
서 론
산림 바이오매스는 임목의 크기와 산림의 성숙도, 임지의 생산성(Productivity)을 대표할 수 있는 인자로 활용된다. 따라서 이를 정량적으로 평가하는 것은 산림경영과 수확뿐만 아니라 이산화탄소(CO2) 흡수를 포함한 생태계 기능과 서비스(Ecosystem service)의 평가를 위해서도 매우 중요하다(Keeling and Phillips, 2007; Kim et al., 2021; Son et al., 2015). 바이오매스와 직접적으로 관련된 목재생산, 온실가스 흡수 및 저장뿐만 아니라, 레크레이션(Kim et al., 2021; Pang et al., 2017), 토사유출 방지(Kim et al., 2017), 생물다양성 평가(Pandey et al., 2020)에 바이오매스가 입력자료로 활용된 바 있다. 또한, 기후위기 대응을 위한 국제사회 노력의 일환인 온실가스 인벤토리 보고서(National Inventory Report; NIR) 작성과 국가 온실가스 감축목표(Nationally Determined Contribution; NDC), 2050 탄소중립에 대한 계획 수립과 이행 평가의 정확도를 높이기 위해서 산림 바이오매스의 측정, 평가, 예측 고도화를 위한 연구가 활발하게 수행되고 있다(Hong et al., 2022; Kim et al., 2018; Kim et al., 2019; Lee et al., 2015).
상대생장식(Allometric equation)은 식물의 상대생장관계(Relative growth rate)를 통해 각 부분의 값을 추정하는 방법으로, 과거부터 현재까지 폭넓게 활용되는 방법론이다(Baskerville et al., 1965; Kim et al., 1995; Henry et al., 2011; Vorster et al., 2020). 대표적으로 수고와 흉고직경(Diameter at breast height; DBH)에 대한 상대생장 연구가 많이 이루어졌는데, 이는 DBH에 비해 상대적으로 복잡하고 오랜 시간이 필요한 수고의 측정을 대체하는데 유용하게 활용되어왔다(Huang et al., 1992; Lumbres et al., 2011; Sharma and Parton, 2007). 또한, 임목의 현존량 추정을 위해 개발된 상대생장식의 경우, 일반적으로 측정이 용이한 흉고직경(Diameter at breast height; DBH)이나 수고 등을 입력자료로 활용하고 있으며, 전세계적으로 온대림(Vargas-Larreta et al., 2017), 한대림(Xing et al., 2019), 열대림(Basuki et al., 2009; Cole and Ewel, 2006) 등 다양한 식생대의 수종들을 대상으로 활발하게 개발되어왔다. 우리나라도 곰솔(Park and Kim, 2018), 소나무(Son et al., 2011), 굴참나무(Son et al., 2011), 백합나무(Kang et al., 2016), 삼나무(Gwon et al., 2014), 신갈나무(Kwon and Lee, 2006) 등 다양한 수종에 대한 상대생장식이 연구되었다. 또한, 국립 산림과학원에서는 주요 22개 수종에 대한 상대생장식을 개발하여 공식적으로 발표하였으며(Son et al., 2014), 이는 우리나라 산림의 임목 축적(Kim et al., 2014), 탄소량(Kim et al., 2019; Lee et al., 2015), 생태계서비스(Kim et al., 2021) 정량화 고도화 등에 활용되어 왔다.
최근 대기중의 CO2 농도증가와 이에 따른 기후변화로 인해 수목의 상대생장관계가 바뀌고 있다는 연구결과들이 도출되면서, 기존에 개발된 상대생장식과 계수들에 대한 재검토 필요성이 제기되고 있다(Chen et al., 2020; Kim et al., 2020; Temesgen et al., 2014; Xu et al., 2022). 묘목을 대상으로 시행되는 인위적 온난화 실험에서는 CO2 시비 처리에 따라 지상부와 지하부의 비율이 달라지는 현상이 보고된 바 있으며(Chung et al., 2012; Song et al., 2019), 캐나다와 중국에서는 지역 규모의 대면적 산림에서 수고-DBH 상대생장의 변화가 확인되었고, 이러한 변화는 기후변화와 밀접한 연관성이 있는 것으로 분석되었다(Chen et al., 2020; Xu et al., 2022). Wang et al.(2006)은 중국 북동부 지역의 산림에서 겨울철 추위로 인한 스트레스가 증가하면, 수고 생장이 저하되는 대신 직경생장이 보다 활발해지는 현상을 밝혔으며, Xu et al.(2022)은 같은 지역의 잎갈나무속(Larix) 수종들의 수고-DBH 상대생장을 분석한 결과, 기후변화가 진행됨에 따라 전반적으로 수고 생장이 보다 활발해질 것으로 전망하였다.
