Article
소나무속 6수종 묘목의 소나무재선충병에 대한 포지 저항성 비교
김양길
1, 이다영
1, 김선정
2, 김수비
1, 최배영
3, 심동환
3, 박연일
3, 강규석
1,*
Comparing Field Resistance with Pine Wilt Disease Among Six Pine Species at Seedling Stages
Yang-Gil Kim
1, Dayoung Lee
1, Sunjeong Kim
2, Su-Vi Kim
1, Bae Young Choi
3, Donghwan Shim
3, Youn-Il Park
3, Kyu-Suk Kang
1,*
1Department of Agriculture, Forestry and Bioresources, Seoul National University, Seoul 08826, Korea
2Forest Bioresources Department, National Institute of Forest Science, Suwon 16631, Korea
3Department of Biological Sciences, Chungnam National University, Daejeon 34134, Korea
© Copyright 2023 Korean Society of Forest Science. This is an Open-Access article distributed under the terms of the
Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits
unrestricted non-commercial use, distribution, and reproduction in any
medium, provided the original work is properly cited.
Received: Feb 28, 2023; Revised: Mar 28, 2023; Accepted: Apr 05, 2023
Published Online: Jun 30, 2023
요 약
소나무재선충병(Pine wilt disease)은 소나무재선충(Bursaphelenchus xylophilus (Steiner & Buhrer, 1934))에 의해 발생하며 한국을 비롯한 동북아시아와 유럽의 소나무속(Pinus) 수목들을 고사시키고 있다. 소나무재선충병 저항성에 대한 연구는 자연상태 및 온실포트묘에서 주로 이뤄진 반면 포지에서의 연구가 부족한 편이다. 본 연구에서는 2021년 6월, 포지에 있는 소나무속 수종인 곰솔(Pinus thunbergii Parl.), 잣나무(P. koraiensis Siebold & Zucc.), 소나무(P. densiflora Siebold & Zucc.), 섬잣나무(P. parviflora Siebold & Zucc.), 리기테다소나무(P. rigida × P. taeda), 스트로브잣나무(P. strobus L.)의 묘목을 대상으로 소나무재선충을 인공접종하였다. 그리고 인공접종 후 2주 간격으로 위조지수를 측정했고, 인공접종 24주후의 고사율, 소나무재선충 검출률 및 밀도를 측정했다. 소나무재선충병에 의한 고사율은 곰솔(80%), 잣나무(77.8%), 소나무(62.5%), 섬잣나무(22.0%) 순이였으며 리기테다소나무와 스트로브잣나무는 고사하지 않았다. 소나무재선충 검출률은 리기테다소나무(22.2%)를 제외하고 모두 고사율과 동일했다. 위조지수의 경우, 소나무재선충병 고사율 순위가 높았던 곰솔, 잣나무, 소나무 3종에서 높은 수치가 나왔으며, 순위가 낮았던 섬잣나무, 리기테다소나무, 스트로브잣나무 3종에서는 대조군과 차이가 없었다. 통계분석 결과, 곰솔, 잣나무는 소나무재선충병에 높은 감수성을, 소나무, 섬잣나무는 중간 수준의 감수성을, 리기테다소나무와 스트로브잣나무는 확실한 저항성을 보여주는 것으로 나타났다. 본 연구는 소나무재선충 저항성 육종의 기초자료나 리기테다소나무와 스트로브잣나무의 조림 필요성에 대한 근거자료가 될 수 있을 것이다.
Abstract
Pine wilt disease is caused by the pine wood nematode (Bursaphelenchus xylophilus) and has killed many pine trees in Europe and Northeast Asia, including South Korea. Resistance to pine wilt disease varies among species. Previous studies were mostly conducted in nature or greenhouses and only a few in test fields. In this study, seedlings of six pine species (Pinus thunbergii, P. koraiensis, P. densiflora, P. parviflora, P. rigida × P. taeda, and P. strobus) were artificially inoculated by pine wood nematodes in the test field. The Wilt Index was measured every 2 weeks after inoculation in addition to the mortality rate, detection rate, and pine wood nematode concentration measurement after 24 weeks. The pine wilt disease mortality rates were P. thunbergii (80%), P. koraiensis (77.8%), P. densiflora (62.5%), and P. parviflora (22.0%), and both P. rigida × P. taeda and P. strobus survived. The pine nematode detection rates were the same among the species except for P. rigida × P. taeda pine (22.2%). High Wilt-Index values were obtained for P. thunbergii, P. koraiensis, and P. densiflora, which had mortality rates higher than the other species. Furthermore, there were no significant differences in the Wilt Indexes between P. parviflora, P. rigida × P. taeda, P. strobus, and the control group. Statistically, P. thunbergii and P. koraiensis showed high susceptibility to pine wilt disease, P. densiflora and P. parviflora showed moderate susceptibility, and P. rigida × P. taeda and P. strobus showed apparent resistance. These results provide basic data for pine wood nematode resistance breeding or as evidence of the need for afforestation of P. rigida × P. taeda and P. strobus.
Keywords: Pine wood nematode; Bursaphelenchus xylophilus; Pinus spp.; susceptibility to pine wilt disease; wilt index
서 론
소나무재선충병(Pine wilt disease) 또는 소나무시들음병은 소나무재선충(Bursaphelenchus xylophilus (Steiner & Buhrer, 1934))에 의해 감염된 소나무속(Pinus)의 수목이 수개월에 걸쳐 잎과 가지가 말라가며 고사에 이르는 병으로 동북아시아와 유럽 침엽수림에 피해를 주고 있다(Futai, 2013). 소나무재선충병의 매개충은 수염하늘소속(Monochamus)의 하늘소로 소나무재선충병에 의해 고사한 나무에 산란을 하며, 부화한 유충은 소나무재선충을 운반하는 성충이 되어 다른 나무로 전파한다(Mamiya and Enda, 1972). 소나무재선충은 원산지인 북아메리카에서 1934년에 처음 발견되었지만, 이 곳의 소나무속 수종은 대부분 저항성을 가지고 있어 문제가 되지 못했다(Steiner and Buhrer, 1934, Sutherland, 2008). 1905년, 일본에서 최초로 나가사키에서 소나무재선충병이 발견되었고, 1971년에 이 병의 원인이 소나무재선충이라는 사실이 밝혀졌다(Yano, 1913; Kiyohara and Tokushige, 1971). 1970년대에는 홋카이도를 제외한 일본 전역과 대만에 소나무재선충병이 확산되었고, 1980년대에는 중국과 한국까지 전파되었다(Yi et al., 1989; Futai, 2008; Zhao, 2008). 또한 유럽에서는 1999년에 포르투갈, 2008년에 스페인에서 발생하기 시작했다(Mota et al., 1999; EPPO, 2010).
