Article
낙엽송 클론 채종원에서 구과 채취시기에 따른 구과특성 및 종자품질
김예지
1, 구다은
1,2, 조계홍
1,3, 최희윤
1, 우영곤
1,2, 이채빈
1, 유성열
3, 주혜준
3, 강규석
1,*
Cone Characteristics and Seed Quality among Harvest Times in the Clonal Seed Orchard of Larix kaempferi
Ye-Ji Kim
1, Da-Eun Gu
1,2, Gyehong Cho
1,3, Heeyoon Choi
1, Yeongkon Woo
1,2, Chae-Bin Lee
1, Sungryul Ryu
3, Hye-Joon Joo
3, Kyu-Suk Kang
1,*
1Department of Agriculture, Forestry and Bioresources, Seoul National University, Seoul 08826, Korea
2Department of Forest Bioresources, National Institute of Forest Science, Suwon 16631, Korea
3Department of Seed and Seedling Management, National Forest Seed Variety Center, Chungju 27495, Korea
© Copyright 2023 Korean Society of Forest Science. This is an Open-Access article distributed under the terms of the
Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits
unrestricted non-commercial use, distribution, and reproduction in any
medium, provided the original work is properly cited.
Received: Jul 06, 2023; Revised: Aug 21, 2023; Accepted: Aug 22, 2023
Published Online: Sep 30, 2023
요 약
일본잎갈나무는 종자 생산의 풍흉 주기가 길고 불규칙하여 채종원산 우량 종자의 안정적인 대량생산을 위해 종자생산 효율을 높이는 계획 수립이 필요하다. 구과의 채취시기는 구과특성과 종자 품질을 결정하는 중요한 요소로 종자 뿐만 아니라 유묘 생산까지도 영향을 준다. 충주 일본잎갈나무 채종원에서 구과의 적정한 채취시기를 판단하고 채취시기별 구과 및 종자 특성을 분석하기 위해 2021-2022년도에 채취시기별로 구과 함수율, 종자 효율(생산량), 발아율, 발아세 등을 조사하여 다변량 분석을 수행하였다. 8월 초에 구과 함수율의 감소와 구과의 갈변이 이루어졌고, 발아율은 점차 감소하는 경향이 있었으나 9월말에 최고점이 나타나 경향성이 뚜렷하지 않았다. 구과 채취시기가 늦을수록 발아속도는 우수하였으나, 종자 비산, 충해 종자의 증가로 종자 효율은 감소하였다. 따라서 발아율과 종자 생산량을 최대로 하는 구과의 최적 채취시기는 8월 초일 것으로 판단되었다. 다만 균일한 묘목 생산에 기여하는 발아속도를 고려한다면 시기적절한 병해충 방제를 통해 종자손실을 절감시킬 수 있는 9월 초에 구과를 채취해야 할 것이다. 또한 일본잎갈나무 채종원에서 구과의 갈변 정도와 함수율은 구과 채취시기의 결정 지표로 활용할 수 있는 것으로 나타났다.
Abstract
Harvest time is one of the most important determining factors of seed quality, especially for species that produce seeds over irregular and long-term periods, such as Larix kaempferi. A cone collection plan must be established to increase seed production efficiency and stable mass production. We investigated seed qualities such as seed efficiency, germination rate, and T50 (germination speed), with 7 or 8 cone collection times at a clonal seed orchard of L. kaempferi in Chungju between 2021 and 2022. A multivariate analysis was then performed for the collected data. In early August, decreases in the moisture contents and browning of cones were observed. These were followed by a decrease in germination rate, with a peak at the end of September, but no clear trend was observed. The later the cones were harvested, the better the seed vigor (T50). However, the seed yield and efficiency decreased owing to increases in seed scattering and the number of insect-damaged seeds. As a result, the optimal time of seed harvest for the seed orchard was in early August. To produce uniform seedlings, insect damage must be reduced through timely control and harvest cones in early September. This shows that the degree of browning and moisture content of cones can be used as indicators of the timing of cone collection in L. kaempferi seed orchards.
Keywords: cone collection; Larch; optimal collection time; seed orchard; seed quality
서 론
채종원은 임목의 유전적 형질 개량을 목적으로 하는 육종 프로그램에서 개량된 조림 재료를 대량 생산하는 시스템이다(Sweet, 1995). 낙엽송을 포함한 주요 조림용 수종은 일반적으로 자연 임분에서 생장이 우수하고 수형이 곧은 개체가 수형목으로 지정되며, 이후 유전적으로 우수한 형질을 가졌음을 검정하고 유·무성 증식된 채종목으로 채종원이 조성된다(Yuan et al., 2016). 채종목은 생산될 종자가 부모 세대의 유전적 다양성을 유지하고 클론 또는 가계 간의 근친교배가 최소화되도록 배치되며(Lindgren et al., 2009; Lstibůrek and El-Kassaby, 2010), 채종원은 외부 화분에 의한 유전자의 이입을 방지할 수 있도록 조성된다(Adams et al., 1997; Torimaru et al., 2009).
채종원에서 생산된 고품질 종자는 자연 임분에서 채취한 종자보다 더 빠르고 균일하게 자라는 반면 조성에서부터 종자공급까지 많은 시간과 비용이 소요된다. 종자의 채취를 위해 매년 많은 인력이 필요하며, 높은 가지에 있는 구과를 채취하거나 이를 위해 가지를 자르는 것은 작업자의 안전과 후년의 종자 생산량에 영향을 미치게 된다. 따라서 과학적인 데이터에 근거한 적절한 종자채취 시기, 방법 등의 계획 수립을 통해 현장에서 발생하는 많은 비용과 노동력을 절감할 수 있다. 현재까지의 관련 연구는 환상박피, 호르몬 또는 광 처리 등(Crain and Cregg, 2018; Matsushita et al., 2020) 종자 생산량을 증가시키는 연구에 집중 되어있으며 이러한 관리기술과 처리를 통해 종자 생산량을 증가시키는 방안은 또다른 비용이 발생한다. 결국 생산될 종자의 품질을 높이기 위해 종자의 활력이 가장 높은 시기에 맞추어 구과를 채취하는 연간 계획을 세우는 것이 매우 중요하다.
일본잎갈나무(L. kaempferi)는 유시 생장이 빠른 침엽수로 내구성이 높고 통직하여 목재로서 가치가 높아 국내에서 조림 수요가 가장 많은 수종이다(Korea Forest Service, 2022). 일본잎갈나무(Larix kaempferi (Lamb.) Carrière) 채종원은 접목묘로 조성되었다. 접목묘는 실생묘보다 개화가 빠르고 약 10년생의 채종목에서 수구화수의 개화가, 15년생부터 암구화수의 개화가 시작된다(Lee et al., 2012). 일본잎갈나무의 화아원기 형성에 대한 연구는 아직 보고된 바 없으나, 당년지의 신초 생장이 멈추는 시기가 6월 중순으로 서양낙엽송(L. occidentalis Nutt.)과 유사하여 같은 속인 일본잎갈나무 역시 7월부터 이듬해 화아원기가 생성된다고 추정된다(Owens and Molder, 1979; Owens and Blake, 1985). 이 때의 기온, 일조량이 일본잎갈나무의 화아 형성에 영향을 미친다고 일반적으로 알려져 있다(Lee et al., 2012). 다만, 화아 분열부터 종자 결실까지 1년이 소요되며 그 사이에 휴면, 생장, 개화, 수분, 수정과 같은 생리적 변화를 거치게 된다. 따라서 최종 종자 생산에 미치는 요인은 다양한 환경적, 유전적 요인에 의해 매우 복합적으로 발생하며 개체목마다 각 생리적 변화를 겪는 시기가 달라져 임목에서는 장기간의 모니터링을 필요로 한다(Mou and Kronstrad, 1994; Kang and Lindgren, 1999; Kim et al., 2008). 특히 일본잎갈나무는 목재 생산을 위한 조림 수요가 높지만 결실 풍흉 주기가 길고 불규칙하여 안정적인 종자 수급에 어려움을 겪고 있는 수종이다. 따라서 종자 생산의 효율을 높이는 근본적인 계획 수립이 필요하다.