우리나라의 경우 2009년부터 산림청에서 수종별 임분수확표를 통해 주요 수종의 상대생장 정보를 제공하고 있으며(Korea Forest Service, 2009), 이후 수종 확장, 바이오매스 정보 보완, 현실림 정보 추가 등 국민과 시대적인 요구들을 반영하여 지속적으로 개선되어 왔다(Korea Forest Service and National Institute of Forest Science, 2021). 그러나 2009년에 구축된 11개 수종과 2012년에 추가로 구축된 3개 수종의 상대생장정보는 현재까지 갱신되지 않고 초기 값 그대로 유지되고 있어 재검토가 필요한 시점이다. 본 연구에서는 2006년(제5차)부터 2020년(제7차)까지 NFI 고정표본점에서 조사된 자료를 분석하여 우리나라 주요 4개 수종(소나무, 낙엽송, 굴참나무, 신갈나무)의 수고와 DBH 생장을 파악하고, 기 개발된 함수식을 통해 수종별 수고-DBH 상대생장관계를 분석한 뒤, 이를 기존 수확표 정보와 비교하여 지난 15년간 상대생장의 변화 유무 및 정도를 파악하고자 하였다.
연구재료 및 방법
본 연구는 5-7차(2006-2020년) NFI 자료의 임목조사 자료 중, 개체목의 직경과 수고를 직접 측정하여 구축된 자료만을 활용하여 수행되었다. NFI는 각 고정 표본 조사구의 임목 중 DBH가 6 cm 이상인 모든 개체목의 DBH를 기록하며, 그 중 표준목으로 선정된 임목들의 수고도 직접 측정한다(Kim et al., 2011). 우리나라 주요 침엽수와 활엽수 중에서 조사 표본이 많고, 비교적 분포 범위가 넓은 소나무(Pinus densiflora; PD), 낙엽송(Larix kaempferi; LK), 굴참나무(Quercus variabilis; QV), 신갈나무(Quercus mongolica; QM), 이렇게 네 수종을 분석 대상으로 선정 하였으며, NFI 고정 조사구 중 자연적(산불, 산사태, 병충해 발생 등), 인위적(간벌, 숲가꾸기 시업 등) 교란이 관측된 지점의 임목들은 분석에 포함하지 않았다(Table 1). 또한, 본 연구는 수종별 상대생장 경향과 이에 대한 변화를 파악하는 것을 주목표로 함으로, 교란 유무를 제외한 다른 임분 특성들(단순림, 혼효림 등)에 따른 자료 구분은 수행하지 않았다. 최종적으로 216,204(PD: 106,105, LK: 13,308, QV: 41,065, QM: 55,726)개의 개체목 측정자료를 활용하여 수종별 수고-직경의 상대생장(Allometric growth)을 분석하였으며, 각 NFI 차수별 자료를 따로 분석하여 시기별 변화 경향도 확인하였다.
NFI 자료를 통해 분석된 수종별 수고-직경의 생장유형(Growth pattern)을 기존에 확인된 현실림의 생장특성과 비교하기 위해, 최신 수종별 임분수확표(Korea Forest Service and National Institute of Forest Science, 2021) 자료를 활용하였다. 본 연구에서는 5-7차 NFI 자료를 통해 분석된 수종별 수고-흉고직경의 분석 결과를 임분수확표에서 제공하는 현실림의 지위지수(Site index)에 따른 생장특성과 비교 분석하여, 우리나라 산림자원의 동적 변화를 파악하고 이에 대한 정책적 시사점을 도출하고자 하였다.
