
서 론
박달목서(Osmanthus insularis Koidz.)는 물푸레나무과 목서속(Osmanthus)에 속하는 상록활엽교목으로, 우리나라, 일본, 대만에 걸쳐 분포한다. 박달목서는 박달나무에서 유래한 국명에서 알 수 있듯이 목질이 단단하며, 꽃향기가 좋고 개화기간이 길어 조경수로서의 가치가 높다(Yun, 2016). 우리나라에는 거문군도와 가거도, 제주도 일대에 서식하며, 2022년 기준 박달목서의 최대 자생지인 거문군도 일대에서 122개체가 확인된 것으로 보고된 바 있다(Chung et al., 2022). 이처럼 자생지가 도서지역으로 한정적이고 개체수가 적어 산림청 지정 희귀등급 위기종(EN, Endangered)으로 지정되어 있으므로, 생물종의 현지내·외 보존을 위한 대책 수립이 요구되는 수종 중 하나이다.
현지내 보존을 위한 자생지 생육 환경 개선, 현지외 보존원 조성 및 관리 또는 복원을 위해서는 해당 종의 생육 단계별 최적 생육 환경에 대한 정보가 뒷받침되어야 한다. 특히 유묘 단계의 생육 환경은 현지 내에서의 천연 갱신, 복원 재료 공급을 위한 양묘, 초기 활착 등을 결정짓는 중요한 요소이다.
식물의 생장과 생리에 영향을 미치는 환경 인자 중에서도 햇빛은 식물의 생장에 필요한 에너지를 공급할 뿐 아니라 각종 생리적 활동의 신호로 작용하며, 생장과 발달에도 영향을 미치는 주요 환경 인자이다. 식물은 발달과 생장, 잎의 형태적 특성, 광합성과 호흡 등 각종 대사 작용에 있어 햇빛을 효율적으로 이용하기 위해 다양한 형태의 적응을 하며(Jackson, 1967; Joggi et al., 1983; Boller and Nösberger, 1985; Abrams and Kubiske, 1990; Reich et al., 1998; Kami et al., 2010), 임분 내에서 햇빛에 대한 경쟁은 식물의 군집 구조에 영향을 준다(Wierman and Oliver, 1979; Ford and Diggle, 1981). 햇빛에 대한 반응은 수종별로 상이한데, 일반적으로 식물 생장에서 햇빛이 적은 환경에서는 햇빛을 더 확보하기 위해 줄기 생장이 촉진되지만, 직경 생장이 영향이 적다(Ballré et al., 1987; Franklin, 2008; Kami et al., 2010). 또한, 햇빛이 적은 환경에서는 수광량을 확보하기 위해 잎이 넓고 얇아지며, 반대로 햇빛이 풍부한 환경에서는 잎이 두껍고 무거우며, 책상조직과 해면조직이 치밀하게 발달해 엽록소 함량이 높아진다(Gonçalves et al., 2008; Liu and Su, 2016). 광합성 특성에 있어 내음성이 높은 음수의 경우, 광포화점은 낮으나, 광보상점이 낮고 호흡량이 적어 낮은 광도에서도 광합성을 효율적으로 실시할 수 있다(Lee, 2011). 생화학적으로는 광합성에서 암반응을 담당하는 탄소고정계의 효소 활성이 저하되고, 명반응을 담당하는 광화학계의 색소 함량과 활성이 증가함으로써 빛의 이용효율이 증가한다(Terashima and Hikosaka, 1995; Kim et al., 2001). 이러한 내음성 기작에 따라 음수나 중용수는 낮은 광도조건에 적응하여 생존 경쟁에서 유리한 위치를 차지하게 한다. 반면 내음성이 약한 양수의 경우, 음수보다 광포화점이 높아 광도가 높은 환경에서의 광합성 효율이 높으며 생장속도가 빠른 대신, 그늘에서는 엽록소 함량 감소 및 광합성의 저하로 생장이 감소한다(Loach, 1967; Osunkoya et al., 1994; Cho et al., 2008). 이러한 내음성의 정도와 광에 대한 반응은 동일 속 내에서도 수종별로 차이가 있을 수 있다. 목서(Osmanthus) 속의 광에 대한 생육 반응에 대한 기존 연구에서, 구골나무(O. heterophyllus) 삽목묘는 저광도에서 생존이 어려웠으며, 광도가 높을수록 수고 및 잎 수가 증가하였다는 보고가 있으며(Song et al., 2012), 금목서(O. fragrans var. aurantiacus) 유묘의 경우, 차광률 75%까지는 양호한 생육을 나타내었으나, 차광률 95%에서는 생육이 불량하였다고 보고된 바 있다(Choi et al., 2012). 이렇듯 수종에 따라 내음성의 정도와 생육에 필요한 광 조건이 다르므로, 수종에 맞는 적절한 생육 관리를 위해서는 광에 대한 형태적, 생리적 반응에 대한 정보가 필수적이다.