개화 이후 수분과 수정을 거친 종자의 성숙 단계는 크게 3단계로 구분된다. 첫째, 활발한 세포 분열로 종자의 구조가 생성되는 단계, 둘째, 물질을 축적하며 건물중이 크게 증가하는 단계, 마지막으로 서서히 종자 생장이 멈추는 단계로 나눌 수 있다(Egli, 2017). 건물중이 점점 증가하며 영양분, 호르몬 등이 축적되고 합성되는 시기를 종자생장기(seed filling period)라 하며, 건물중이 최대가 되고 종자생장이 멈추었을 때를 생리적 성숙(physiological maturity)이 되었다고 표현한다(Shaw and Loomis, 1950). 이 때가 종자 품질이 최대가 되고 이후에는 모수로부터 영양 공급이 중단되어 종자 품질이 점차 낮아진다고 주장하는 가설(Harrington and Kozlowski, 1972)이 지배적이다. 반면에 최대 건물중에 도달한 이후에도 종자 품질이 향상된 경우가 다수 보고되며, 생리적 성숙 단계 이후에도 건조 저항성, 수명 등과 관련된 성숙이 이루어진다는 보고도 있다(Pieta and Ellis, 1991; Zanakis et al., 1994; Hay et al, 2010; Leprince et al., 2017). 따라서 생리적 성숙 시기를 대신할 용어로 대량 성숙 시기(mass maturity)를 사용하기도 했으며(Ellis and Pieta, 1992), 종자의 최대 건물중이 반드시 최대 품질을 의미하는 것은 아니다. 즉, 최종적으로 수확 성숙(harvest maturity)에 도달하는 시기는 수종마다 다르며(Elias and Copeland, 2001; Bedane et al., 2006), 유전인자와 환경조건의 상호작용에 의해 종자 품질의 향상과 퇴화가 나타난다(Ellis, 2019; Gu et al., 2017).
종자가 적정한 수확 성숙기에 도달한 이후에도 구과 가해 병해충 피해에 의해 종자 생산량이 감소할 수 있으므로 집약적인 종자 생산을 위한 채종원에서는 해충 방제 역시 필수적이다(Grosman et al., 2002; Rosenberg and Weslien, 2005). 즉 너무 이른 시기에 구과를 채취하면 미성숙으로 인해 품질이 낮아 종자의 수확량이 떨어지며(Kelly and George, 1998), 지나치게 늦은 채취는 과도한 건조, 퇴화에 따라 종자의 품질을 떨어뜨리고(Likoswe et al., 2008) 구과 충해로 인해 종자 생산량이 감소할 수 있다. 따라서 낙엽송 채종원에서 적절한 구과 채취시기란 종자의 생리적 발달 단계가 고려되어 종자 생산량을 증대시킴과 동시에 종자의 품질이 최대화될 수 있는 시기이다.
종자의 함수율은 발달 초기단계에서는 모수와 평형을 이루는 상태를 유지하다가 생리적 성숙 이후 서서히 감소하기 시작한다. 함수율이 일정 이하로 떨어지면 대사가 중지되고 퇴화가 시작되므로(Bewley et al., 2013), 채취 직후의 함수율은 종자의 품질을 간접적으로 판단할 수 있는 기준이 되고(Probert et al., 2007) 종자 품질이 최대가 되는 적정 함수율은 수종마다 다르다. 종자 품질을 평가하기 위해서는 주로 종자 발아능력(seed viability)과 종자 활력(seed vigor)을 함께 사용한다. 종자 발아능력이란 적절한 환경에서 종자가 발아하여 유묘를 생산할 수 있는 능력으로 국제종자검정협회(ISTA)에서 규정한 표준 발아검사에 의해 측정된 발아율을 통해 평가될 수 있다. 종자 활력은 광범위한 환경에서 빠르고 균일하게 발아하여 정상묘를 생산할 수 있는 능력을 말하며, 전기전도도, 노화 처리, 발아 속도 등을 통해 검사할 수 있다. 양묘 관리에서 파종 후 일시에 많은 종자가 발아되는 것이 중요하므로 채종원에서의 종자 품질은 균일한 묘목 생장과 산림 생산성 증대에 관여한다.
본 연구에서는 충주 일본잎갈나무 채종원에서 구과 채취시기 별로 종자 생산량과 활력에 관련한 지표들의 변화를 통계 분석하여 적정한 구과 채취시기를 구명하고자 하였다.
재료 및 방법
1. 조사 대상지 및 채종목 선정
일본잎갈나무 클론 채종원은 충청북도 충주시 수안보면 수회리 산40-1(36°52′29′′N, 127°58′30′′E)에 위치해 있으며, 1968년도에 조성되었다. 조성 면적은 7 ha이고 현존 본수는 495본(2017년 기준)이다. 여러 차례의 성숙단계별 구과를 채취하여야 하기 때문에 개화량이 충분하고 안정적인 시료 채취가 가능할 것으로 예측되는 10클론 25본(2021년) 그리고 11클론 26본(2022년)을 선정하였다.
2. 구과채취 시기
2021년에는 구과가 미성숙한 7월 말부터 자연적인 종자산포가 충분히 진행된 10월 초까지 채취하였다. 2주 간격으로 채취하였으며 구과 성숙이 진행되는 시기라고 여겨지는 8월 둘째 주부터 9월 둘째 주(Barnett, 1976) 사이에는 1주 간격으로 채취하여 총 8회 채취하였다. 2022년에는 채취 시기를 3주 앞당겼고, 전년도 조사로 일본잎갈나무 구과 및 종자 성숙 단계에 큰 변화가 있다고 여겨진 7월 말에서 8월 초에는 1주 간격으로 채취하였다.
3. 구과 형태적 특성 및 구과분석
구과의 형태적 변화를 관찰하기 위해 채취 시기별로 구과 형태와 내부의 종자가 온전한 구과를 채취하여 구과 크기(무게, 길이, 폭)와 색을 조사하였다. 채취 시기별로 같은 클론에서 5-7개 구과를 채취 직후 사진 촬영하였고 HSV 색 공간 변환 방법을 통해 구과 이미지 파일에서 갈색 면적(Hue 18 이하)을 도출하여 전체 구과 면적(픽셀 수) 대비 갈변한 면적의 비율을 계산했다. 이후에 클론당 1-3개 구과는 함수율 측정기(AND MF-50)를 이용해 구과 함수율을 측정하였다. 다만 구과 채취기간 동안 장마 기간이 겹쳤을 경우에는 이상치로 간주하여 통계 분석에서 제외하였다.
구과의 발달과 피해 정도를 조사하기 위해 각 채취 시기 별로 클론당 3-5개의 구과를 미 농무부(1977)의 가이드라인을 참고하여 분석하였다. 먼저 각 구과의 인편을 분리하여 염성 인편, 비염성 인편의 수를 세고 인편 안쪽의 1차 및 2차 고사배주, 충해종자, 성숙종자의 수를 측정하였다. 성숙 종자는 발아 실험 후 내부의 조직이 온전한 지 직접 확인하는 절단 검사를 거쳐 비립종자, 발아 또는 비발아 종자로 분류하였다.
4. 발아율 검정 및 절단 검사
충해종자를 제외하고 성숙종자만을 대상으로 클론 당 최대 50립, 최소 30립씩 3개 플레이트에 반복하여 발아 검사를 실시하였다. 발아 촉진을 위해 4°C에서 3주 간 수분을 공급하며 저온에서 보관하였고, 빛 8시간, 온도 25°C, 상대습도 70%로 유지되는 생장상으로 옮겨 28일 간 발아종자 수를 확인했다. 발아 조사 기간 동안 발아하지 않은 종자들은 직접 절단하여 배와 배유 조직이 온전한지 확인하고 비립 종자와 충실 종자로 구분했다.