본 연구에서는 수종별 수고-흉고직경의 발달 관계를 설명하기 위해 기존에 개발된 7개의 상대생장식(Allometric equation)을 사용하였다(Table 2). 각 상대생장식은 다양한 선행 연구들에서 일반적으로 활용되어온 모형이며(Lumbres et al., 2011; Arcangeli et al., 2013), 종속변수인 수고(H)를 설명하기 위한 독립변수로 흉고직경(DBH) 1개만을 필요로하는 비선형 함수 형태로 되어 있다. 수종별 5-7차 NFI 자료를 상대생장식들을 통해 분석한 후, 잔차의 독립성(Independence of errors), 정규성(Normality), 등분산성(Homoscedasticity)을 검토하였다. 이후 결정계수(R2), 평균 제곱근 오차(Root Mean Square Error, RMSE), 평균잔차(residual Mean Difference, MD), 절대평균잔차(Absolute MD, AMD), Akaike Information Criterion (AIC) 값을 산출하여 각 상대생장식의 통계적인 적합성을 평가하였고, 이를 기반으로 수종별로 가장 적합한 상대생장식을 선정하였다. 본 연구는 회귀모형의 개발과 검증에 초점이 맞춰져 있지 않기 때문에, 모형분석과 검증을 위한 자료를 따로 구분하여 구축하지 않았으며, 각 상대생장식에 대한 수종별 계수 추정과 관련 통계량 산출은 오픈소스 소프트웨어인 R version 4.2.1 (R Project for Statistical Computing, Vienna, Austria)을 통해 분석하였다.
| Equation No. | Form | References |
|---|---|---|
| 1 | H = 1.2+a·(1−e−b·DBH)c | Richards 1959; Chapman 1961 |
| 2 | Yang et al. 1978 | |
| 3 | Yuancai and Parresol, 2001 | |
| 4 | Stage 1963; Zeide 1989 | |
| 5 | Schnute 1981 | |
| 6 | Ratkowsky 1990 | |
| 7 | Ratkowsky and Reedy 1986; Huang et al. 1992 |
연구 결과 및 고찰
5-7차 NFI 자료 분석 결과 각 수종의 수고와 DBH의 평균값은 계속 증가해온 것으로 확인되었다(Table 1, Figure 1). PD, LK, QV, QM의 평균 DBH는 5차 NFI 자료에서 20.5, 22.4, 18.1, 17.2 cm로 분석되었고, 6차 NFI 자료에서는 각각 11.4, 9.7, 12.4, 10.8%가 증가한 22.8, 24.6, 20.4, 19.0 cm로 확인되었다. 7차 NFI 자료를 분석한 결과 각 수종의 평균 DBH는 24.7, 26.2, 22.0, 20.3 cm까지 증가한 것으로 나타났으며, 이는 수종별 5차 평균 DBH에 비해 각각 20.6, 16.8, 21.6, 18.1% 커진 값이다. 이러한 결과는 우리나라 주요 수종의 평균 DBH는 여전히 증가하고 있으나, 임령의 증가와 임분의 성숙 등의 이유로 인해 증가율은 점차 감소하고 있다는 것을 잘 보여준다(Korea Forest Service and National Institute of Forest Science, 2021; Kim et al., 2012; Seo et al., 2001). 수종별로 비교해 보면, 5차와 7차 NFI 기간에 평균직경의 절대값은 소나무가 가장 크게 증가하였으나(+4.2 cm), 비율 기준으로는 굴참나무의 직경생장(+21.6%)이 가장 활발한 것으로 나타났다. PD, LK, QV, QM의 평균 수고는 5차에서 7차 NFI 기간 중, 각각 11.0, 17.0, 11.9, 10.9 m에서 13.2(+20.4%), 21.1(+23.7%), 14.2(+19.3%), 12.2 m(+12.7%)로 증가하였다. 이는 과거 15년(2006-2020년) 기간 동안 상대적으로 LK의 수고 생장이 활발했고, QM이 저조했다는 것을 보여준다(Table 1, Figure 1). 위의 분석내용을 통해 수종별 DBH와 수고 생장 경향을 검토해보면, PD는 직경과 수고가 유사한 비율로 증가했고, LK는 직경생장보다 수고생장이 활발하게 이루어졌다. 반면 QV와 QM은 상대적으로 수고보다 직경생장이 두드러진 것으로 확인되어, 각 수종 간에 생장 경향이 다른 것으로 나타났다. 수종별로 고유의 생장 특성을 갖는 것은 일반적이나(Korea Forest Service and National Institute of Forest Science, 2021), 지속적인 조사와 분석을 통해 이러한 특성이 시간의 흐름이나 기후변화 등의 요인에 따라 변화하는지에 대한 여부를 밝히는 것이 필요할 것으로 판단된다.