따라서 본 연구는 희귀 식물인 박달목서의 생육 환경에 따른 초기 생장 및 생리적 특성을 분석함으로써 현지내⋅외 보존 전략을 수립하기 위한 기초 자료를 제공하고자 수행되었다. 이를 위해 차광 처리를 통해 4수준의 광조건을 조성하고, 박달목서 유묘의 생장 특성, 잎 형태, 광합성 특성, 광합성 색소 함량을 조사하였다.
재료 및 방법
본 연구는 경기도 수원시 권선구 오목천동에 위치한 국립산림과학원 산림생명자원연구부 야외 묘포장(37°15'03.0"N, 126°57'19.7"E)에서 수행되었다. 공시수종인 박달목서는 2022년 6월 거문도에서 종자를 채집하여 생장상에서 발아시킨 후 포트로 옮겨 심고, 시험 전까지 온실에서 동일한 환경 조건 하에 관리하였다. 2023년 4월에 그 중 생장이 균일한 48개체를 선별하여 처리별로 12개체의 유묘를 20 cm 간격으로 묘포장에 식재하였다. 식재 당시 유묘의 초기 수고는 4.3±1.5 cm, 근원경은 1.85±0.34 mm였으며(n=48), 초기 수고와 근원경의 처리구 간 통계적인 차이가 없었다. 생육이 활발하게 이루어진 2023년 5월부터 9월까지 묘포장의 최저 기온은 5.1℃, 최고 기온은 34.9℃, 월평균 강수량은 215.7 mm였다. 식재한 묘목의 관수는 1주일 간격으로 실시하였고, 유묘의 생육 환경을 보호하기 위하여 주기적으로 제초작업을 실시하였다. 차광 처리는 0%(대조구), 30%, 50%, 70% 차광막을 이용하여 실시하였으며, 상대 광량을 측정하기 위하여 각 처리구별로 광 센서 및 데이터 로거(HOBO, Onset Computer Corporation, USA)를 설치하였다. 상대 광량은 5월부터 9월까지 각 처리구의 광량을 측정하여 평균을 구한 후 대조구인 차광 0%에 대한 비율을 계산하였으며, 순서대로 100%(전광), 55%, 20%, 10%로 나타냈다.
각 처리구의 모든 묘목을 대상으로 생장 특성을 분석하기 위하여 2023년 4월에 초기 수고와 근원경을 측정하였고, 생장이 종료된 11월에 최종 수고와 근원경을 측정하였다. 그 결과를 이용하여 상대 생장률(relative growth rate)을 [Ln(x2)-Ln(x1)]/(t2-t1)의 식에 의해 계산하였다(Beadle, 1993). 여기서 x2와 x1은 생장 종료 후(t2)와 이식 초기(t1)의 수고를 나타낸다.
또한, 생육이 종료된 것으로 판단되는 11월에 처리별로 4개체를 굴취하여, 잎, 줄기, 뿌리로 구분하고 세척하여, 65℃에서 48시간 동안 건조시킨 후 건중량을 측정하였으며, 이를 이용하여 수목의 지상부와 지하부의 비(S/R)를 구하였다.