구과분석 및 발아율, 절단 검사 결과 등을 이용하여 채종원의 종자 품질을 진단할 수 있는 지표인 종자생산능력, 고사 배주율, 충해 종자율, 비립종자율, 종자효율, 발아율 그리고 최종 발아 종자의 50%가 발아하는데 까지의 소요일수(T50)를 계산하였다(Table 3).
5. 구과 및 종자 특성 분석
측정된 구과 및 종자 특성이 채취시기 간 차이가 있는지 검증하기 위한 선형 회귀모델은 식 1과 같다.
Yik : i 번째 채취시기에 k 번째 개체로부터 얻어진 관측치
Ti : i 번째 채취시기의 효과
εik : i 번째 채취시기의 k 번째 개체에 대한 관측치의 임의오차(random error) 효과
통계적으로 채취시기 간 차이가 유의미한 경우 Tukey-Kramer HSD로 사후 검정하였고, 아래 공식 2와 같이 채취 시기, 클론, 시기와 클론 간의 상호작용 효과를 검증하였다.
Yijk : i 번째 채취시기에 j 번째 클론에서 k 번째 라멧으로부터 구해진 관측치
Cj : j 번째 클론의 상가적 유전효과
TCij : i 번째 시기와 j 번째 클론 간의 상호작용 효과
εijk : i 번째 시기에 j 번째 클론의 k 번째 라멧에 대한 관측치의 임의오차(random error) 효과
2021년 조사된 변수들의 공분산 행렬을 이용하여 주성분 분석을 실시하였으며, 2개의 주성분으로 군집화 과정을 거쳤다. 통계분석과 그래프 작성은 Python 3.9.7 version에서 Statsmodels, Pandas, Scipy, Matplotlib, Pillow Library와 SAS JMP 17.1.0를 활용하였다.
결 과
1. 구과의 형태적 특성(크기와 색)과 종자 생산 변화
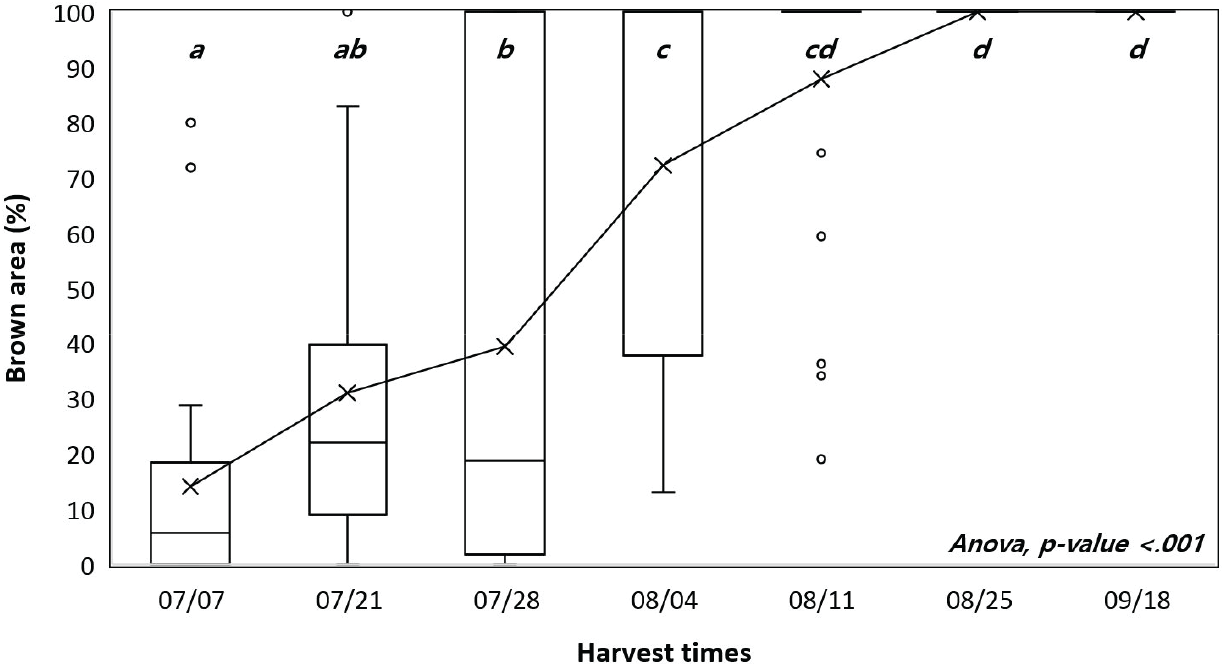
채취시기에 따른 구과의 무게, 길이 및 폭은 채취시기에 따른 차이가 나타나지 않았다. 구과의 색은 8월 초부터 대부분의 채종목에서 갈색으로 변하기 시작하였고, 빠른 경우는 7월부터 갈변이 시작된 개체도 있었으며 8월 말부터는 모든 구과가 갈색을 띄었다(Figure 1). 채취 시기 별로 평균을 비교했을 때, 7월 28일부터 8월 4일까지 비교적 짧은 기간 동안 그래프의 기울기가 가장 큰 변화가 나타났다.
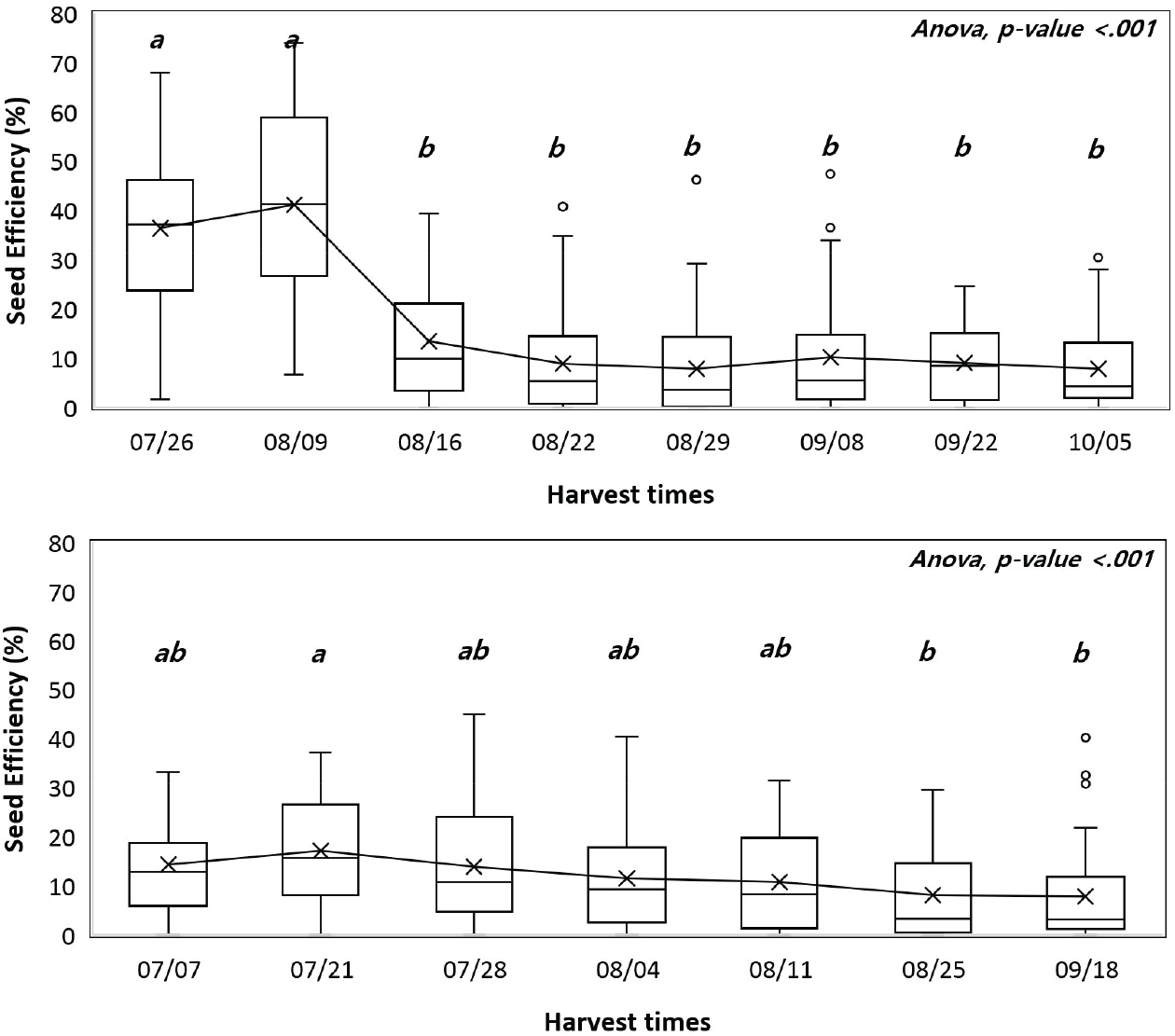
고사 배주율은 조사가 시작된 7월 이후에는 감소 또는 증가하는 경향성이 나타나지 않았다. 비립 종자율과 충해 종자율은 2022년 충해 종자율을 제외하고 채취시기 간 유의미한 차이가 나타났다(p<.001). 2021년(52.52%)에는 2022년 (49.52%)보다 연평균 충해 종자율이 약 3% 높게 나타났으며, 2021년에는 최소-최대값의 차이가 57.05%으로 20.46%인 2022년 보다 높게 나타났다(Table 2). 또한, 조사 시기가 늦어질수록 충해 종자율이 높아졌으며 채취시기 간의 통계적 차이도 유의하였다(p<.001). 그러나 2022년에는 최초 조사일부터 충해 종자율이 42.17%로 높게 조사되었고 시기 간의 차이도 유의하지 않았으며(p>.05) 충해 종자율이 조사 기간 동안 꾸준히 높게 나타났다(Table 2). 종자 효율은 2021년에는 8월 9일(41.15%)부터 8월 16일(13.37%) 사이에 유의미한 감소가 나타났고 이후에는 차이가 없었으며, 2022년에는 7월 28일(13.88%) 이후에 점차 감소했고 8월25일 이후로는 통계적인 차이가 없었다(Figure 2). 또한 2021년에 종자 효율은 충해 종자율과 높은 확률의 부의 상관관계(−0.88)가 나타났으며 비립 종자율과 양의 상관관계(0.41)를 보였다(Table 3).
Table 1.