R 프로그램을 사용하여 각 상대생장식의 계수를 5-7차 NFI 수종별로 추정한 결과는 Table 3~5와 같다. 모든 계수는 매우 높은 수준(p < 0.001)에서 유의성이 입증되었으며, 모형의 적합도(R2)는 0.42~0.62 수준으로 확인되어 과거 연구들과 마찬가지로 DBH와 수고의 발달은 높은 상관성이 있는 것으로 나타났다(Lumbres et al., 2011; Kim et al., 2012). 각 모형의 R2 값을 확인한 결과 PD는 일부 모형 간에 값의 차이가 확인되었으나, 나머지 수종들은 모형 간에 차이가 거의 없는 것으로 나타났다. 모형의 신뢰도를 평가하는 통계값(R2, RMSE, MD, AMD, AIC)들을 종합적으로 검토한 결과 PD, LK, QV, QM에 대한 최적 상대생장식은 2, 1, 6, 7번 회귀모형으로 분석되었고, 잔차에 대한 가정들도 만족하는 것으로 확인되었다.
본 연구에서 확인된 수종별 최적 상대생장식을 통해 5-7차 NFI 자료 기반의 수고-DBH 발달에 대한 회귀곡선을 도출하였다(Figure 2). 분석결과 모든 수종에서 NFI 차수가 증가할수록, 동일 DBH에 대한 기대 수고값이 증가한 것으로 확인되었다. 예를 들면, 모형 분석결과 PD, LK, QV, QM의 DBH 25 cm에 대한 수고의 기댓값은 5차 NFI 자료에서 12.48, 19.17, 14.47, 13.19 m로, 7차 NFI 자료에서 13.61(+9.1%), 21.58(+12.7%), 15.76(+8.9%), 13.93 m(+5.6%)로 추정되어 수종별로 5~13%정도 증가한 것으로 나타났다. 이러한 결과는 수확표 자료와 비교했을 때 더욱 분명하게 확인된다. 5차 NFI 자료를 통해 추정된 PD의 수고-DBH 발달은 수확표의 지위지수 12에 해당하는 수고-DBH 발달과 거의 유사하나, 7차 NFI 자료를 통해 추정된 회귀곡선은 지위지수 14에 근접한 것으로 나타났다. 동일한 접근방법으로 QV는 지위지수 14에서 16으로, QM은 12에서 14에 가까워진 것으로 확인되었다. LK의 수고-DBH 발달은 본 연구에서 분석된 최적 상대생장식과 수확표에 활용된 회귀모형 간에 차이가 있어 다른 수종과 같이 직접적인 비교를 할 수는 없지만, DBH 20 cm 기준으로는 지위지수 22에서 24로, DBH 25 cm 기준으로는 지위지수 20에서 24에 가까워진 것으로 확인되었다.