한국은 1988년 부산에서 최초로 소나무재선충병이 발견되었다(Yi et al., 1989). 주로 남부지방의 곰솔(Pinus thunbergii Parl.)과 소나무(P. densiflora Siebold & Zucc.)에서 발병하였으며 매개충은 솔수염하늘소(Monochamus alternatus Hope) 1종이었다(Shin, 2008). 2006년부터는 새로운 매개충인 북방수염하늘소(M. saltuarius (Gebler))에 의해 중부지방의 잣나무(P. koraiensis Siebold & Zucc.)도 전파되기 시작했다(Han et al., 2008, Shin, 2008). 최근 5년간 한국에서 소나무재선충병으로 고사하는 나무는 매년 30만본 이상이며, 잠재적 발생 위험지역인 소나무, 곰솔, 잣나무림의 면적은 한국 산림에서 차지하는 비율은 약 40.1%에 달하기 때문에 소나무재선충병은 한국 산림의 가장 큰 위협 중 하나이다(KFS, 2022). 소나무재선충병 방제방법에는 Emamectin benzoate 등의 수간주사로 예방하거나, 감염된 피해목을 훈증하는 방법, 살충제를 항공살포하는 방법 등이 있다(Takai et al., 2000; Kamata, 2008). 보다 근본적인 해결을 위해, 일본은 1978년부터 저항성 육종을 시작했고, 중국, 한국, 포르투갈도 저항성을 가진 품종을 선발해내기 위한 연구를 진행중이다(Nose and Shiraishi, 2008; Carrasquinho et al., 2018; Woo et al., 2018; Gao et al., 2022).
소나무재선충병에 대한 수목들의 저항성은 수종에 따라 달라지며, 북아메리카가 원산인 수종은 주로 저항성을 가지지만 다른 곳이 원산지인 수종들은 대부분 감수성을 가진다. 한국에 자생하는 곰솔, 잣나무, 소나무, 섬잣나무(P. parviflora Siebold & Zucc.)는 감수성을 나타냈다(Mamiya, 1983; Koh et al., 2016; Health et al., 2019). 일본의 P. luchuensis Mayr, 중국의 P. tabulaeformis Carr., P. massoniana Lamb., 대만의 P. taiwanensis Hayata 등도 감수성을 가지고 있지만 백송(P. bungeana Zucc. ex Endl.)은 저항성을 가지는 것으로 보고되었다(Mamiya, 1983; Yang and Wang, 1989; Takeuchi, 2008). 유럽의 구주소나무(P. sylvestris L.)와 P. pinaster Aiton, P. nigra Arnold는 감수성을 보여주었으나 P. pinea L.은 저항성을 가지고 있다(Mamiya, 1983; Nunes da Silva et al., 2015). 북아메리카 원산인 스트로브잣나무(P. strobus L.), 리기다소나무(P. rigida Mill.), 테다소나무(P. taeda L.), 그리고 리기다소나무와 테다소나무의 교잡종인 리기테다소나무(P. rigida × P. taeda)는 저항성을 가지고 있었다(Mamiya, 1983). 하지만 같은 북아메리카 원산인 라디아타소나무(P. radiata D.Don)는 감수성을 지니고 있다(Mamiya, 1983; Nunes da Silva et al., 2015). 자연상태에서 소나무속 외에는 소나무재선충병의 발생이 매우 드물지만, 인공접종실험 결과 낙엽송(Larix kaempferi (Lam.) Carrière)은 감수성을 가지고 있었고(Mamiya and Shoji, 2009; Koh et al., 2016). 멕시코 삼나무(Cupressus lusitanica Mill.)는 저항성을 가지고 있었다(Nunes da Silva et al., 2013). 이외에도 다양한 종들에 대한 감수성 및 저항성 연구들이 이루어졌으나, 대부분 자연상태의 발병보고라서 형질에 대한 정량화가 어려우며, 인공접종을 통한 연구도 있었지만 주로 온실내에서의 어린 포트묘를 대상으로 했다는 점에서 한계를 가진다(Woo et al., 2008; Franco et al., 2011; Koh et al., 2016).
본 연구에서는 위와 같은 한계를 극복하기 위해 포지에서 생육 중인 소나무속 6종의 5~8년생 묘목에 인공접종 실험을 수행하였다. 한국에서 자생하는 곰솔, 잣나무, 소나무, 섬잣나무, 도입종인 스트로브잣나무, 그리고 도입종을 인공교잡한 리기테다소나무에 소나무재선충을 인공접종한 뒤 2주 간격으로 24주 후까지 관찰하여 감염율, 고사율, 소나무재선충 밀도, 위조지수(wilt index) 등을 측정하고 소나무재선충병에 대한 수종들의 감수성 및 저항성을 비교 분석하였다.
재료 및 방법
1. 공시재료
본 연구에 사용된 소나무속 6종은 곰솔, 잣나무, 소나무, 섬잣나무, 리기테다소나무, 스트로브잣나무로 5~8년생 묘목들이며 같은 종의 묘목들은 연령이 동일했다(Table 1). 묘목들은 종마다 다른 곳에서 양묘되다가 2021년 3월 30일과 4월 2일에 서울대학교 칠보산 학술림 포지로 이식되었다. 소나무 묘목은 국립품종관리센터의 안면도 채종원 종자를 2016년에 채집하여 칠보산 학술림 온실에서 2017년에 파종한 뒤 2019년에 포지로 이식되었다. 리기테다소나무 묘목은 국립산림과학원 산림생명자원연구부에 있는 리기테다소나무 성목로부터 종자를 받아 국립산림과학원 포지에서 양묘되었다. 나머지 4종은 양묘상으로부터 구입한 것이며, 그중 섬잣나무의 경우 곰솔을 대목으로 한 접목묘였다. 실험에 사용한 묘목들은 하나의 주간이 곧게 뻗어 올라왔으며 건강한 생육상태였다. 묘목들의 수고 및 근원경(Root collar diameter)은 2021년 7월 9일에 측정하였다(Table 1).
Table 1.