Indexes and formulas of cone and seed quality.
|
Index |
Formula |
|
Seed potential |
Fertile scale × 2 |
|
Percent of aborted ovules (%) |
Aborted ovules / Seed potential × 100 |
|
Percent of insect-damaged seeds (%) |
Insect-damaged seeds / Developed seeds × 100 |
|
Percent of empty seeds (%) |
Empty seeds / Developed seeds × 100 |
|
Seed efficiency (%) |
Filled seeds / Seed potential × 100 |
|
Percent of germinated seeds (%) |
Germinated seeds / Total seeds × 100 |
|
T50
|
|
|
(Number of days until achieve 50% of the final germination rate) |
N, final number of germinated seeds; Ni, number of seeds just before 50% germination for N; Nj, number of seeds just after 50% germination for N; Ti, number of days for Ni; Tj, number of days for Nj. |
Download Excel Table
Table 2.
Seed production among cone harvest times.
|
Index |
Mean |
Year |
Harvest times |
P-value |
|
2021 |
|
|
07/26 |
08/09 |
08/16 |
08/22 |
08/29 |
09/08 |
09/22 |
10/05 |
|
2022 |
07/07 |
07/21 |
07/28 |
08/04 |
08/11 |
08/25 |
09/18 |
|
|
|
|
Aborted ovules rate (%) |
12.72 |
2021 |
- |
- |
12.13 |
14.96 |
16.93 |
9.13 |
9.66 |
6.16 |
14.55 |
16.45 |
<.001 |
|
20.91 |
2022 |
39.13 |
15.42 |
16.35 |
18.99 |
18.07 |
19.66 |
19.09 |
- |
- |
- |
<.001 |
|
Insect-damaged Seeds rate (%) |
52.52 |
2021 |
- |
- |
25.8 |
16 |
49.71 |
67.63 |
73.05 |
57.07 |
69.79 |
69.87 |
<.001 |
|
49.52 |
2022 |
42.17 |
38.94 |
51.02 |
45.9 |
56.15 |
59.4 |
53.86 |
- |
- |
- |
>.05 |
|
Empty seeds rate (%) |
25.15 |
2021 |
- |
- |
25.72 |
32.1 |
31.73 |
20.31 |
17.82 |
30.67 |
18.45 |
19.81 |
<.001 |
|
35.43 |
2022 |
65.82 |
65.05 |
64.68 |
74.12 |
73.37 |
76.15 |
78.25 |
- |
- |
- |
<.001 |
Download Excel Table
Table 3.
Pearson’s correlation coefficients of 10 variables in 2021.
|
|
Cone length |
Cone width |
Cone dry weight |
Aborted ovule |
