서 론
생명자원의 보존은 해당 생물의 서식지 내에서 보존하는 현지내(in-situ) 보존과 서식지 밖에서 보존하는 현지외(ex-situ) 보존으로 크게 구분할 수 있다. 이 중 현지외 보존은 기후 변화나 인위적 교란 등 환경 변화에 따른 서식지 쇠퇴 및 유전자원의 소실이 우려되는 상황에서 생명자원의 보존과 복원재료를 확보하기 위한 주요 대응 전략이다. 현지외 보존 전략 중에서도 종자 은행(seed bank)은 가장 쉽고 효율적인 방법 중 하나이다(Linington and Pritchard, 2001). 종자를 이용한 현지외 보존을 효율적으로 수행하기 위해서는 해당 종자의 형태적 특성, 휴면 원인과 발아 조건, 저장성 등에 대한 기초 연구가 뒷받침되어야 하며, 이에 맞는 저장 전략을 수립해야 한다.
종자의 저장성과 건조 특성을 이해하는 것은 현지외 보존의 기본이며, 종자가 건조에 견디는 정도에 따라 저장성 종자(orthodox seed)와 난저장성 종자(recalcitrant seed), 그리고 그 중간 특성을 가지는 종자(intermediate seed)로 분류할 수 있다(Hong and Ellis, 1996). 저장성 종자는 5~10% 이내의 낮은 함수율 수준 또는 약한 결합수가 제거되는 정도까지 건조하여도 활력을 잃지 않고 수년 이상 저장할 수 있다(Roberts, 1973 in Pritchard, 2004; Walters et al., 2005). 난저장성 종자는 초기 함수율이 30~40%로 높고, 유리수만 제거되어도 쉽게 활력을 잃기 때문에(Pritchard, 2004) 사실상 건조가 불가능하여 저장 중 발아하거나, 곰팡이 감염 및 노화 등으로 저장이 어려운 특징을 갖는다. 이들의 중간 특성을 가지는 중간저장성 종자(intermediate seed)는 함수율 10~12% 수준 또는 결합수가 제거되지 않는 정도까지 건조될 수 있으나, 이 이상으로 건조하면 활력이나 저장성이 떨어진다(Pritchard and Dickie, 2003).
저장성 종자의 경우, 발달 과정 중 건조 내성(desiccation tolerance)을 획득한다. 물은 세포의 원형질막과 고분자의 3차 구조를 유지하는 데 필수적이므로, 일정 수준 이상으로 건조될 경우 건조나 재수화 과정에서 세포가 손상을 입게 되는데, 건조 내성이 있는 경우 이를 복구하는 방어 기작을 가진다(Bewley et al., 2013). 예를 들면, 대부분의 저장성 종자들은 발달 단계에서 세포에 당을 축적하는데, 이들이 물을 대신하여 원형질막이나 단백질의 구조를 유지하게 한다는 가설이 있으며(Koster and Leopold, 1988; Chen and Burris, 1990; Bryant et al., 2001), 종자 발달 단계에서 축적되는 late embryogenesis abundant (LEA), heat-shock proteins (HSPs) 단백질, 그리고 LEA 단백질 생성에 영향을 주는 abscisic acid (ABA) 역시 건조 내성과 연관이 있다고 알려져 있다(Oliver et al., 2020). 그러나 난저장성 종자와 같이 건조에 민감한(desiccation sensitive) 수종의 경우 세포막 손상을 회복하지 못하며, 배의 양분전달체계에 피해를 입거나 유근 또는 자엽 발육에 문제가 생기는 등의 피해를 입게 된다(Kim et al., 2014). 따라서 수종별로 종자의 건조 내성을 파악하는 것은 현지외 보존 전략 수립을 위한 첫번째 단계라고 할 수 있으며(Millenium seed bank, 2022), 보존 대상 수종의 종자가 건조 내성을 가지는지 여부와 활력을 잃지 않는 건조 수준을 확인하는 것이 반드시 필요하다.