광 처리구에 따른 잎 형태 변화를 확인하기 위하여, 처리구별로 개체별 잎 수, 채취한 각각 잎의 면적, 두께 및 무게를 측정하였다. 잎 면적은 엽면적 측정기(LI-3100 Area meter, LI-COR)으로 측정하였으며, 각각의 잎 면적을 생중량으로 나누어 잎면적비(specific leaf area, SLA)을 계산하였다.
광합성은 생장이 활발하게 이루어지고 있는 시기인 8월 말부터 9월 초에 오전 9시부터 오후 5시 사이에 휴대용 광합성 측정기(LI-6800, LI-COR)를 사용하여 측정하였다. 장비의 측정 환경은 비가 오지 않는 맑은 날에 챔버 내의 상대습도를 60%, 공기의 온도를 24∼28℃로 설정하여 측정하였다. 광합성 측정은 각 처리구별로 완전히 성숙한 잎을 보유한 3개체를 선정하고, 각 잎을 대상으로 광도를 0, 25, 50, 100, 250, 500, 750, 1000, 1200, 1500μmol m-2 s-1로 조절하면서 각 광량에 따른 광합성 속도를 3반복으로 측정한 후 평균값을 이용하였다. 이 측정값을 이용하여 각각의 광-광합성곡선(light curve)을 작성한 후, 암호흡 속도, 순양자수율, 광보상점을 산출하였다. 또한 광포화점에서의 광합성 속도, 기공전도도, 증산속도 및 수분이용효율(Instantaneous water use efficiency, Ripullone et al., 2004)을 구하여 각각 처리간 차이를 비교하였다.
잎 내 광합성 색소 함량 측정은 dimethyl sulphoxide (DMSO)를 이용하여 추출하는 Hiscox와 Israelstam(1979)의 방법을 사용하였다. 광합성 색소 분석을 위한 잎 시료는 각 처리구에서 광합성을 측정했던 3개체로부터 채취하였다. 각 처리구별로 시료 채취기(0.785 cm2)를 이용하여 개체별로 원형 잎 시료 3개를 무작위로 채취하였으며, 채취한 잎에 DMSO 10 mL를 첨가하고, 70℃의 항온 수조에서 2시간 동안 유지하여 색소를 추출하였다. 추출액은 470, 645, 663 nm에서 흡광도를 3반복으로 측정하여 Lichtenthaler (1987)의 방법으로 엽록소 a, 엽록소 b, 카로테노이드의 함량을 계산하였다.
결 과
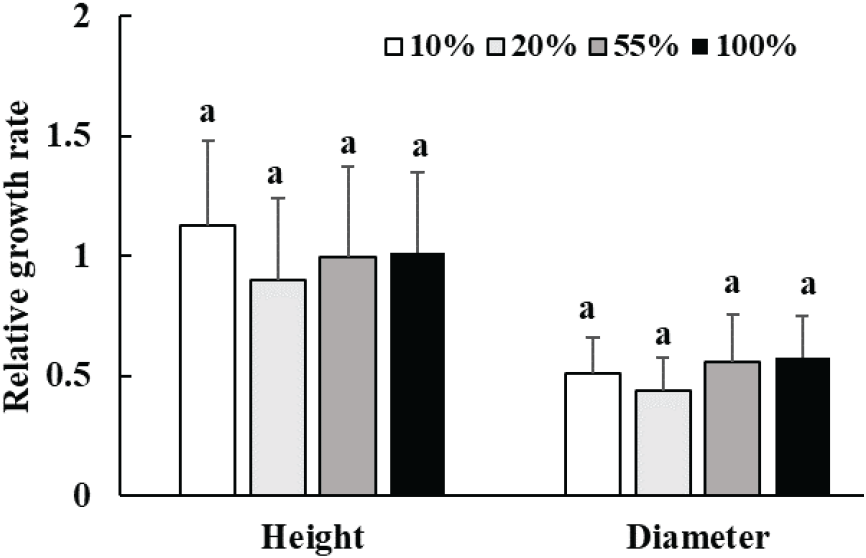
광량에 따른 박달목서 유묘의 생장 특성을 파악하기 위해 수고와 근원경의 상대 생장률을 분석하여 Figure 1과 같이 나타내었다. 박달목서 유묘의 수고와 근원경의 상대 생장률은 광량에 따른 통계적인 차이를 보이지 않았다. 그러나 수고의 상대 생장률은 10% 상대 광량에서 0.51로 가장 높았으며, 20% 상대 광량에서 가장 낮은 0.90를 보였으며, 근원경의 상대 생장률은 55%와 100% 상대 광량에서 각각 0.56, 0.58로 높은 값을 나타냈다.