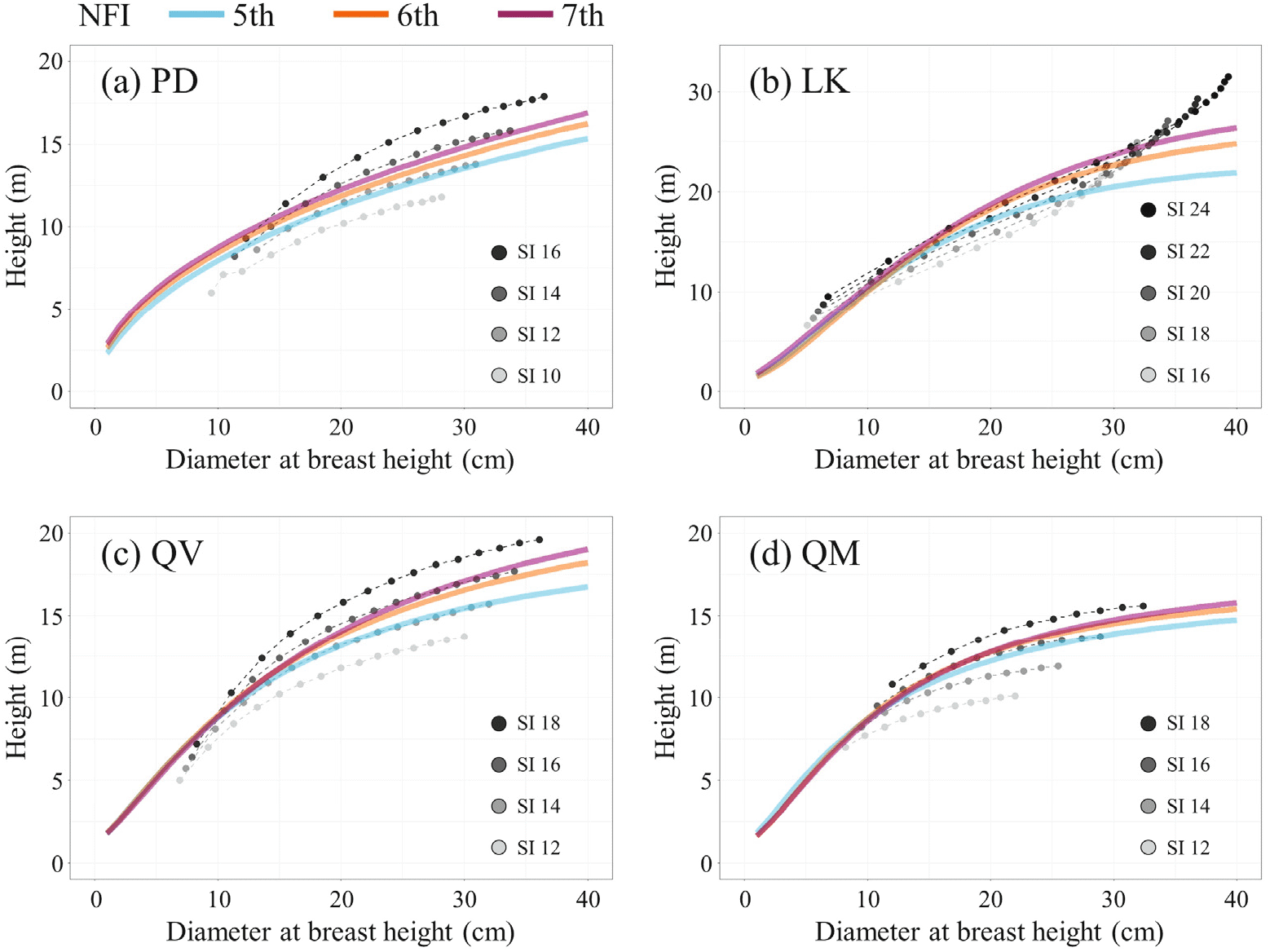
현실림에 대한 수확표 정보(Korea Forest Service and National Institute of Forest Science, 2021)와 본 연구에서 분석한 결과를 비교하여 수종별 수고-DBH 발달 경향의 변화 유무를 간접적으로 평가해보았다. 5차 NFI를 통해 파악된 수종별 평균 수고와 DBH 크기(Table 2)에 가장 근접한 임분정보를 수확표에서 찾은 결과, PD는 지위지수 12에 임령 40년(수고: 11.0 m, DBH: 20.4 cm), LK는 지위지수 18에 임령 35년(수고: 17.6 m, DBH: 22.8 cm), QV는 지위지수 12에 임령 40년(수고: 11.0 m, DBH: 17.5 cm), QM은 지위지수 12에 임령 40년(수고: 10.7 m, DBH: 16.6 cm)으로 확인되었다. 7차 NFI는 5차 NFI 이후 10년 뒤에 행해졌기 때문에 PD, LK, QV, QM의 해당 지위지수 임령에서 10년 뒤의 수고값을 확인 해본 결과, 각각 11.5(+4.5%), 19.7(11.9%), 11.6(+5.5%), 11.3 m(+5.6%)였으며, DBH는 24.3(+19.1%), 28.1(+23.2%), 20.4(+16.6%), 19.8 cm(+19.3%)로 제시되었다. 이 값들을 5-7차 NFI 자료 분석결과(Table 2)와 비교해 보면 DBH의 발달은 임분수확표 정보와 큰 차이가 없으나, 수고의 경우 모든 수종에서 임분수확표에서 확인된 값 보다 두 배 이상의 생장률을 보였다. 본 결과는 임목 조사 자료의 평균값과 수확표에 제시된 임분 단위의 평균값을 통한 간접적인 비교라는 한계가 있으나, 현재 우리나라 주요 수종들의 임목 생장이, 과거 조사자료를 통해 파악된 경험적인 수종별 수고-DBH 발달 경향과 차이가 있을 수 있음을 보여주는 의미 있는 결과라고 사료된다. 추후 지속적인 조사, 자료 구축, 분석 등을 통해 보다 명확한 결론을 도출하는 것이 필요하다고 판단된다.