죽절초(Sarcandra glabra)는 주로 동남아, 중국 남부, 일본 등 아열대 지방에 흔하게 분포하는 상록 활엽 관목으로, 우리나라에서는 분포의 북방 한계선에 가까운 서귀포시 상록활엽수림에서만 드물게 분포한다. 건조한 죽절초는 전통적으로 감기, 종양 등의 질병을 치료하는 데 쓰거나, 차로 음용되어 왔다(Liu et al., 2016; Zeng et al., 2021). 죽절초 추출물은 항염작용(Yaermaimaiti et al., 2016), 항생작용 등이 있는 것으로 보고되었고, 특히 항암작용, 혈소판 감소증에 대한 효과가 임상적으로 검증되었다(Zeng et al., 2021). 또한 열매가 10월부터 붉게 성숙하여 이듬해 2월까지 남아있기 때문에 겨울 내내 붉고 둥근 열매를 감상할 수 있어 관상가치가 높다. 이렇게 죽절초는 유용식물자원으로서, 그리고 관상식물로서의 가치가 매우 높은 식물이나, 국내에서는 자생지가 한정적인 데다 무분별한 채취가 이루어져 위협을 받고 있다(National institute of biological Resources, 2018). 현재 우리나라에서는 환경부 멸종위기 야생생물 II급, 국가적색목록 위기종(EN), 수목원∙정원의 조성 및 진흥에 관한 법률 시행규칙 제2조3항에 따른 희귀식물 멸종위기종(CR)로 분류되었다.
일반적으로 죽절초는 실생 번식을 하고 발아율은 90% 가까이 되는 것으로 알려져 있으며(Seo et al., 2006), 삽목(Seo et al., 2006), 조직배양(Kil et al., 2012)을 이용한 증식 연구가 이루어졌다. 실생묘에 관한 연구로는 죽절초의 파종 및 육묘법에 관한 연구에서 외종피의 유무가 발아에 영향을 미치지 않았다고 보고된 바 있다(Tu, 2018). 그러나 죽절초나 동일 속의 현지외 보존을 위해 필요한 종자 건조 내성과 저장 특성과 같은 정보는 Millennium seed bank에서 제공하는 Seed information database에 제시되어 있지 않으며(Kew, 2023), 학술적인 연구 결과 역시 보고된 바 없다. 일반적으로, 고온 다습한 환경에서 자라는 수종은 건조에 민감한 종자를 가지는 경우가 많은 것으로 알려져 있다. 즉, 죽절초는 아열대 지방에 주로 분포하는 수종으로 종자를 이용한 현지외 보존 전략 수립을 위해 먼저 건조 내성 및 저장 특성을 확인하는 것이 반드시 필요하다.
따라서 본 연구는 죽절초의 현지외 보존을 위해 종자 기초 특성과 건조 내성 및 저장성에 관한 정보를 얻기 위해 수행되었다. 이를 위해 죽절초 종자를 채집하여 먼저 기본적인 발아 특성을 조사하였으며, 종자를 여러 함수율 수준으로 건조하여 발아 특성을 비교하고 건조에 의한 세포막 기능의 손상을 간접적으로 확인하기 위해 종자 침출수 분석을 수행하였다. 또한 저장에 따른 발아 특성을 비교하여 종자의 저장성을 확인하고자 하였다.
재료 및 방법
본 연구를 위하여 2023년 2월 2일 국립산림과학원 난대아열대산림연구소의 구내 보존원에서 죽절초 열매를 채취하였다. 채취 직후 열매 형태 특성을 조사하였으며, 과육을 제거하여 정선한 후 종자 형태 특성을 조사하였다. 열매는 원형 혹은 타원형이며, 긴 지름 6.20±0.36mm, 짧은 지름 5.76±0.39mm, 1개당 무게 0.14±0.02g로 조사되었다. 종자는 원형 혹은 타원형으로, 긴 지름 3.79±0.21mm, 짧은 지름 3.16±0.09mm, 천립중 21.3±0.04g이었다. 탈종 직후의 종자 함수율은 32.8±1.5%였다.
죽절초 종자의 기본 발아율을 조사하고, 휴면에 대한 정보를 얻기 위해 탈종 직후의 종자를 무처리와 30일간 저온습윤처리로 구분하여 각각 발아 실험을 실시하고, 발아 특성을 나타내는 지수들을 계산하였다. 발아 실험은 직경 90mm 페트리 디쉬에 여과지(Whatman No. 2) 2장을 깔고 증류수로 적신 것을 발아상으로 사용하였고, 죽절초 종자를 25립 4반복으로 치상하였다. 치상한 종자는 24 °, 광 16시간, 암 8시간으로 설정된 생장상에 넣어 발아시켰다. 저온습윤처리는 발아지(Anchor paper)로 종자를 감싸고 증류수로 적셔 폴리에틸렌 봉투에 넣은 후, 밀봉하지 않은 채로 4 °에서 30일간 처리한 후 발아 실험하였다. 발아 조사는 유근이 2mm 이상 자라나온 종자를 발아한 것으로 간주하고 발아가 일어나지 않을 때까지 매일 발아한 종자의 수를 조사하였다.