Growth characteristics of pine tree seedlings for the artificial inoculation test.
|
Pine species |
Age (years) |
Height (cm) |
Root collar diameter (mm) |
|
Pinus thunbergii
|
5 |
103.6±6.7a
|
18.93±2.11 |
|
Pinus koraiensis
|
5 |
84.5±10.3 |
23.32±3.06 |
|
Pinus densiflora
|
5 |
97.3±12.3 |
21.09±2.35 |
|
Pinus parviflora
|
7 |
94.0±14.9 |
35.60±5.17 |
|
Pinus rigida × P. taeda
|
8 |
73.8±13.6 |
22.55±3.17 |
|
Pinus strobus
|
5 |
62.1±9.8 |
20.15±3.96 |
Download Excel Table
2. 소나무재선충 배양
본 연구에 사용한 소나무재선충은 국립산림과학원에서 2021년에 분양 받아 계대배양하였다. 소나무재선충의 먹이원인 Botrytis cinereal Pers.를 먼저 배양하였으며 이를 위해 고형 PDA 배지(potato dextrose agar medium)를 만들었다. PDA 배지는 Difco™ Potato Dextrose Agar (BD, USA) 용액(39 g·l−1)을 멸균한 뒤 90 mm Petri dish(현대마이크로, 대한민국)에 약 35 ml씩 부은 다음 15시간 이상 자외선소독을 하면서 굳혔다. 완성된 고형 PDA배지에 B. cinerea를 접종한 다음 배양기에서 25°C, 암조건으로 7일간 배양했다. B. cinerea가 잘 자란 PDA 배지에 소나무재선충을 접종한 다음 배양기에서 25°C, 암조건으로 7일간 배양했다. 이후 소나무재선충은 4°C 냉장고에서 보관했으며 30일이 지나기 전에 새로운 B. cinerea가 자란 PDA 배지에 다시 계대배양하였다.
3. 소나무재선충 접종 및 경과 관찰
소나무속 6종에 대한 소나무재선충 접종은 2021년 6월 25일에 시행되었으며 Woo et al.(2018)의 방법을 참고하였다. 각 수종당 묘목 9그루(실험군)에 소나무재선충 1만마리가 들어있는 증류수 100 μl를 접종했으며, 다른 묘목 9그루에는 대조군으로 사용하기 위해 증류수 100 μl를 접종했다. 다만, 곰솔은 실험군이 10그루, 대조군이 8그루였다. 접종방법은 지상 10 cm 높이에서 묘목 수피를 세로로 잘라 틈을 만든 다음, 화장솜을 끼우고 화장솜과 형성층 사이로 액체를 주입했다. 접종 후에는 파라필름을 감싸서 소나무재선충이 밖으로 새지 않고 형성층으로 잘 들어갈 수 있게 하였다. 모든 소나무 묘목들은 접종일부터 2주 간격으로 사진을 촬영했으며 기온이 영하로 내려가 소나무재선충 증식이 더 이상 이뤄지지 않는 24주 후에 실험을 종료했다.
4. 묘목 내의 소나무재선충 검출
접종 24주 후 묘목에 있는 소나무재선충의 검출을 위해 Baermann funnel 법을 사용하였다(Baermann, 1917). 우선 묘목의 주간과 가지를 잘게 잘라(<5 mm) 24시간 후에 밑에 모인 용액을 20 ml 추출하였다. 그리고 슬라이드 글라스에 10 μl씩 옮겨서 실체현미경(Olympus SZ61, Tokyo, Japan)을 통해 10~20배율로 관찰하였다. 묘목 당 총 5번 관찰하여 소나무재선충의 검출여부를 확인하고, 검출이 된 경우 소나무재선충의 수를 측정했다. 그리고 Baermann funnel 법에 사용한 주간과 가지를 80°C에 7일간 건조한 뒤 건중량을 측정하여 소나무재선충의 밀도를 계산했다.
소나무재선충 접종 후 10주 이내에 고사한 묘목들은 즉시 Baermann funnel 법으로 소나무재선충 검출 여부를 판별하였다. 검출되지 않은 경우 소나무재선충 이외의 요인(이식 스트레스 등)에 의해 고사한 것으로 판단하고 실험 대상에서 제외하였다. 정상적으로 실험이 이뤄진 묘목들 중에서, 접종 24주 후 묘목의 고사 여부를 판단하여 고사율을 측정했다. 또한 소나무재선충을 접종한 묘목들은 Baermann funnel 법을 통해 소나무재선충 검출율 및 밀도를 측정했다. 그리고 증류수를 접종한 대조군에서도 종별로 1개 묘목에서 소나무재선충 검출 여부를 확인하였다.
5. 위조지수(wilt index) 측정
소나무재선충에 의한 피해 정도는 사진을 이용해 위조지수로 수량화 하였다(Figure 1). 위조지수는 모든 잎이 푸르고 증상이 없는 상태가 0, 25% 미만의 잎이 마르거나 갈변했을 때가 1, 25~50%의 잎이 마르고 갈변한 경우는 2, 50~75%의 잎이 마르거나 갈변한 경우는 3, 마지막으로 75% 이상의 잎이 마르고 갈변하거나 나무가 완전히 고사한 경우는 4로 등급화했다(Yu et al., 2012; He et al., 2016; Modesto et al., 2022a; Modesto et al., 2022b).
Figure 1.
Wilting process of a Pinus densiflora seedling inoculated by pine wood nematode. Wilt indexa of (A), (B), (C), (D) and (E) are 0, 1, 2, 3 and 4, respectively.
a 0 : absence of wilt or brown leaves, 1 : <25% of wilt or brown leaves, 2 : 25~50% of wilt or brown leaves, 3 : 50~75% of wilt or brown leaves, 4 : 75≤ wilt or brown leaves or death of the seedling.
Download Original Figure
6. 통계 분석
인공접종 24주 후의 소나무재선충 밀도, 검출율, 고사율 및 위조지수의 수종간 차이를 확인하기 위해 비모수 일원분산분석(One-way ANOVA)인 Kruskal–Wallis 검정과 Bonferroni correction 사후검정을 실시하였다. 통계분석은 R software v. 4.2.2 (R Core Team, 2022)으로 수행하였다.