Insect-damaged seed |
Empty seed |
Seed efficiency |
Seed germination |
T50
|
Moisture contents |
|
Cone length |
- |
0.54***
|
0.87***
|
−0.14 |
−0.03 |
−0.15 |
0.19 |
−0.05 |
0.26 |
0.09 |
|
Cone width |
|
- |
0.71***
|
0.03 |
−0.01 |
−0.13 |
0.14 |
0.2 |
−0.11 |
−0.14 |
|
Cone dry weight |
|
|
- |
−0.12 |
−0.28*
|
−0.04 |
0.48***
|
0.1 |
0.23 |
0.2 |
|
Aborted ovule |
|
|
|
- |
−0.31**
|
0.44***
|
0.07 |
0.23 |
−0.25*
|
−0.14 |
|
Insect-damaged seed |
|
|
|
|
- |
−0.78***
|
−0.88***
|
−0.13 |
−0.36**
|
−0.41***
|
|
Empty seed |
|
|
|
|
|
- |
0.41***
|
0.02 |
0.18 |
0.14 |
|
Seed efficiency |
|
|
|
|
|
|
- |
0.11 |
0.44***
|
0.52***
|
|
Seed germination |
|
|
|
|
|
|
|
- |
−0.43***
|
−0.07 |
|
T50 |
|
|
|
|
|
|
|
|
- |
0.35**
|
|
Moisture contents |
|
|
|
|
|
|
|
|
|
- |
Download Excel Table
2. 구과의 함수율 및 종자품질 변화
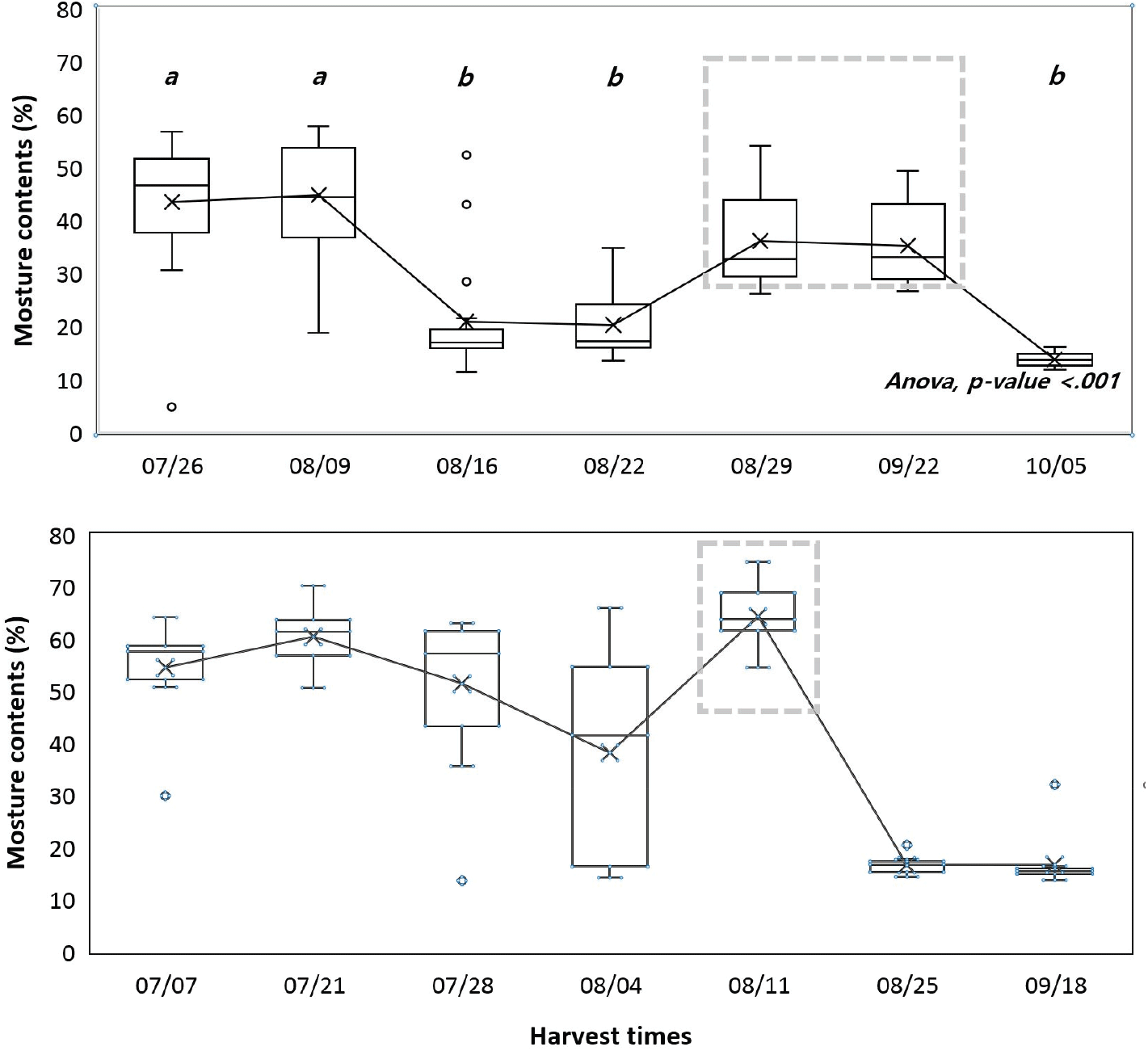
7월 중순 이전에는 구과 함수율의 변화가 없었으며, 2021년에는 8월 9일(44.81%)부터 16일(20.89%), 2022년에는 7월 28일(51.35%)부터 8월 4일(38.12%) 사이에 유의미한 감소를 보였다(Figure 3). 구과 함수율의 변화는 구과의 색깔 변화보다 늦게 시작되었고 함수율과 갈변 모두 8월 초에 급격한 변화가 나타났다.
Figure 3.
Moisture contents among harvest times in 2021 (above) and 2022 (below). Dotted boxes are outliers due to rain 1 day ago.
Download Original Figure
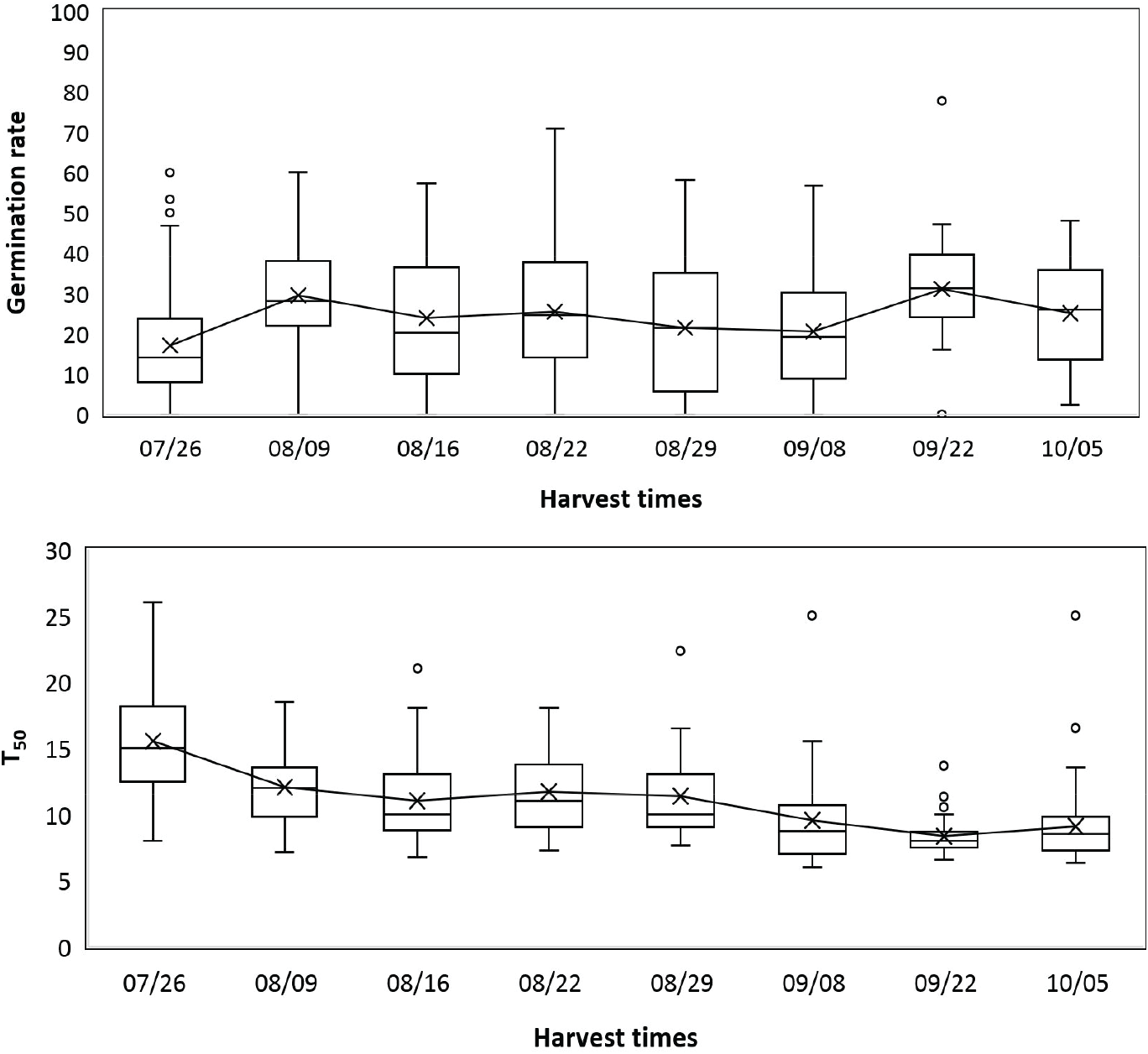
발아율은 2021년 채취시기 간에 유의미한 차이를 보였으며(p<.001), 첫번째 채취시기인 7월 26일(17.11%)에서 8월 9일(29.5%) 사이에 크게 증가했고 9월 8일(20.3%)까지는 점차 감소하다가 9월 22일(31.08%)에 가장 높았다(Figure 4). T50은 7월 26일(15.49일)에서 8월 9일(12.01일) 사이에 유의미하게 감소하였고(p<.001) 8월 29일(11.33) 이후로는 시기가 늦어질수록 발아 속도가 빨라져 9월 8일 이후에는 10일 이하로 나타났다(Figure 4). 또한 T50과 함수율, 종자효율의 상관관계는 유의한 양의 계수(0.35, 0.44)가 나타났으며 발아율, 충해 종자율은 부의 상관(−0.43, −0.36)이 있었다(Table 3). 2022년의 발아율은 채취시기 간에 통계적인 차이가 있었으나(p<.001) 값이 매우 낮아(평균 1%) 발아율에 영향을 준 다른 요인이 있었을 것으로 사료되었다. 이에 2022년도 발아율 검정 결과 자료는 채취시기 영향을 보기 어렵다고 판단하여 이후 분석에서 제외하였다.