발아 실험 데이터를 이용한 종자 발아 특성은 종자의 활력을 평가하기 위한 발아율(GP, germination percentage), 종자세를 평가하기 위한 T50(Farooq et al., 2005), 평균발아일수(MGT, mean germination time)(Labouriau, 1983 in Ranal and Santana, 2006), 발아속도(GS, germination speed)(AOSA, 1983 in McNair et al., 2012), 발아균일지수(CUG, coefficient of uniformity of germination)(Heydecker, 1973; Bewley and Black, 1994 in Ranal and Santana, 2006)를 계산하였다.
죽절초 종자의 건조 내성을 평가하기 위해, 채취 직후의 종자를 여러 함수율 수준으로 조절하여 발아 실험을 실시하였다(Hong and Ellis, 1996; Millennium seed bank, 2022). 종자의 건조는 종자 무게와 같은 양의 실리카 겔과 종자를 함께 용기에 담고 15°의 인큐베이터에서 건조하였다(Millennium seed bank, 2022). 건조 과정 중 매일 종자 무게를 측정하여 함수율 변화를 유추하였고, 건조 단계별로 총 3번의 시료를 채취하여 함수율을 측정하고 발아 실험을 실시하였다. 함수율은 종자의 건조 전 무게를 측정하고, 103° 건조기에서 17시간 건조한 후의 무게를 다시 측정하여 계산하였다. 함수율은 10립 3반복으로, 발아 실험은 25립 4반복으로 실시하였다.
죽절초 종자의 건조 내성에 대한 정보가 없었으므로, 건조하지 않은 종자의 저장 수명에 대한 정보를 얻기 위해 종자를 52일, 73일 저장한 후 발아 실험을 실시하였다. 저장 전 종자의 함수율은 28.1±1.4%이었으며, 종자를 폴리에틸렌 봉투에 넣고 진공 포장하여 4 °에서 저장하였다.
일정 기간 저장된 종자의 건조 내성과 건조에 따른 발아 특성 및 세포막 기능을 확인하기 위해, 52일간 저장된 종자를 대상으로 발아 실험을 수행하고 종자 침출수를 분석하였다. 대상 종자는 건조 단계별로 총 3번의 시료를 채취하였고, 함수율을 측정하고 종자 침지 및 발아 실험을 실시하였다. 종자 침지는 침출수 분석을 위해, 발아 실험 전 대상 종자를 10회 이상 세척하여 표면의 오염물질을 제거한 후, 3차 증류수 25ml에 넣고 교반기 위에 올려 24시간 동안 110rpm으로 회전하였다. 이후 종자를 분리하여 발아 실험을 수행하고, 침출수는 전기전도도를 측정하고 무기질을 분석하였다. 침출수의 전기전도도는 pH/Conductivity Meter (Orion Star A215 pH/Conductivity Benchtop Multiparameter Meter, Thermo Fisher Scientific Inc., Waltham, MA, USA)를 이용하여 측정하였다. 무기질 분석은 유도결합 플라즈마 분광광도계(PlasmaQuant PQ 9000 (Elite) High-resolution Array ICP-OES, Analytik Jena AG, Jena, Germany)를 이용하여 침출수에 포함된 Na, Mg, K, Ca, Mn, Fe, Cu, Zn의 농도를 분석하였다. 건조 내성을 평가하기 위한 함수율 측정은 10립 4반복, 종자 침지 및 발아 실험은 15립 4반복으로 실시하였다.
결 과
죽절초 종자의 휴면 여부를 확인하기 위해 저온습윤처리구와 무처리구를 비교한 결과, 30일 저온습윤처리한 경우, 무처리구와 비교하여 발아율, 발아속도 및 발아균일지수에서 통계적인 차이가 나타나지 않았다(Table 1). 발아율의 경우, 30일 저온습윤처리구에서 무처리구에 비해 평균 14% 더 높았으나, 반복 간 편차가 커 그 차이가 통계적으로 유의하지 않았으며, T50과 평균발아일수의 경우, 무처리구에서 각각 21.6일, 22.3일인 것에 비해 저온습윤처리구에서는 22.3일, 25.0일로 오히려 높은 값을 보여 발아를 촉진하는 효과는 나타나지 않았다.