결과 및 고찰
1. 접종 24주 후 소나무재선충 검출율 및 고사율
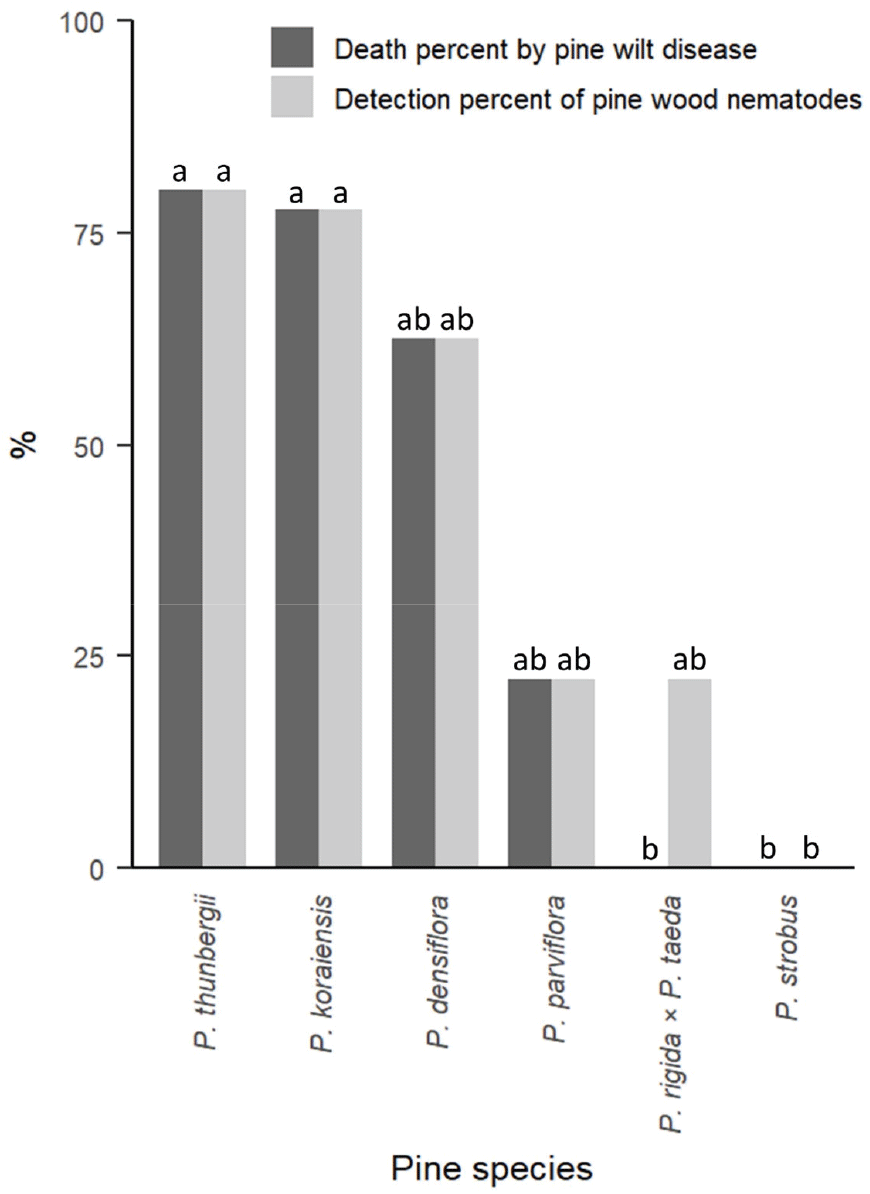
소나무재선충을 접종한 묘목들의 접종 24주 후 소나무재선충 검출률은 Table 2와 같으며, 고사율과 함께 막대그래프로 시각화 하였다(Figure 2). 소나무재선충을 접종한 묘목들 중, 조기에 고사한 소나무 1그루(8주차)와 스트로브잣나무 1그루(6주차)는 소나무재선충이 검출되지 않았으며, 이식 스트레스로 인한 고사로 판단하여 통계에서 제외하였다. 소나무재선충의 검출율은 수종 간에 통계적으로 유의한 차이를 보였다. 곰솔(80.0%)과 잣나무(77.8%)가 스트로브잣나무(0%) 보다 소나무재선충 검출율이 유의하게 높았으며, 중간 수치를 보여준 소나무(62.5%), 섬잣나무(22.2%), 리기테다소나무(22.2%)는 다른 수종과 유의한 차이를 나타내지 않았다. 증류수를 접종한 묘목에서 수종별로 1그루씩 선정하여 소나무재선충을 검출해본 결과 6그루 모두 검출되지 않았다.
Table 2.
Resistant characteristics of six pine species in 24 weeks post-inoculation with pine wood nematode.
|
Pine species |
n
c
|
Detection percent of PWNd (%) |
Death percent (%) |
PWN densitye
|
Average wilt indexf
|
|
Pinus thunbergii
|
10 |
80.0a
|
80.0a
|
105.05a
|
3.5a
|
|
Pinus koraiensis
|
9 |
77.8a
|
77.8a
|
153.65a
|
3.2ab
|
|
Pinus densiflora
|
8 |
62.5ab
|
62.5ab
|
36.04ab
|
2.9ab
|
|
Pinus parviflora
|
9 |
22.2ab
|
22.2ab
|
83.39ab
|
1.7ab
|
|
Pinus rigida × P. taeda
|
9 |
22.2ab
|
0b
|
0.27ab
|
0.9b
|
|
Pinus strobus
|
8 |
0b
|
0b
|
0b
|
1.0b
|
|
Mean |
8.8 |
44.1 |
40.4 |
54.67 |
1.94 |
|
Kruskal-Wallis χ2
|
- |
19.761**
|
25.385***
|
20.664***
|
23.894***
|
Download Excel Table
Figure 2.
Death percent by pine wilt disease and detection percent of pine wood nematodes of six pine species in 24 weeks post-inoculation with pine wood nematode.
Download Original Figure
소나무재선충으로 인한 고사율의 경우 리기테다소나무를 제외하면 모두 고사율과 동일하였으며, 고사한 묘목들은 소나무재선충이 검출된 묘목들과 일치했다. 리기테다소나무는 소나무재선충이 검출된 묘목 2그루 모두 생존하여 고사율이 0%였다. 통계 분석 결과 고사율도 수종에 따라 유의한 차이를 보였는데, 곰솔(80.0%)과 잣나무(77.8%)가 리기테다소나무(0%)와 스트로브잣나무(0%) 보다 유의하게 높았다. 중간 수치를 기록한 소나무(62.5%), 섬잣나무(22.2%)는 다른 수종과 유의한 차이를 나타내지 않았다. 증류수를 접종한 대조군의 경우 섬잣나무 1그루(10주차), 잣나무 2그루(6주차), 스트로브잣나무 1그루(4주차)를 제외하고는 모두 생존하였다. 조기에 고사한 묘목 4그루 모두 소나무재선충이 검출되지 않았으며, 이식에 따른 스트레스로 인하여 고사한 것으로 판단된다.