3. 구과 및 종자 특성에 대한 채취시기 간 유전적 효과 분석
함수율, 종자 효율, 발아세에 대해 채취시기 효과, 유전적 효과, 상호작용 효과를 검증하고자 이원분산분석을 실시하였다(Table 4). 세 지표 모두 일원분산분석과 마찬가지로 채취시기에 대해 유의미한 효과가 있었다(p<.001). 종자 효율과 발아세는 유전적인 차이가 있었으나(p<.001), 함수율에서는 나타나지 않았다. 채취시기와 유전적 효과 간의 상호작용 효과는 발아세에서 유의하게 나타났고(p=.004), 종자효율은 2021년도에서만 5% 유의수준(p=.013)에서 차이를 나타냈으며, 함수율은 2022년에만 유의미한 차이(p<.001)를 나타났다.
Table 4.
Two-way ANOVA for moisture contents, seed efficiency, and T50.
|
|
Year |
Moisture contents |
Seed efficiency |
T50
|
|
DF |
Sum of squares |
Mean of squares |
F-value |
P-value |
DF |
Sum of squares |
Mean of squares |
F-value |
P-value |
DF |
Sum of squares |
Mean of squares |
F-value |
P-value |
|
Harvest time |
2021 |
4 |
16401.73 |
4100.43 |
53.19 |
<.001 |
7 |
76966.88 |
10995.27 |
77.23 |
<.001 |
7 |
1841.64 |
263.09 |
37.33 |
<.001 |
|
2022 |
5 |
21624.95 |
4324.99 |
54.01 |
<.001 |
6 |
2541.95 |
423.66 |
5.41 |
<.001 |
|
|
|
|
|
|
Clone |
2021 |
9 |
1088.05 |
120.89 |
1.57 |
0.15 |
9 |
5603.55 |
622.62 |
4.37 |
<.001 |
9 |
683.59 |
75.95 |
10.78 |
<.001 |
|
2022 |
3 |
547.86 |
182.62 |
2.28 |
0.09 |
10 |
7316.24 |
731.62 |
9.34 |
<.001 |
|
|
|
|
|
|
Harvest time * Clone |
2021 |
36 |
2766.7 |
76.85 |
1 |
0.5 |
63 |
13470.37 |
213.82 |
1.5 |
0.013 |
63 |
720.87 |
11.44 |
1.62 |
0.004 |
|
2022 |
15 |
3998.72 |
266.58 |
3.33 |
<.001 |
60 |
5988.44 |
99.81 |
1.27 |
0.111 |
|
|
|
|
|
|
Error |
2021 |
57 |
4394.23 |
77.09 |
|
|
349 |
49686.92 |
142.37 |
|
|
317 |
2234.27 |
7.05 |
|
|
|
2022 |
47 |
3763.39 |
80.07 |
|
|
198 |
15508.21 |
78.32 |
|
|
|
|
|
|
|
Download Excel Table
4. 구과와 종자특성에 대한 주성분 분석
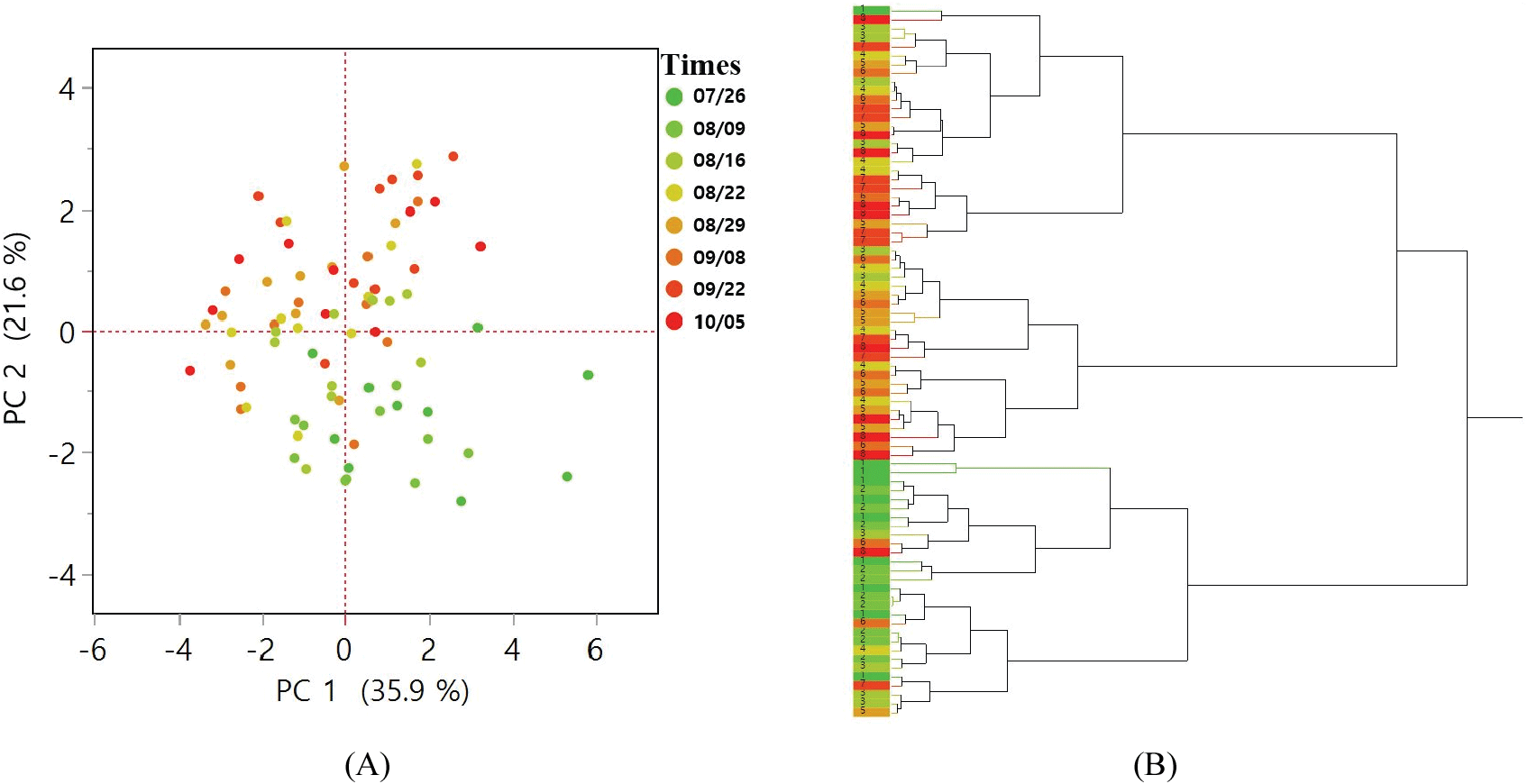
종자 생산 및 품질과 관련된 10개의 변수를 2021년 데이터로 주성분 분석을 수행한 결과, 제1주성분과 제2주성분의 누적기여도는 57.45%이었다. 제1주성분은 구과의 형태적 특성과 관련한 변수들인 구과 길이, 구과 폭, 구과 무게, 염성인편 수, 고사배주 수, 비립종자 수, 충실종자 수의 기여도가 높았고, 제2주성분은 채취시기에 따른 종자품질 변화와 관련된 변수들인 충해종자 수, 비립종자 수, 충실종자 수, 발아세에 대한 기여도가 높았다(Table 5). 4번째 채취시기(8월 22일) 이후로 대부분의 클론에서 제2주성분이 양수로 나타났으며, 군집도에서 가장 크게 분기된 두 그룹은 각각 7월 26일~8월 16일과 8월 22일~10월 5일에 채취된 클론들이다(Figure 5).