또한, 본 연구의 분석결과는 이러한 상대생장의 변화가 우리나라 임분의 생산성 증가와도 밀접하게 연관되어 있다는 것을 보여준다. 현재까지도 지위지수는 임지의 생산력을 정량적으로 평가할 수 있는 중요한 지표로 고려되고 있으며, 산림경영 및 관리에서 중요하게 활용되고 있다(Kim et al., 2014; Kim et al., 2016; Yue et al., 2017). 수확표 자료를 기반으로 지위지수에 따른 임목발달을 해석해 보면, 지위지수가 높을수록 같은 직경급에서 수고의 생장이 활발한 것으로 나타난다. 즉, 본 연구의 분석결과는 2006-2020년 기간 동안 우리나라 주요 수종이 분포하는 임분의 지위지수가 전반적으로 증가했다는 것으로 해석될 수 있다. 이는 앞서 NFI 자료를 통해 산출된 임목의 수고 생장률이, 수확표 상의 고정된 지위지수에 따른 수고 생장률 보다 두 배 이상 높다는 결과와도 일치한다. 그리고 이러한 지위지수의 증가는 산림의 임분축적 및 탄소저장량 증가율이, 기존의 자료들을 통해 평가하고 예측했던 것보다 더 높은 수준으로 유지되고 있다는 것을 의미한다.
최근 몇몇 연구들에서도 대면적 산림에 대한 상대생장 및 생장량의 변화가 보고되었다(Chen et al., 2020; Kim et al., 2021; Xu et al., 2021). 중국 북부 및 북동부 지역의 NFI 조사구 중 2000-2010년 기간에 조사된 잎갈나무속(Larix) 535개 조사구 자료를 분석한 결과, 수고-DBH 발달관계가 달라지고 있다는 것이 확인되었고, 이러한 변화는 온도, 강수량, 습도 등 기후의 변화와 밀접한 연관이 있는 것으로 밝혀졌다(Xu et al., 2021). 캐나다 Ontario 지역 북방림(Boreal forest)의 고정 조사 표본점에 대한 최근 30년 조사자료를 분석한 결과에서도 평균기온과 건조도 등에 따라 임령-수고-DBH 관계의 변화가 일어난 것이 확인되었다(Chen et al., 2020). 또한, 미국 노스캐롤라이나주 듀크 포레스트(Duke Forest)에서 인위적으로 CO2 농도처리를 한 임분과 그렇지 않은 임분의 임목들을 비교 분석한 결과, 우세수종인 Loblolly pine (Pinus taeda L.)의 경우 실험군의 지상부 바이오매스 증가량이 대조군에 비해 20% 이상 높은 것으로 분석되었으며, 직경 생장에 비해 수고 생장이 더 활발하게 이루어져 상대생장식의 변화가 필요하다는 제언이 도출되었다(Kim et al., 2021).
우리나라에서는 주로 실험실 규모에서 기후(Park et al., 2016), CO2 농도(Song et al., 2020), 질소 시비(Chung et al., 2012) 등을 복합적으로 조절하여 유모나 묘목의 생장 및 생리적 반응을 평가 및 분석하는 연구들이 수행되었다. 각 연구의 실험 조건 및 대상 수종이 상이하지만, 위의 연구들은 공통적으로 기온, 강수량, 질소 등의 환경적인 요건이 제한요소로 작용하지 않는 상황에서의 CO2 시비는 임목의 생물량 증가에 긍정적인 영향을 미치며, 지상부와 지하부의 비(T/R)의 변화를 유발한다고 보고하였다. 이러한 현상은 다른 국가들의 선행연구에서도 보고된 바 있다(Huang et al., 2007; Stitt and Krapp, 1999; Way and Oren, 2010). 위에서 언급한 지역 단위의 연구 사례와 실험실 규모의 분석 결과를 종합하여 볼 때, 대기 중 CO2 농도증가와 이로 인한 기후변화가 5-7차 NFI 자료를 통해 파악된 주요 수종의 상대생장 변화에 영향을 미쳤을 것으로 사료된다. 안면도 기후변화감시소의 관측자료에 따르면, 우리나라의 2006~2020년 기간 연평균 대기 중 CO2 농도는 387.1 ppm에서 420.4 ppm으로 증가했다(http://www.climate.go.kr/). 그러나 본 연구에서 확인된 임목 전반의 생장량 증가와 수고-DBH 상대생장 변화는 우리나라 NFI의 임목조사 기준인 DBH 6 cm 이상인 임목들에 해당하는 것으로, 추후 유모에서 유령림, 장령림 등으로 이어지는 연속적인 실험과 모니터링 연구들이 필요하다고 판단된다. 또한, 다양한 기상적인 제한 요인들도 복합적으로 분석되어야만 CO2 농도 효과가 지역 및 국가 규모의 산림발달에 미치는 영향에 대한 종합적인 결론을 도출할 수 있을 것이다.