곰솔, 잣나무는 통계적으로 유의미하게 높은 소나무재선충병 감수성을 보인다고 할 수 있으며 이는 기존 연구결과와 동일하다(Mamiya, 1983). 소나무는 62.5%의 고사율로 곰솔과 잣나무보다 낮았으며, 곰솔이 소나무보다 감수성이 높다는 연구결과(Woo et al., 2020)와도 부합하지만, 통계적으로 다른 수종과 유의한 차이를 보이지는 못했다
섬잣나무는 검출률과 고사율이 22.2%로 상당히 낮은 수치를 보여주었지만 통계적으로 다른 수종과 유의한 차이를 보이지 못했다. 섬잣나무는 한국에서는 울릉도에서만 자생하고 조경용으로 일부 식재되어 감염보고가 드물었으나, 본 연구에서처럼 소나무재선충병으로 인하여 고사한다는 연구결과가 일부 있었기 때문에 섬잣나무에 대한 소나무재선충 방제에도 주의를 기울여야 할 것이다(Takeuchi, 2008; Koh et al., 2016).
리기테다소나무와 스트로브잣나무는 고사율이 0%로서 기존 연구결과처럼 강한 저항성을 보였다고 해석된다(Mamiya, 1983). 리기테다소나무의 소나무재선충 검출률은 22.2%였지만, 24주 후까지 고사는 없었으며 잘 생육하였으므로 소나무재선충병에 걸리지 않았다고 볼 수 있다. 종합해보면 곰솔, 잣나무는 강한 감수성을, 소나무와 섬잣나무는 중간 수준의 감수성을, 리기테다소나무와 스트로브잣나무는 저항성을 보여주었다.
2. 접종 24주 후 소나무재선충 밀도
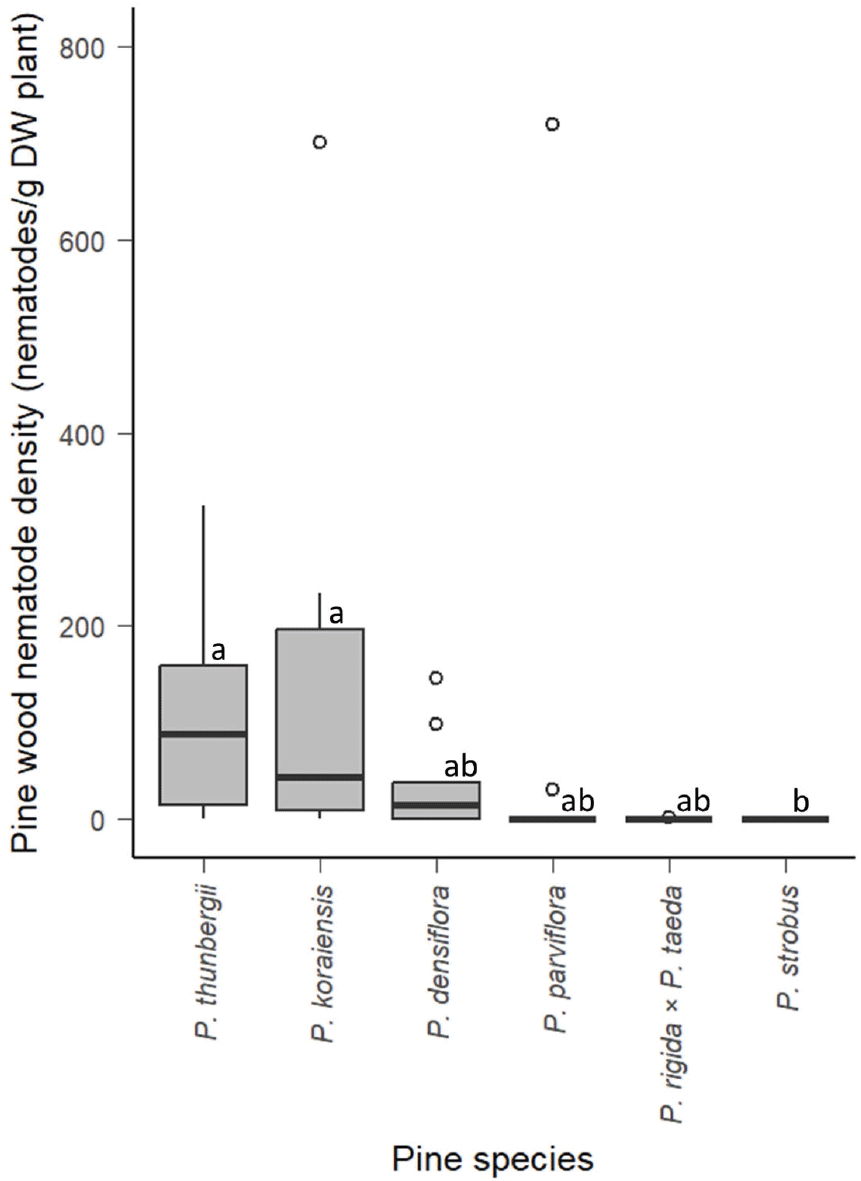
접종 24주 후 묘목에서 추출된 소나무재선충의 평균 밀도는 Table 2에 명시했으며, Figure 3에 상자 수염 그림(Box-and-Whisker Plot)으로 나타냈다. 소나무재선충 평균 밀도는 6종 중에서 잣나무가 153.65 마리·g−1으로 가장 높았다. 그리고 내림차순으로 곰솔(105.05 마리·g−1),섬잣나무(83.39 마리·g−1), 소나무(36.04 마리·g−1) 순이였다. 리기테다소나무(0.27 마리·g−1)는 소나무재선충이 극소량이 발견되었고, 스트로브잣나무는 모든 묘목에서 검출되지 않았다. 통계 분석 결과 곰솔과 잣나무가 스트로브잣나무보다 유의하게 높았으며, 다른 수종은 유의한 차이를 보이지 않았다.
Figure 3.
Pine wood nematode density of six pine species in 24 weeks post-inoculation with pine wood nematode.
a,b Grouping by Bonferroni correction test at α = 0.05.