수고와 근원경의 생장과는 달리, 박달목서 유묘의 건중량은 광량에 따른 차이를 보였다(Table 1, p<0.05). 박달목서 유묘의 총 건중량은 상대 광량 55%에서 가장 높은 2.95g을 보였으며, 20% 상대 광량에서 가장 낮은 0.90g을 나타냈다. 잎, 줄기, 뿌리의 건중량 역시 55% 상대 광량에서 가장 높았으며, 지상부와 지하부의 비인 SR 비율 역시 55% 상대 광량에서 4.58로 가장 높았다.
All the values are means ± SD of four replicates. Values with different letters within the columns indicate statistical differences among four light intesities at the 5% levels by Duncan’s multiple range tests.
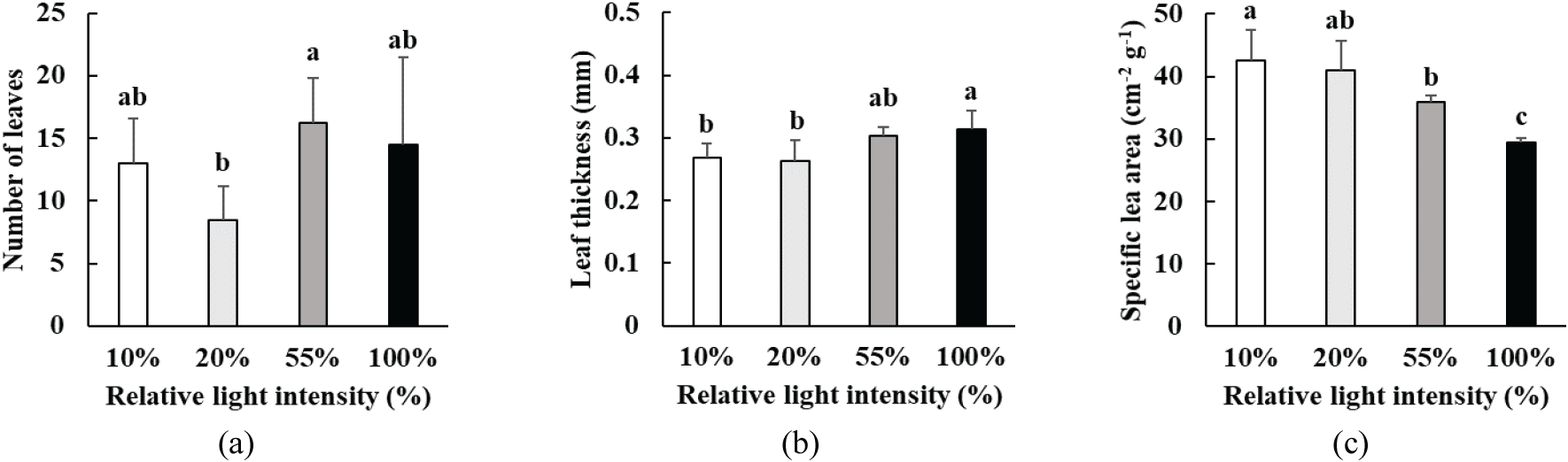
광량에 따른 박달목서 유묘의 잎 형태 특성 변화는 Figure 2와 같았다. 박달목서 유묘의 잎 형태 특성은 광량에 따라 뚜렷한 차이를 보였다(p<0.05). 박달목서의 잎 수는 55% 상대 광량에서 16개로 가장 많았으며[Figure 2(a)], 잎 두께는 상대 광량 10%에서 가장 얇았고, 광량이 증가함에 따라 점차 증가하여 상대 광량 100%에서 가장 두꺼웠다[Figure 2(b)]. 잎 면적비는 상대 광량 10%에서 42.5로 가장 높았으며, 광량이 증가함에 따라 점차 감소하여 상대 광량 100%에서 29.5로 가장 낮았다[Figure 2(c)].