산림 생장 및 탄소 분야에서 모형 기반의 미래 예측 연구는 경혐적 자료를 통한 초기값 설정이 매우 중요하다. 이러한 모형에서 각 수종의 수고-DBH 상대생장은 산림의 바이오매스 및 탄소량 추정에 가장 기본적인 인자 중 하나로, 이에 대한 변화는 최종 결과값에 직접적인 영향을 미칠 수 밖에 없다. 한 예로 Son et al.(2014)에서 개발한 수종별 상대생장식을 통해 앞서 언급한 PD 임목의 DBH 25 cm에 대한 5차, 7차 NFI 자료의 수고 기대값을 적용하여 개체목의 수간(Stem) 바이오매스를 계산하면 120.0, 131.2 kg이며, 두 값은 약 9.3%의 차이를 갖는다. 물론 이는 PD 임목의 평균 값에 대한 결과이기 때문에, 추가적으로 임분이나 지역, 국가 단위에서 어떠한 차이를 갖는지에 대해 면밀한 분석과 검토를 해야될 필요가 있을 것이다. 그러나 분명한 것은 이러한 변화들을 수종별 국가 고유계수 등에 빠르게 반영하지 못할 경우 현재 산림 자원 및 미래 예측에 대한 불확도(Uncertainty)가 높아질 수 밖에 없으며, 이는 관련 정책의 성패에도 부정적인 영향을 미칠 것으로 사료된다.
결 론
본 연구에서는 5-7차 NFI 자료 중 임목조사 자료를 활용하여, 우리나라 주요 4개 수종의 수고-DBH 상대생장관계를 분석하고, 시간의 흐름에 따른 변화 유무와 그 경향에 대해 제시하였다. 자연적, 인위적 교란이 관측된 조사구의 자료를 제외하고, 총 198,609개 임목을 분석에 활용하였다. 먼저 DBH와 수고의 발달을 분석한 결과, 4개 수종 모두 NFI 차수에 따라 평균 DBH와 수고의 값이 증가한 것으로 확인되었으나, 그 정도와 경향은 수종 간에 차이가 있는 것으로 나타났다. 다음으로 기 개발된 7개의 수고-DBH 상대생장식 중, 수종별 최적 상대생장식을 찾고 이를 통해 각 차수별 DB에 대한 평균적인 수고-DBH 발달을 모의한 결과, 모든 수종에서 NFI 차수가 증가할수록 동일 DBH에 대한 기대 수고값이 높아지는 것으로 확인되었다. 또한, 이를 기존 수확표 자료의 수종별 수고-DBH 발달과 비교한 결과, NFI 차수가 증가할수록 점차 지위지수 값이 높은 수고-DBH 발달 곡선에 가까워지는 것으로 나타났다. 이러한 결과는 현재 우리나라 소나무, 낙엽송, 굴참나무, 신갈나무의 수고-DBH 상대생장 관계가 계속해서 변화하고 있다는 것을 보여주며, 과거 조사 및 분석을 통해 파악된 수종별 수고-DBH 발달 경향과 비교 했을 때, 직경생장보다 수고생장을 더 활발하게 하고 있다는 것을 의미한다. 기존 연구들을 통해 이러한 변화가 대기 중 CO2 농도 증가 및 이로 인한 기후변화, 질소 순환 변화 등에 영향을 받았을 가능성을 제시할 수는 있었지만, 이를 분석 및 규명하지 못했다는 한계가 있다. 따라서 지속적인 모니터링과 추가연구를 통해 이에 관한 과학적인 해석이 필요하다고 사료되며, 이는 국가의 산림탄소 인벤토리 평가 및 예측 고도화에 기여할 것으로 기대된다.