Treatment cs30 means cold stratification for 30 days.
Each field value represents mean±standard deviation (n=4).
Different letters in a column represent significant difference among treatments according to Tukey’s HSD tests at α = 0.05.
죽절초 종자를 채취한 직후 종자에 대해 건조 수준에 따른 함수율과 발아 특성을 조사한 결과는 Table 2와 같았다. 죽절초 종자는 함수율의 변화에 따라 발아 특성이 달라졌으며, 통계적으로 유의한 차이가 있었다(p<0.05). 처리 I에서 함수율은 4.8%로 크게 감소했으나, 발아율과 T50, 평균발아일수, 발아속도는 감소하지 않았다. 즉, 종자 함수율의 감소에 따른 발아 활력과 종자세의 감소는 종자 수분 함량 변화가 일어난 후 곧바로 나타나지는 않은 것으로 보인다. 그러나 건조가 계속되어 종자 함수율이 각각 2.3%, 2.2%로 감소한 처리 II와 처리 III에서는 발아율이 각각 55.0%, 30.0%로 크게 감소하였으며, T50과 평균발아일수가 증가하였고, 발아 속도와 발아균일지수가 감소하여 활력과 종자세가 크게 떨어졌다.
The seeds were dried for 1, 7, and 13 days in treatment I, II, and III, respectively.
Each field value represents mean±standard deviation (n=4).
Different letters in a column represent significant difference among treatments according to Tukey’s HSD tests at α = 0.05
죽절초 종자의 저장 기간과 온도에 따른 발아 특성을 확인한 결과는 Table 3과 같았다. 대조구인 채취 직후의 종자(함수율 28.1±1.4%)를 각각 -18°, 4°, 25°에 52일, 73일 저장한 후 발아 시험을 실시한 결과, -18°에 저장한 종자들은, 종자의 동결이 일어났으며, 발아 상에서도 전혀 발아하지 않았다. 25°에 저장한 종자 역시 곰팡이 감염이심하게 나타나고, 전혀 발아하지 않아 활력을 모두 소실한
The seeds were stored for 52 and 73 days in treatment 52d and 73d, respectively.
Each field value represents mean±standard deviation (n=4).
Different letters in a column represent significant difference among treatments according to Tukey’s HSD tests at α = 0.05.
것으로 나타났다. 그러나 4°에 저장한 종자의 발아 특성은 모든 지표에서 차이가 있었다(p<0.05). 종자 저장 기간이 52일인 경우, 저장 종자의 발아율은 대조구와 유의한 차이가 없었으나, T50과 평균발아일수가 늘어나고 발아속도와 발아균일지수는 감소하여, 종자세가 감소한 것을 알 수 있었다. 4°에서 73일간 저장한 종자의 경우, 평균 발아율은 46.0%로, 대조구 종자의 초기 발아율인 81.0%의 57% 수준으로 감소하여, 활력이 크게 감소한 것으로 나타났다.
죽절초 종자의 건조 내성을 평가하기 위하여 실시한 건조 실험 결과, 함수율에 따른 발아 특성은 Table 4와 같았다. 건조하지 않은 채취 직후의 종자(함수율 28.1±1.4%)를 4°에서 52일 저장한 후, 수준별로 건조하여 발아 실험과 종자 침출수 분석을 실시한 결과, 발아균일지수를 제외한 모든 발아 특성에서 처리 간에 통계적으로 유의한 차이가 있었다(p<0.05). 먼저, 종자 함수율이 평균 18.6%인 처리 I에서는 발아율과 발아 속도가 감소하지 않았다. 그러나 함수율이 5.3%로 감소한 처리 II에서는 발아율이 20.0%로 크게 낮아지고, 2.5%로 감소한 처리 III에서는 활력을 거의 소실하였다. 이것은 채취 직후 종자의 함수율을 2.3%, 2.2%까지 조절한 후 조사한 발아율인 55%, 30%와 비교했을 때, 종자 활력이 더 빠르게 감소한 것을 확인할 수 있었다.
The seeds were dried for 0, 5, 20, and 48 hours in treatment control, I, II, and III, respectively.
Each field value represents mean±standard deviation (n=4).
Different letters in a column represent significant difference among treatments according to Tukey’s HSD tests at α = 0.05.