Download Original Figure
소나무재선충 밀도는 소나무재선충의 검출률 및 고사율과 크게는 유사한 경향을 보였지만, 자세히 들여다보면 어느 정도 차이를 보였다. 우선 검출률과 고사율 분석에서 감수성을 나타내는 곰솔, 잣나무, 소나무, 섬잣나무의 소나무재선충 평균 밀도가 모두 30 마리·g−1 이상이었으며 이들 수종에서 소나무재선충이 활발히 번식했음을 보여준다. 반면 검출률과 고사율 분석에서 저항성을 보여준 리기테다소나무는 평균 0.27 마리·g−1에 그쳤으며 스트로브잣나무는 모두 검출되지 않았다. 스트로브잣나무의 경우 접종 후에 소나무재선충이 번식에 실패한 것으로 보이며, 리기테다소나무의 경우 소나무재선충이 번식은 했지만, 병을 일으킬 만큼 증식하는 데는 실패하고 소수 만이 생존에 성공한 것으로 보이며 선행연구에서도 비슷한 결과가 나타난 바 있다(Woo et al., 2018).
감수성을 나타내는 수종의 경우, 곰솔 보다 약간 낮은 고사율을 보인 잣나무에서 오히려 1.5배 정도 높은 소나무재선충 평균 밀도가 나타났고, 다른 3개 수종보다 확연히 낮은 고사율을 보여준 섬잣나무의 소나무재선충 평균 밀도는 소나무보다 2배 이상 높았다. 소나무재선충의 평균밀도는 검사하는 부위나 시기에 따라 달라질 수 있으며, 일정 수 이상보다 높아지면 소나무재선충병이 발병한다고 볼 수 있기 때문에 본 연구의 밀도 분석을 자세히 하기에는 한계가 있었다.
3. 소나무재선충 접종 후 위조지수의 변화 및 고사율
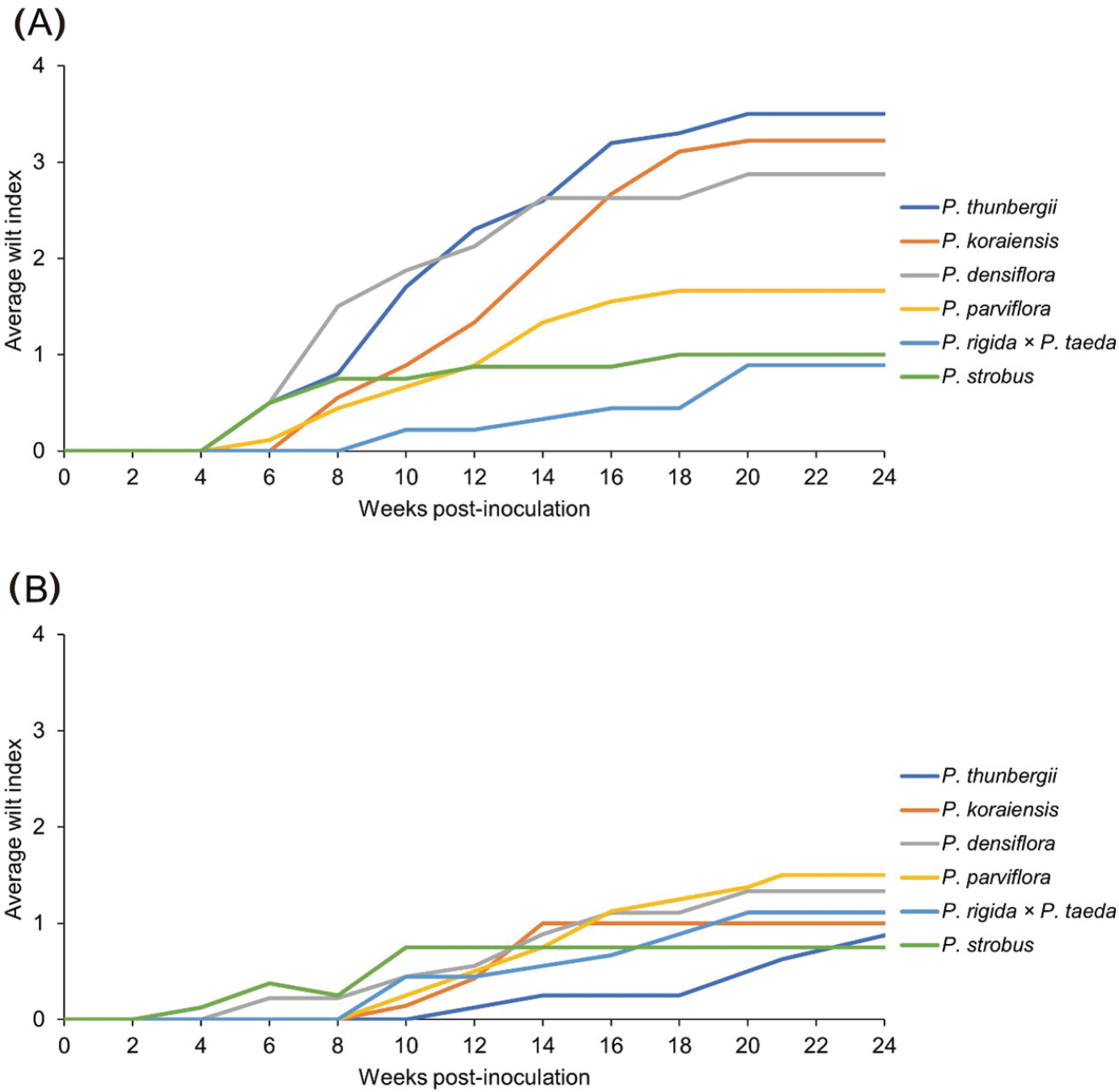
소나무재선충 접종에 사용한 묘목들의 위조지수는 처음에는 모두 0이였으며, 접종 후 4주까지 그대로 유지되었다[Figure 4(A)]. 곰솔, 소나무, 스트로브잣나무, 섬잣나무의 위조지수가 6주차부터 증가하기 시작했으며, 잣나무는 8주차, 리기테다소나무는 10주차부터 증가하였다. 곰솔과 소나무는 12주차에 평균 위조지수가 2를 넘었고, 잣나무는 14주차에 2에 도달했다. 곰솔은 12주차부터 계속 1위를 기록했으며, 16주차에 평균 위조지수가 3을 돌파했다. 잣나무는 16주차부터 소나무보다 평균 위조지수가 커지기 시작했고, 18주차에 3을 돌파했다. 소나무는 평균 위조지수가 2.9로 3을 돌파하지는 못했으며, 섬잣나무는 18주차부터 1.7을 유지했다. 리기테다소나무와 스트로브잣나무는 1 이하를 유지했다. 이후 20주차부터 24주차까지 모든 수종에서 위조지수가 그대로 유지되었다. 24주차에서 가장 높은 위조지수를 보인 것은 곰솔(3.5)이였으며, 잣나무(3.2), 소나무(2.9), 섬잣나무(1.7), 스트로브잣나무(1.0), 리기테다소나무(0.9) 순으로 감소하였다(Table 2). 통계 분석 결과 곰솔은 리기테다소나무와 스트보르잣나무 보다 유의하게 높은 수치를 보여주었으며 다른 수종에서는 유의한 차이를 보여주지 못했다.