Table 5.
Covariance matrix of 10 variables with the 2 principal component scores.
|
Variable |
PC1 |
PC2 |
|
Cone Length |
0.84 |
0.34 |
|
Cone Width |
0.53 |
0.27 |
|
Cone Dry Weight |
0.92 |
0.15 |
|
Fertile Scale |
0.84 |
0.3 |
|
Aborted Ovule |
0.49 |
0.03 |
|
Insect-damaged Seed |
−0.04 |
0.92 |
|
Empty Seed |
0.49 |
−0.63 |
|
Filled Seed |
0.66 |
−0.59 |
|
Germination rate |
0.26 |
0.16 |
|
T50
|
0.26 |
−0.49 |
|
Variance Explained (%) |
35.89 |
21.56 |
Download Excel Table
Figure 5.
(A) PCA of 10 variables related to cone and seed quality, (B) Hierarchical clustering for 2 principal components. The colors represent the harvest times in 2021.
Download Original Figure
고 찰
1. 종자채취 적기
본 연구는 일본잎갈나무 종자 생산을 위한 채취적기를 구명하기 위해 두 해에 걸쳐 시기별로 구과를 채취하여 구과 특성과 종자 품질을 분석하였으며, 이에 따른 종자의 채취 적기는 크게 종자 품질과 생산량이라는 두 가지 측면에서 논의될 수 있다. 첫째, 종자 품질 측면에 있어서는, 종자의 발아율이 유의하게 높아진 시기는 조사 결과 8월 초였으며, 발아속도 또한 8월 초부터 빨라지기 시작하였고 구과 채취가 늦어질수록 발아속도가 빠른 종자를 얻을 수 있었다. 그러나 종자의 성숙은 연기후에 영향을 받으므로(Buechling et al., 2016; Henttonen et al., 1986), 연중 채취일을 특정하는 것보다 채취 적기를 판단하는 지표를 마련하는 것이 중요하다. 이와 관련된 구과 특성을 살펴보면, 8월 초에는 구과의 갈변이 급격히 증가하면서 구과 함수율 역시 감소하기 시작하였으며, 이 시기는 종자의 발아능이 높아진 시기와도 일치한다. 또한 모든 구과가 갈변한 8월 말에는 구과 함수율 역시 유의미하게 감소하였는데, 건조저항성이 있는 진정종자(orthodox seed)의 경우 열매나 구과가 성숙한 후 건조함에 따라 건조저항성을 획득하고 이후 종자세가 증가한다(Bewley et al., 2013; Society for Ecological Restoration, International Network for Seed Based Restoration and Royal Botanic Gardens Kew, 2023). 따라서 진정종자인 일본잎갈나무 종자의 경우에도 구과의 색이 변하고 함수율이 감소하기 시작하는 시기에 발아능을 먼저 획득하고, 구과 함수율 감소와 종자 성숙이 계속되면서 종자세가 증가한 것으로 볼 수 있다. 이와 같이 구과의 갈변 정도와 함수율은 종자 품질과 큰 상관이 있었으며 현장에서 채종목 집단의 종자 채취 적기를 판단하는 간편한 지표로 활용할 수 있을 것으로 보인다.
종자 생산량 측면에서의 채취 적기는 병해충 피해, 종자 비산 등에 의한 손실을 최소화하여 충실 종자 생산량을 최대로 하는 시기여야 한다. 이를 위해 미국 농무성에서 채종원의 종자 생산을 진단하기 위해 개발된 구과 분석 방법과 지표들을 활용하였다(Bramlet et al., 1977). 그 결과, 구과의 전체 종자생산능력 대비 충실 종자 비율을 나타내는 지표인 종자 효율이 2021년에는 8월 16일, 2022년에는 7월 21일 이후에 점차 감소한 것을 확인할 수 있었다. 이 같은 경향은 충해 종자율이 채취 시기가 늦어짐에 따라 증가하는 추세를 보였으므로 주로 해충 피해에 기인한 것으로 보인다.
이와 같은 결과를 종합할 때, 충실 종자 생산량을 최대로 하면서 발아율이 높은 종자를 얻기 위한 채취 적기는 8월 초로 나타났으며, 이는 구과의 갈변 면적이 크게 증가하고 구과 함수율이 크게 감소하는 시기와 일치한다. 이후에는 종자 성숙이 진행되어 종자세가 증가하나 병해충 피해와 종자 비산에 따른 손실이 있으므로, 종자 품질을 우선시한다면 해충 발생 시기에 적절한 방제를 시행하고 채취 시기를 늦추는 전략도 가능할 것이다. 구과의 성숙 시기는 연간 기후나 채종원 관리에 따라 달라질 수 있으므로 육안으로 확인하여 대부분의 구과에서 갈변이 약 70% 진행되었을 때를 적정 채취시기로 결정하는 것을 제안한다.
2. 구과특성 및 종자품질
본 연구는 2021년과 2022년, 두 해에 걸쳐 동일한 품질 지표를 대상으로 수행되었으나, 각 지표들의 경향성이 다르게 나타났다. 2022년의 경우, 발아율이 평균 1%로 매우 저조하였고, 고사 배주와 비립 종자 비율도 2022년에 각 8%, 10% 만큼 더 높았다(Table 2). 이는 화분 부족이나 수정, 발달 시기의 병해충 피해 또는 매우 낮은 암구화수 생산량(개체목당 평균 4개)과 생식능력 저하가 원인일 수 있다. 특히 일본잎갈나무는 침엽수 중에서도 종자 생산의 풍흉 주기(masting period)가 길고 차이가 큰 것으로 알려져 있다. 침엽수의 풍흉 주기는 꾸준한 종자 생산에 제한 요소가 되며(Creain and Cregg, 2018, Sala et al, 2012), 2022년에는 극심한 흉년으로 해당 일본잎갈나무 채종원의 생식능력이 저하된 것으로 보였다.
충해종자 비율은 채취시기가 늦어질수록 증가하는 경향이 있었던 2021년과 다르게 2022년에는 7월 초부터 40% 이상으로 높게 나타났다(Table 4). 이는 2022년에 매우 저조했던 암구화수 생산으로 인해 암배우체나 접합자 단계부터 충해를 많이 받았던 것으로 보였다. 충해 종자 비율은 종자 생산량에 직접적으로 영향을 주는 지표이므로 암구화수 생산량이 적은 해에는 종자효율을 최대화하기 위해 특히 해충 방제가 중요하다.
주성분 분석과 군집분석을 통해 채취시기별로 구과와 종자 특성을 그룹화하였다. 군집화에서 크게 두 그룹으로 나뉘었고 한 그룹은 대부분 채취시기 7월26일~8월16일에, 다른 그룹은 8월22일~10월5일에 속했다(Figure 5). 이에 따라 본 일본잎갈나무 채종원의 구과와 종자 특성이 크게 변화한 시기는 8월 초에서 중순으로 나타났다. 이 시기는 구과의 갈변과 함수율에 큰 변화가 있었던 시기와 동일하다(Figures 1, 3). 대부분의 지표들은 클론에 따라 차이가 나타났던 반면 함수율은 통계적으로 유의한 유전적 차이가 나타나지 않는 지표였다(Table 4). 따라서 함수율과 구과의 갈변 정도는 채종원 관리자가 채종목의 구과 성숙 상태를 파악할 수 있는 적절한 지표가 될 수 있다.