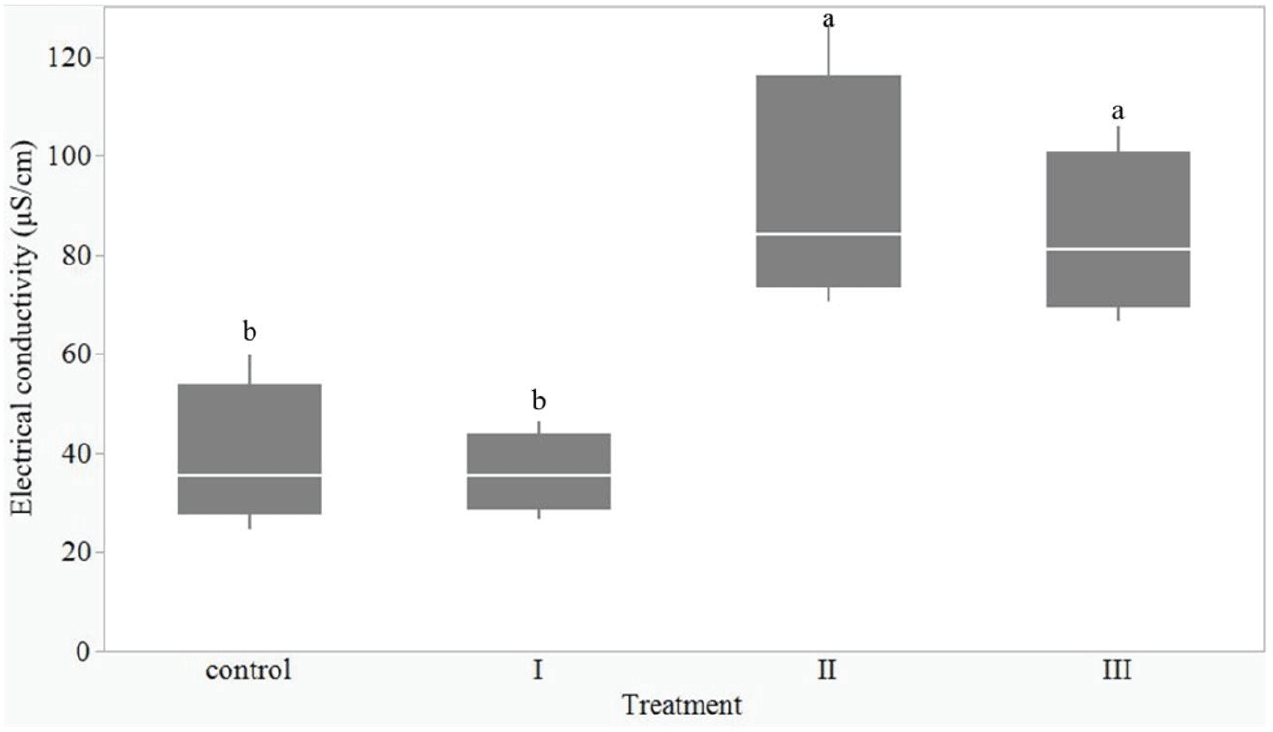
저장된 종자의 건조에 따른 종자 침출수의 전기전도도 분석 결과, 전기전도도에서 처리 간 차이가 뚜렷하게 나타났으며(p<0.05, Figure 1), 처리간 다중비교 결과, 두 그룹으로 분류되었다. 먼저, 처리 I은 종자 함수율이 감소하였으나 발아율과 발아 속도가 감소하지 않은 그룹으로, 전기전도도의 평균값은 36.0 μS/cm로 대조구(38.9 μS/cm)와 유의한 차이가 나타나지 않았다. 그러나 종자의 발아율과 발아속도가 크게 감소한 처리 II와 III은 침출수의 전기전도도 역시 각각 91.2 μS/cm, 83.8 μS/cm로 높은 값을 보였다. 종자 침출수의 무기질 구성을 분석한 결과, Na, Mg, K, Ca, Mn, Fe, Cu, Zn 중 Mn과 Zn은 검출되지 않았으며, K의 농도가 가장 높았다. 이 중 Na, K, Ca, Fe, Cu의 농도에서 처리 간 통계적인 차이가 있었고, Mg 농도는 처리 간 통계적인 차이가 없었다(p<0.05, Table 5). 농도가 가장 높았던 K와 Ca의 경우는 처리 II과 처리 III에서 대조구보다 더 높은 값을 보였으며, Cu 농도는 모든 처리구에서 대조구보다 높은 값을 보였다. 그러나 Fe 농도는 모든 처리구에서 대조구보다 낮은 값을 나타냈으며, Na 농도는 처리에 따른 뚜렷한 경향을 보이지 않았다.