Figure 4.
Pine wilt disease symptoms progression expressed in average wilt indexa for every 2 weeks post-inoculation with pine wood nematode (A) and distilled water (B).
a 0 : absence of wilt or brown leaves, 1 : <25% of wilt or brown leaves, 2 : 25~50% of wilt or brown leaves, 3 : 50~75% of wilt or brown leaves, 4 : 75≤ wilt or brown leaves or death of the seedling.
Download Original Figure
증류수를 접종한 대조군 묘목들의 위조지수들은 처음의 0에서 크게 증가하지는 않았지만, 24주차의 값이 1.5~0.8 사이의 값으로 증가하였다[Figure 4(B)]. 이들 묘목들은 일부 잎이 마르거나 변색되었지만 전체적인 생육상태는 양호했으며, 이식으로 인한 스트레스나 겨울 휴지기를 대비해 엽록소가 제거되는 과정이라고 생각된다.
실험군과 대조군의 24주차 평균 위조지수를 비교해보면, 검출률과 고사율이 62.5% 이상이었던 곰솔(0.9)이나 잣나무(1.0), 소나무(1.3)는 실험군과 확연히 차이가 났지만, 22.2% 또는 0%를 보여준 섬잣나무(1.5), 스트로브잣나무(1.1), 리기테가소나무(0.8)의 경우 실험군과 큰 차이를 보이지 않았다. 그러나 강한 감수성을 지닌 수종이라도 소나무재선충병의 병징 발현이 느려져서 위조지수가 증가하지 않거나, 다른 원인으로 인해 잎마름이 발생하여 위조지수가 증가할 수 있기 때문에 위조지수만으로 소나무재선충병을 판단할 수는 없을 것이며 현장에서의 보조적인 지표로 활용할 수 있을 것이다.
결 론
본 연구에서는 소나무속 6종에 대해서 인공접종 실험을 통해 소나무재선충병에 대한 저항성을 비교하였다. 한국에서 자생하는 소나무, 곰솔, 잣나무, 섬잣나무는 감수성을 보였고, 도입종들의 교잡종인 리기테다소나무와 도입종인 스트로브잣나무는 강한 저항성을 보였다. 감수성을 보인 4개 수종에서 통계적으로 곰솔, 잣나무는 높은 감수성을 보여주었으며 소나무와 섬잣나무는 중간 수준의 감수성을 보여주었다. 리기테다소나무와 스트로브잣나무는 전혀 고사하지 않았으며, 리기테다소나무의 경우 낮은 밀도의 소나무재선충이 형성층에 존재하는 경우가 일부 있었다. 이러한 실험 결과는 이전 실험 결과들과 대부분 부합하였으며, 포지에서 5년생 이상의 묘목을 대상으로 6종을 동시에 인공접종을 하는 비교실험을 통해 다시 검증했다는 점에서 그 의의를 지닌다고 할 수 있겠다. 향후 이들 감수성 수종과 저항성 수종에 대한 유전연구를 통해 소나무재선충 저항성과 관여하는 유전자에 대한 탐색이 가능할 것이며, 장기적으로는 저항성 육종에 활용할 수 있을 것이다. 또한 스트로브잣나무와 리기테다소나무의 저항성을 재확인함으로써 소나무재선충에 대응할 수 있는 조림대상 수종으로서 근거자료로 사용할 수 있을 것이다. 다만 신규조림 대상 수종 지정을 위해서는 스트로브잣나무와 리기테다소나무 목재의 경제적 가치 및 다른 병해충 취약성 등을 고려해야 할 것이며, 기존 조림수종인 소나무나 곰솔, 잣나무의 경제적 가치 및 공익적 가치와 비교하여 신중히 선정해야 할 것이다. 마지막으로 본 연구에 사용한 수종의 수나 반복수에는 한계가 존재하기 때문에 추후 연구에서 수종의 수와 반복수를 늘린다면 정확도를 높이고 연구의 가치 및 활용도를 높일 수 있을 것이다.
감사의 글
본 연구는 산림청(한국임업진흥원) 교란생태계 적응 및 복원 기술개발 사업(과제번호: 2020185D10-2022-AA02)의 지원에 의하여 이루어진 것입니다.
References
Baermann, G. 1917. Eine einfache methode zur auffindung von Ancylostomum (Nematoden) larven in erdproben. Geneeskd Tijdschr Ned Indie 57: 131-137.

Carrasquinho, I., Lisboa, A., Inácio, M.L. and Gonçalves, E. 2018. Genetic variation in susceptibility to pine wilt disease of maritime pine (
Pinus pinaster Aiton) half-sib families. Annals of Forest Science 75(3): 85.

European and Mediterranean Plant Protection Organization (EPPO). 2010. Isolated finding of
Bursaphelenchus xylophilus in Spain. EPPO Reporting Service 3.
Franco, A.R., Santos, C., Roriz, M., Rodrigues, R., Lima, M.R.M. and Vasconcelos, M.W. 2011. Study of symptoms and gene expression in four
Pinus species after pinewood neematode infection. Plant Genetic Resources 9(2): 272-275.
Futai, K. 2008. Pine wilt in Japan: from first incidence to the present. pp. 5-12. In: B.G. Zhao, et al. (Ed.). Pine wilt disease. Springer. Tokyo, Japan.
Futai, K. 2013. Pine wood nematode,
Bursaphelenchus xylophilus. Annual Review of Phytopathology 51(1): 61-83.
,

Gao, J., Pan, T., Chen, X., Wei, Q. and Xu, L. 2022. Proteomic analysis of Masson pine with high resistance to pine wood nematodes. Plos one 17(8): e0273010.
,
,

Han, H., Chung, Y.-J. and Shin, S.-C. 2008. First report of pine wilt disease on
Pinus koraiensis in Korea. Plant Disease 92(8): 1251-1251.
,
,
He, L.X., Wu, X.Q., Xue, Q. and Qiu, X.W. 2016. Effects of endobacterium (
Stenotrophomonas maltophilia) on pathogenesis-related gene expression of pine wood nematode (
Bursaphelenchus xylophilus) and pine wilt disease. International Journal of Molecular Sciences 17(6): 778.