종자 품질에는 종자 수명과 저장 동안의 성숙 정도도 영향을 미칠 수 있다(Bonner, 1987; Pieta and Ellis, 1991). 본 연구에서는 채취 후 바로 파종하였을 때의 종자 발아율과 발아 속도만을 확인하였으므로 주요 조림 수종의 장기간 종자 보관을 위해서는 채취시기별 종자 저장 및 품질 유지와 해당 종자로부터 생산된 묘목의 품질 평가에 대한 연구가 추후 수행되어야 한다.
감사의 글
본 연구는 국립산림품종관리센터의 지원으로 수행된 연구용역사업 ‘낙엽송 개화특성 및 구과 채취시기별 종자 품질조사(2021)’, ‘낙엽송 결실 특성 조사 및 채종원 식재 배열 연구(2022)’ 연구 결과의 일부입니다. 시료 채취 및 현장조사에 도움을 주신 국립산림품종관리센터 관계자들께 감사드립니다.
References
Adams, W.T., Hipkins, V.D., Burczyk, J. and Randall, W.K. 1997. Pollen contamination trends in a maturing Douglas-fir seed orchard. Canadian Journal of Forest Research 27(1): 131-134.


Barnett, J.P. 1976. Cone and seed maturation of southern pines. Vol. 122. Department of Agriculture, Forest Service, Southern Forest Experiment Station.
Bedane, G.M., Gupta, M.L. and George, D.L. 2006. Optimum harvest maturity for guayule seed. Industrial Crops and Products 24(1): 26-33.
Bewley, J.D., Bradford, K., Hilhorst, H. and Nonogaki, H. 2013. Seeds: physiology of development, germination and dormancy. 3
thEd. Springer Science & Business Media.
Bramlett, D.L. 1977. Cone analysis of southern pines: a guidebook. Vol. 13. Department of Agriculture, Forest Service, Southeastern Forest Experiment Station.
Bonner, F.T. 1987. Effect of storage of loblolly and slash pine cones on seed quality. Southern Journal of Applied Forestry 11(1): 59-65.
Buechling, A., Martin, P.H., Canham, C.D., Shepperd, W.D. and Battaglia, M.A. 2016. Climate drivers of seed production in
Picea engelmannii and response to warming temperatures in the southern Rocky Mountains. Journal of Ecology 104(4): 1051-1062.
Crain, B.A. and Cregg, B.M. 2018. Regulation and management of cone induction in temperate conifers. Forest Science 64(1): 82-101.
Egli, D.B. 2017. Seed biology and yield of grain crops. 2
nd Edition. CABI.
Elias, S.G. and Copeland, L.O. 2001. Physiological and harvest maturity of canola in relation to seed quality. Agronomy Journal 93(5): 1054-1058.
Ellis, R.H. and Pieta Filho, C. 1992. The development of seed quality in spring and winter cultivars of barley and wheat. Seed Science Research 2(1): 9-15.
Grosman, D.M., Upton, W.W., McCook, F.A. and Billings, R.F. 2002. Systemic insecticide injections for control of cone and seed insects in loblolly pine seed orchards-2 year results. Southern Journal of Applied Forestry 26(3): 146-152.
Gu, R., Li, L., Liang, X., Wang, Y., Fan, T., Wang, Y. and Wang, J. 2017. The ideal harvest time for seeds of hybrid maize (
Zea mays L.) XY335 and ZD958 produced in multiple environments. Scientific Reports 7(1): 17537.


Harrington, J.F. and Kozlowski, T.T. 1972. Seed storage and longevity. Seed biology 3, 145-245.
Henttonen, H., Kanninen, M., Nygren, M. and Ojansuu, R. 1986. The maturation of
Pinus sylvestris seeds in relation to temperature climate in northern Finland. Scandinavian Journal of Forest Research 1(1-4): 243-249.
Kang, K.S. and Lindgren, D. 1999. Fertility variation among clones of Korean pine (
Pinus koraiensis S. et Z.) and its implications on seed orchard management. Forest Genetics 6(3): 191-200.
Kelly, A.F. and George, R.A. 1998. Encyclopaedia of Seed Production of World Crops. John Wiley and Sons, Chichester.
Kim, I.S., Kim, J.H., Kang, J.T., and Lee, B.S. 2008. Clonal variation in female flowering of
Larix leptolepis. Korean Journal of Plant Resources 21(1): 1-4.
Korea Forest Service. 2022. Statistical Yearbook of Forestry No. 52.
Lee, W.Y., Park, E.J., Han, S.U. and Park, B.S. 2012. Strobilus Production Characteristics and Enhancement for Seed Production of
Larix kaempferi. National Institute of Forest Science, Research Report 457: 5-18.
Likoswe, M.G., Njoloma, J.P., Mwase, W.F. and Chilima, C.Z. 2008. Effect of seed collection times and pretreatment methods on germination of
Terminalia sericea Burch. ex DC. African Journal of Biotechnology 7(16).
Lindgren, D., Danusevicius, D. and Rosvall, O. 2009. Unequal deployment of clones to seed orchards by considering genetic gain, relatedness and gene diversity. Forestry 82(1): 17-28.
Lstibůrek, M. and El-Kassaby, Y.A. 2010. Minimum-inbreeding seed orchard design. Forest Science 56(6): 603-608.
Matsushita, M., Nishikawa, H., Tamura, A. and Takahashi, M. 2020. Effects of light intensity and girdling treatments on the production of female cones in Japanese larch (
Larix kaempferi (Lamb.) Carr.): Implications for the management of seed orchards. Forests 11(10): 1110.
Mou, B. and Kronstrad, W.E. 1994. Duration and rate of grain filling in selected winter wheat populations: I. Inheritance. Crop Science 34: 833–837.
Owens, J.N. and Molder, M. 1979. Bud development in
Larix occidentalis. I. Growth and development of vegetative long shoot and vegetative short shoot buds. Canadian Journal of Botany 57(7): 687-700.
Owens J.N. and Blake M.D. 1985. Forest tree seed production. A review of literature and recommendations for future research. Canadian Forestry Service Information Report No.PI-X-53, pp. vi+161.
Probert, R., Adams, J., Coneybeer, J., Crawford, A. and Hay, F. 2007. Seed quality for conservation is critically affected by pre-storage factors. Australian Journal of Botany 55(3): 326-335.
Rosenberg, O. and Weslien, J. 2005. Assessment of cone-damaging insects in a Swedish spruce seed orchard and the efficacy of large-scale application of
Bacillus thuringiensis variety
aizawai× kurstaki against Lepidopterans. Journal of Economic Entomology 98(2): 402-408.
Sala, A., Hopping, K., McIntire, E.J., Delzon, S. and Crone, E.E. 2012. Masting in whitebark pine (
Pinus albicaulis) depletes stored nutrients. New Phytologist 196(1): 189-199.
Society for Ecological Restoration, International Network for Seed Based Restoration and Royal Botanic Gardens Kew. 2023. Seed Information Database (SID). Available from:
https://ser-sid.org/ (February 2023)
Torimaru, T., Wang, X.R., Fries, A., Andersson, B. and Lindgren, D. 2009. Evaluation of pollen contamination in an advanced Scots pine seed orchard. Silvae Genetica 58(1-6): 262-269.
Zanakis, G.N., Ellis, R.H. and Summerfield, R. J. 1994. Seed quality in relation to seed development and maturation in three genotypes of soyabean (
Glycine max). Experimental Agriculture 30(2): 139-156.