The seeds were dried for 0, 5, 20, and 48 hours in treatment control, I, II, and III, respectively.
Each field value represents mean±standard deviation (n=4).
고 찰
종자를 이용한 증식과 복원을 위해서는 우선적으로 해당 수종 종자의 발아 특성에 대한 정보를 얻는 것이 필요하다. 죽절초 종자의 경우, 발아 전 처리 없이도 90% 이상 발아하는 것으로 알려져 있으나(Seo et al., 2006), 휴면이 없는 종자가 수 일 내 발아하는 것에 비해 죽절초 종자는 발아 개시에 2주 이상, 발아 종자의 50%가 발아하는 데 20일 이상 소요되어 깊지 않은 휴면이 있는 것으로 예상되었다. 깊지 않은 생리적 휴면(nondeep physiological dormancy)의 경우, 최소 5일에서 60~90일 동안의 저온습윤처리로 쉽게 타파할 수 있으나(Baskin and Baskin, 2014), 죽절초 종자의 발아 전처리 효과를 비교한 실험 결과, 30일 저온습윤처리가 발아율과 발아속도, 발아균일도 향상에 큰 효과가 없는 것으로 나타났다. 따라서 죽절초 종자의 휴면 원인이 생리적 휴면은 아닐 것으로 판단되며, 후속 연구에서 배의 형태를 관찰하여 형태적 휴면 여부를 밝히고 발아 속도를 높이는 처리를 탐색하는 것이 필요하다.
종자의 건조 내성의 정도를 파악하기 위해서는 주로 건조 처리를 통해 함수율을 여러 수준으로 조절하여 활력을 평가하는 방법이 이용된다(Hong and Ellis, 1996; Millennium seed bank, 2022). 죽절초 종자의 건조 수준에 따른 발아 특성을 조사한 결과, 종자의 탈종 직후의 함수율은 32.8%이며, 채취 직후 종자는 함수율이 5% 미만으로 감소하였을 때에도 활력과 종자세를 유지하였다. 반면, 저장된 종자는 함수율 18.3%로 건조되었을 때까지 활력과 종자세를 유지하였으나 함수율이 5.5%로 떨어질 경우 활력과 종자세가 크게 감소하였다. 또한, 추가적인 건조에 의해 함수율이 2% 대로 감소하자 활력을 거의 소실하였다. 채취 직후의 종자가 5% 미만의 낮은 함수율 수준에서도 활력을 잃지 않은 것은 건조에 의한 피해가 회복 불가능한 상태가 되기 전에 바로 수분이 공급되었기 때문인 것으로 생각된다. 그러나 저장된 종자의 경우, 초기 활력은 채취 직후 종자와 큰 차이가 나지 않았으나, 종자가 건조됨에 따라 활력이 급격히 감소하였는데, 이는 저장 기간 동안 세포 수준에서 노화가 시작된 종자에 건조에 의한 피해가 더해졌기 때문으로 보인다.
건조 내성을 가지는 세포는 건조 시 원형질막의 손상을 회복하는 기작이 있으나, 건조에 민감한 종자의 경우, 이러한 기작이 없어 건조 시 세포 원형질막의 기능과 구조에 손상을 입으므로 재수화 시 세포질 내부 용질의 용출이 일어나는 것으로 알려져 있다(Bewley et al., 2013). 이렇게 누출된 성분들이 종자 침출수의 전기전도도를 결정하므로, 침출수의 전기전도도를 분석함으로써 세포막 기능의 손상 정도를 간접적으로 확인할 수 있다. 본 연구에서 수준별로 건조한 종자를 침지하여 침출수를 분석한 결과, 더 많이 건조된 종자일수록 전기전도도가 높게 측정되는 것을 확인할 수 있었다. 이러한 결과는 건조에 의한 종자 활력의 저하가 세포막 손상과 연관이 있음을 암시한다. 침출수에서 분석한 무기질 중 가장 농도가 높았던 원소는 K였는데, 소나무(Gu et al., 2022), 참느릅나무(Kim et al., 2011), 대두(Dias et al., 1997), 목화(Woodstock et al., 1985) 등에 대한 연구에서도 종자 침출수에 포함된 무기원소가 주로 K인 것으로 보고된 바 있다. K는 세포의 삼투압을 조절하며 각종 효소의 활성제로 작용하는데, 식물이 가뭄, 염해, 병원균 등 각종 스트레스를 받았을 때 유출되는 전해질 중 대부분을 차지하는 것으로도 알려져 있다(Demidchik et al., 2014). 본 실험 결과, 지나친 건조로 발아율이 20% 미만으로 떨어진 종자의 경우, 침출수에서 검출된 K의 농도가 높게 나타나 건조에 의한 종자 활력 저하와 연관이 있는 것으로 추정되었다.