박달목서 유묘의 광합성 특성은 광량에 따라 뚜렷한 차이를 보였다(Table 2, Figure 3, p<0.05). 광합성 속도는 55% 상대 광량에서 가장 높은 값인 6.5 μmol m-2 s-1를 나타냈고, 10% 상대 광량에서 가장 낮았다. 암호흡 속도 역시 55% 상대 광량에서 가장 높았고, 10% 상대 광량에서 가장 낮았다. 순 양자수율 또한 광합성 속도와 암호흡 속도와 마찬가지로 55% 상대 광량에서 가장 높았고, 10% 상대 광량에서 가장 낮았다. 광보상점은 광량에 따른 차이를 보이지 않았다. 그러나 광보상점은 광량에 따라 증가하는 경향을 보였으며, 10% 상대 광량에 가장 낮았고, 100% 상대 광량에서 가장 높았다.
All the values are means ± SD of three replicates. Values with different letters within the columns indicate statistical differences among four light intensities at the 5% levels by Duncan’s multiple range tests.
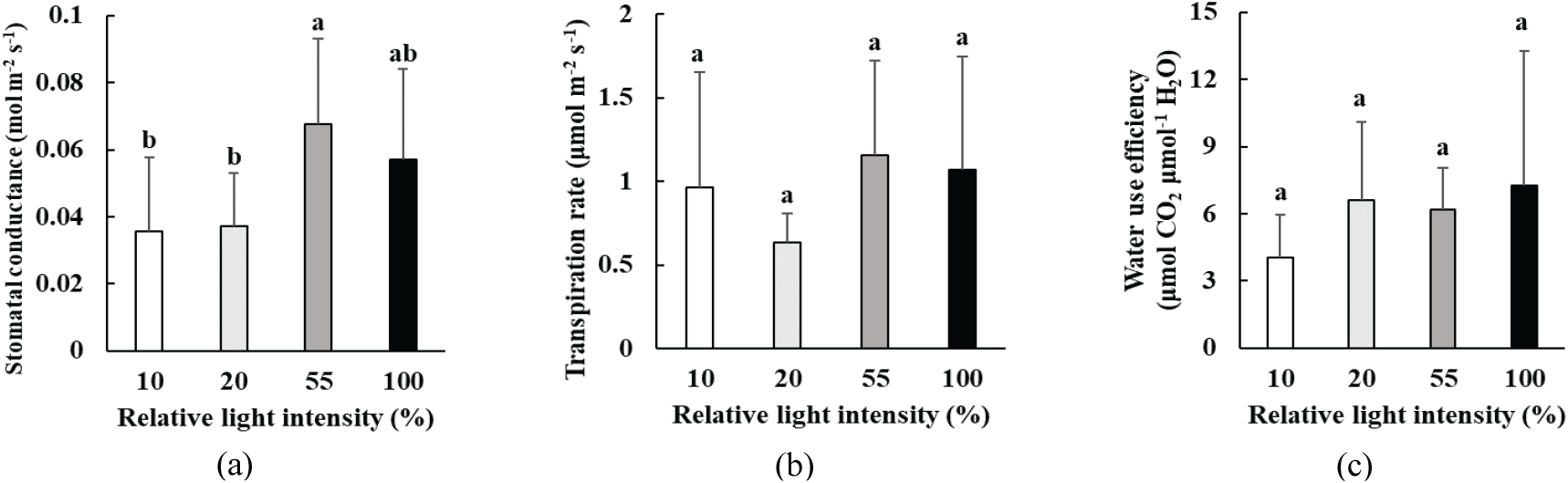
박달목서 유묘의 잎의 기공전도도는 55% 상대 광량에서 가장 높은 0.068 mol H2O m-2 s-1이었으며, 10%, 20% 상대 광량에서 낮은 값을 나타냈다[Figure 3(a)]. 증산속도는 광량에 따른 유의한 차이는 없었으나, 20% 상대 광량에서 가장 낮은 값을 나타냈고, 55%와 100% 상대 광량에서 비교적 높은 값을 보였다[Figure 3(b)]. 수분이용효율 역시 광량에 따른 통계적 차이가 없었으며, 10% 상대 광량에서 가장 낮은 값을, 전광에서 가장 높은 값을 보였다[Figure 3(c)].