,
,
Health, E.Panel o.P., et al. 2019. Commodity risk assessment of black pine (
Pinus thunbergii Parl.) bonsai from Japan. EFSA Journal 17(5): e05667.
,
Kamata, N. 2008. Integrated pest management of pine wilt disease in Japan: tactics and strategies. pp. 304-322. In: B.G. Zhao, et al. (Ed.). Pine wilt disease. Springer. Toyko, Japan.
Korea Forest Service (KFS). 2022 Statistical yearbook of forestry No. 52. Korea Forest Service: Seoul, Republic of Korea. pp. 455.
Kiyohara, T. and Tokushige, Y. 1971. Inoculation experiments of a nematode,
Bursaphelenchus sp., onto pine trees. Journal of the Japanese Forestry Society 53(7): 210-218.
Koh, S.H., Kwon, T.S., Nam, Y., Seo, S.T., Jung, C., Choi, K.S., Choi, K.S. and Han, H. 2016. Analysis of damage pattern from pine wilt disease by tree species and improvement of control technology. National Institute of Forest Science: Seoul. pp. 168.
Mamiya, Y. 1983. Pathology of the pine wilt disease caused by
Bursaphelenchus xylophilus. Annual Review of Phytopathology 21(1): 201-220.
,
Mamiya, Y. and Enda, N. 1972. Transmission of
Bursaphelenchus lignicolus (nematoda: Aphelenchoididae) by
Monochamus alternatus (coleoptera: Cerambycidae). Nematologica 18(2): 159-162.
Mamiya, Y. and Shoji, T. 2009. Pathogenicity of the pinewood nematode,
Bursaphelenchus xylophilus, to Japanese larch,
Larix kaempferi, seedlings. Journal of Nematology 41(2): 157-62.
Modesto, I., Inácio, V., Novikova, P., Carrasquinho, I., Van de Peer, Y. and Miguel, C.M. 2022a. SNP detection in
Pinus pinaster transcriptome and association with resistance to pinewood Nematode. Forests 13(6): 946.
Modesto, I., Inácio, V., Van de Peer, Y. and Miguel, C.M. 2022b. MicroRNA-mediated post-transcriptional regulation of
Pinus pinaster response and resistance to pinewood nematode. Scientific Reports 12: 5160.
,
,
Mota, M.M., Braasch, H., Bravo, M.A., Penas, A.C., Burgermeister, W., Metge, K. and Sousa, E. 1999. First report of
Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1(7): 727-734.
Nose, M. and Shiraishi, S. 2008. Breeding for resistance to pine wilt disease. pp. 334-350. In: B.G. Zhao, et al. (Ed.). Pine wilt disease. Springer. Tokyo, Japan.
Nunes da Silva, M., Lima, M.R.M. and Vasconcelos, M.W. 2013. Susceptibility evaluation of
Picea abies and
Cupressus lusitanica to the pine wood nematode (
Bursaphelenchus xylophilus). Plant Pathology 62(6): 1398-1406.
Nunes da Silva, M., Solla, A., Sampedro, L., Zas, R. and Vasconcelos, M.W. 2015. Susceptibility to the pinewood nematode (PWN) of four pine species involved in potential range expansion across Europe. Tree Physiology 35(9): 987-999.
,
Shin, S.C. 2008. Pine wilt disease in Korea. pp. 26-32. In: B.G. Zhao, et al. (Ed.). Pine wilt disease. Springer. Tokyo, Japan.
,
Steiner, G.M. and Buhrer, E.M. 1934.
Aphelenchoides xylophilus n. sp., a nematode associated with blue-stain and other fungi in timber. Journal of Agricultural Research 48: 949-951.
Sutherland, J.R. 2008. A brief overview of the pine wood nematode and pine wilt disease in Canada and the United States. pp. 13-17. In: B.G. Zhao, et al. (Ed.). Pine wilt disease. Springer. Tokyo, Japan.
Takai, K., Soejima, T., Suzuki, T. and Kawazu, K. 2000. Emamectin benzoate as a candidate for a trunk-injection agent against the pine wood nematode,
Bursaphelenchus xylophilus. Pest Management Science 56: 937-941.
Takeuchi, Y. 2008. Host fate following infection by the pine wood nematode. pp. 235-249. In: B.G. Zhao, et al. (Ed.). Pine wilt disease. Springer. Tokyo, Japan.
Woo, K.S., Lee, D.H., Koo, Y.B. and Yeo, J.K. 2008. Inoculation of seven pine species or hybrid seedlings with Korean isolates of pinewood nematode under greenhouse conditions. Annals of Forest Science 65(8): 1.
Woo, K.S., Shim, D., Il, H.L., Kim, J. and Kim, I.S. 2018. Selection of resistent pine tree against pine wilt disease through artificial inoculation. National Institute of Forest Science. pp. 90.
Woo, K.S., Shim, D., Na, S.J., Kim, J. and Lee, I.H. 2020. Breeding for resistance to pathogenic pine wood nematodes. pp. 166. In : C. D. Nelson et al. (Ed.). Proceedings of the sixth International workshop on the genetics of host-parasite interactions in forestry tree resistance to insects and diseases: putting promise into practice. Department of Agriculture Forest Service, Southern Research Station. Asheville, USA.
Yang, B. and Wang, Q. 1989. Distribution of the pinewood nematode in China and susceptibility of some Chinese and exotic pines to the nematode. Canadian Journal of Forest Research 19(12): 1527-1530.
Yano, S. 1913. Investigation on pine death in Nagasaki prefecture (in Japanese). Sanrin-Kouhou 4: 1-14.
Yi, C.K., Byun, B.H., Park, J.D., Yang, S.I. and Chang, K.H. 1989. First finding of the pine wood nematode,
Bursaphelenchus xylophilus (steiner et buhrer) nickle and its insect vector in Korea. The Research Reports of the Forestry Research Institute 3: 141-149.
Yu, L.Z., Wu, X.Q., Ye, J.R., Zhang, S.N. and Wang, C. 2012. NOS-like-mediated nitric oxide is involved in
Pinus thunbergii response to the invasion of
Bursaphelenchus xylophilus. Plant Cell Rep 31(10): 1813-21.
,
Zhao, B.G. 2008. Pine wilt disease in China. pp. 18-25. In: B.G. Zhao, et al. (Ed.). Pine wilt disease. Springer. Tokyo, Japan.
,