이상을 종합하면, 본 연구 범위에서 죽절초 종자가 활력을 잃지 않는 최소 함수율 수준은 채취 직후 종자에서 함수율 5%, 저장된 종자의 경우 함수율 18%였으며, 건조에 의해 활력 저하가 나타난 종자의 경우 세포막 기능이 손상된 것으로 나타났다. 통상 함수율 10~12.5% 수준으로 건조되는 것에 견디나 그 이상으로 건조되었을 때 활력을 잃는다면 이들은 중간저장성 종자로 여겨진다(Hong and Ellis, 1996). 따라서, 죽절초 종자의 건조 내성은 난저장성 종자에서 중간저장성 종자 사이의 특징을 가지는 것으로 판단되며, 건조 내성의 정도는 노화 정도에 따라 다를 수 있다.
종자의 수명은 노화되어 활력을 잃기 전까지의 기간이며, 같은 저장 환경에서도 수종에 따라 매우 다양하다. 일반적으로 실온에서 50% 이상의 발아율을 유지하는 것을 기준으로 수명이 2년 이내인 단명종자, 3~5년인 상명종자, 5년 이상인 장명종자로 구분한다(Kim et al., 2012; Kim et al., 2014). 종자 노화에 의한 증상은 먼저 종자세가 감소하여 유묘의 출현이 지연되고, 그 다음으로 발아율이 감소한다(Delouche and Caldwell, 1960). 죽절초 종자를 건조하지 않고 저장한 결과, 18°에서는 동결이 일어나고, 25°에서는 급격히 노화가 일어나 전혀 발아하지 않았다. 4°에 저장한 경우, 52일차에서 발아율은 감소하지 않았으나 종자세의 감소가 먼저 나타났으며, 73일차에서는 발아율이 큰 폭으로 감소하였다. 본 실험은 종자의 건조 특성에 관한 정보가 없는 상황에서 건조하지 않은 종자를 진공포장해서 저장한 결과이므로, 저장 환경을 다르게 할 경우 종자 수명이 달라질 수 있으나, 두 달 반 만에 활력이 절반 가까이 감소했으므로 죽절초 종자는 종자 수명이 수 개월 이내인 단명종자로 분류할 수 있다.
FAO(2014)의 진뱅크 표준에 따르면, 저장성 종자의 경우 주기적인 활력 평가를 통해 발아율이 초기 발아율의 85% 이하로 감소하기 전에 갱신이 이루어지는 것이 좋다. 본 연구 결과, 함수율을 조절하지 않고 4°에서 저장한 죽절초 종자의 경우 저장 기간이 73일이 되었을 때 발아율이 초기 발아율의 56.8%로 감소하였으므로, 이와 같은 저장 방법으로는 종자의 수명이 결실 주기보다 짧아 종자를 이용한 현지외 보존이 어려울 것으로 판단된다. 또한 종자 채집 후 파종 전까지 단기 저장할 경우에도 저장 기간을 최대한 짧게 해야 할 것으로 보인다.
종합하면, 죽절초 종자는 건조에 의해 활력을 잃기 쉬운 난저장성 종자 또는 중간저장성 종자이자 수명이 짧은 단명종자로 판단된다. 단명종자나 난저장성 종자는 기본적으로 종자를 이용한 장기 보존에 적합하지 않으며, 파종 전 단기적인 보관을 위한 습윤 저장 방법이나 배나 배축 등 기타 생식질의 초저온 보존 등을 이용한 장기 보존 방법을 개발해야 한다(Pritchard, 2004; FAO, 2014). 따라서 후속 연구를 통해 활력 감소를 최소화하는 종자 건조 기술 혹은 초저온 보존 등의 장기 보존 방법을 개발하는 것이 필요하다.