사용이 필요한 것으로 나타났다.
결 론
도시지역 정주지에 조경수로 식재된 단풍나무의 수목 부위별 탄소 농도를 비교하고 탄소저장량 추정을 위한 Log 및 GMM 상대생장식을 개발하였다. 부위별 탄소 농도는 줄기 목질부(49.8%)>뿌리(49.3%)>가지(48.8%)>잎(47.6%)>줄기 수피(46.5%) 순이었으며, 줄기 수피와 잎은 타 임목 부위에 비해 유의적으로 낮은 탄소 농도를 보였다. 부위별 탄소저장량(줄기 목질부, 줄기 수피, 가지, 잎, 뿌리, 지상부, 지상부+지하부 총량) 추정 Log 및 GMM 상대생장식의 결정계수는 두 방법 사이에 큰 차이가 없었다. 한편, DBH 또는 DBH2×H를 독립변수로 한 가지나 잎의 탄소저장량 추정 상대생장식의 결정계수는 타 수목부위에 비해 매우 낮은 값을 보여 추정식의 설명력을 향상하기 위한 새로운 변수 개발이 필요한 것으로 나타났다. 본 연구결과로부터 단풍나무의 수목 부위별 탄소저장량 추정식이 개발됨으로써 정주지의 탄소 저장량 및 흡수량의 산정의 정확도를 향상할 수 있을 것으로 사료된다.
감사의 글
본 연구는 국토교통부/국토교통과학기술진흥원의 지원으로 수행되었습니다(과제번호 23UMRG-B158194-04).
References
Ahn, S.E., Lee, J. and Kim, C. 2022. Relationships between soil carbon storage and soil properties of urban parks in Jinju-si, Gyeongsangnam-do. Korean Journal of Agricultural and Forest Meteorology 24(2): 115-123.

Bi, H., Turner, J. and Lambert, M.J. 2004. Additive biomass equations for native eucalypt forest trees of temperate Australia. Trees 18(4): 467-479.

Choi, S.E., Ham, B., Song, C., Park, E,, Kim, J. and Lee, W.K. 2020. Pilot study and development of activity data for greenhouse gas inventory of settlement categories in Korea: A case of Incheon Seo-gu. Journal of Climate Change Research 11(3): 187-196.
Ha, J., Baek, G., Choi, B., Lee, J., Son, Y. and Kim, C. 2022. Development of allometric equations for carbon storage of
Ginkgo biloba Linn.,
Zelkova serrata (Thunb.) Makino. and
Prunus × yedoense Matsum. planted in Jinju-city. Journal of Climate Change Research 13(2): 135-145.
Herrero de Aza, C., Turrión, M.B., Pando, V. and Bravo F. 2011. Carbon in heartwood, sapwood and bark along the stem profile in three Mediterranean
Pinus species. Annals of Forest Science 68(6): 1067-1076.
IPCC (Intergovernmental Panel on Climate Change). 2006. 2006 IPCC guidelines for national greenhouse gas inventories. Prepared by the National Greenhouse Gas Inventories Programme. In: Eggleston, H.S., Buendia, L., Miwa, K., Ngara, T,, and Tanabe, K (eds). Japan.
Jo, H.K. and Ahn, T.W. 2012. Carbon storage and uptake by deciduous tree species for urban landscape. Journal of the Korean Institute of Landscape Architecture 40(5): 160-168.
Jung, S.S., Hwang, H.S., Yang, J.C., Oh, S.H. and Lee, Y.M. 2013. Consideration of street trees’ distribution status in Korea. In Proceedings of the 2010 Meeting of the Korean Forest Society. pp. 522-524.
Kalra, Y.P. and Maynard, D.G. 1991. Methods manual for forest soil and plant analysis. Forest Canada, Northwest region, Northern Forestry Centre. Canada. Information Report NOR-X-319. pp. 116.
Kim, C. 2019. Carbon and nitrogen distribution of tree components in
Larix kaempferi Carriere and
Quercus variabilis Blume stands in Gyeongnam province. Journal of Korean Society of Forest Science 108(2): 139-146.
Korea Forest Research Institute. 2007. Survey manual for forest biomass and soil carbon. Korea Forest Research Institute. Seoul. pp. 60.
Lee, S., Lee, S., Han, Y., Lee, J., Son, Y. and Yoon, T.K. 2022. Determining the aboveground allometric equations of major street tree species in Wonju, South Korea using the nondestructive stem analysis method. Journal of Korean Society of Forest Science 111(4): 502-510.
McHale, M.R., Burke, I.C., Lefsky, M.A., Peper, P.J. and McPherson, E.G. 2009. Urban forest biomass estimates: is it important to use allometric relationships developed specifically for urban trees? Urban Ecosystem 12: 95-113.
Monteiro, A.V., Doick, K.J. and Handley, P. 2016. Allometric relationships for urban trees in Great Britain. Urban Forestry & Urban Greening 19: 223-236.
Morhart, C., Sheppard, J.P., Schuler, J.K. and Spiecker, H. 2016. Above-ground woody biomass allocation and within tree carbon and nutrient distribution of wild cherry (
Prunus avium L.) - a case study. Forest Ecosystem 3: 4.
Park, E.J. and Kang, K.Y. 2010. Estimation of C storage and annual CO
2 uptake by street trees in Gyeonggi-do. Korean Journal of Environment and Ecology 24(5): 591-600.
Roy, S., Byrne, J. and Pickering, C. 2012. A systematic quantitative review of urban tree benefits, costs, and assessment methods across cities in different climatic zones. Urban Forestry & Urban Greening 11(4): 351-363.
SAS Institute Inc. 2003. SAS/STAT
® 9.1 User’s Guide. SAS Institute Inc., Cary, NC, USA.
Seamans, G.S. 2013. Mainstreaming the environmental benefits of street trees. Urban Forest & Urban Greening 12(1): 2-11.
Son, Y.M., Kim, R.H., Lee, K.H., Pyo, J.K., Kim, S.W., Hwang, J.S., Lee, S.J. and Park, H. 2014. Carbon emission factors and biomass allometric equations by species in Korea. Korea Forest Research Institute, Korea. pp. 95.
Sprugel, D.G. 1983. Correcting for bias in log-transformed allometric equations. Ecology 64(1): 209-210.
Thomas, S.C. and Martin, A.R. 2012. Carbon content of tree tissues: a synthesis. Forests 3(2): 332-352.
Vargas-Larreta, B., López-Sánchez, C.A., Corral-Rivas, J.J., López-Martínez, J.O., Aguirre-Calderón, C.G. and Álvarez-González, J.G. 2017. Allometric equations for estimating biomass and carbon stocks in the temperate forests of north-western Mexico. Forest 8(8): 269.
Wang, J., Zhang, L. and Feng Z. 2018. Allometric equations for the aboveground biomass of five tree species in China using the generalized method of moments. Forest Chronicle 94(3): 214-220.