박달목서 유묘의 잎내 광합성 색소 함량은 광량에 따라 뚜렷한 차이를 보였다(Table 3, p<0.05). 엽록소 a, b 및 총 엽록소 함량은 55% 상대 광량에서 가장 높았으며, 100% 광에서 가장 낮았다. 카로테노이드 함량 역시 55% 상대 광량에서 가장 높은 8.23 μg cm-2을 나타냈고, 10% 상대 광량에서 가장 낮은 3.87 μg cm-2를 나타냈다. 엽록소 a와 b의 비는 55% 상대 광량에서 가장 높은 2.56이었고, 10% 상대 광량에서 가장 낮은 2.22를 나타냈다. 총엽록소와 카로테노이드 함량의 비는 10% 상대 광량에서 가장 높은 5.78이었으며, 100% 상대 광량에서 가장 낮은 4.50을 나타냈다.
All the values are means ± SD of three replicates. Values with different letters within the row indicate statistical differences among four light intensities at the 5% levels by Duncan’s multiple range tests.
고 찰
식물은 광조건에 따라 줄기·직경 생장, 잎의 형태, 광합성 특성, 생리 활성 등에서 수종에 따라 다양한 적응 반응을 보인다(Franklin, 2008; Morales and Kaiser, 2020). 일반적으로 식물은 광이 적은 환경에서는 수광량을 확보하기 위해 잎의 면적을 증가시키며(Pearcy and Sims, 1994; Niinemets, 1998; Liu and Su, 2016), 반대로 광이 풍부한 환경에서는 수분 손실을 줄이기 위해 잎이 작고 두꺼워지고, 책상조직과 해면조직이 발달해 광 이용 효율을 높이는 것으로 알려져 있다(Gonçalves et al., 2008; Liu and Su, 2016; Gjindali and Johnson, 2023). 박달목서 유묘의 잎 형태는 이러한 경향을 잘 따랐으며, 광 조건에 맞게 적응한 것을 알 수 있다. 그러나 박달목서 유묘의 광량에 따른 수고와 근원경의 상대 생장률 차이를 비교한 결과, 절대적인 수치는 수고의 경우 10% 상대 광량에서, 근원경은 55%, 100%일 때 다소 높게 나타났으나, 수고와 근원경에서는 통계적인 차이가 나타나지 않았다. 이는 식재 후 1년차 생장에 대한 결과이므로 줄기, 직경 생장에 대한 광의 효과가 충분히 나타나지 않았기 때문으로 보인다.
유묘의 잎, 줄기, 뿌리의 건중량과 SR 비율은 55% 상대 광량에서 가장 높게 나타났는데, 이는 해당 광 조건에서 잎 수가 가장 많고 뿌리 발달이 좋았기 때문이다. 건중량은 생리 대사에 따른 바이오매스 축적의 최종 결과로서, 해당 광 조건에서 가장 효율적으로 광을 이용하여 생장하였다는 것을 의미한다.
광합성 특성과 엽록소 함량 측정 결과는 55% 상대 광량 조건에서 광합성이 가장 활발하게 이루어졌음을 보여주었으며, 이는 해당 광량에서 가장 높은 건중량을 보인 결과를 뒷받침한다. 먼저, 광포화점에서의 광합성 속도와 순양자수율이 55% 상대 광량에서 가장 높게 나타났다. 순양자수율은 광-광합성 곡선의 초기 기울기이며, 약광 조건에서 광화학계의 활성을 나타내는 지표이다(Kim and Lee, 2001). 따라서 강광, 약광 조건에서 모두 광합성이 활발하게 이루어졌음을 알 수 있다. 또한, 빛 에너지를 받아들이는 엽록소 a, b와 보조색소인 카로테노이드 함량 역시 가장 높게 나타나, 빛의 포집 능력 역시 가장 좋았다. 이러한 특성들로 보아 55% 상대 광량에서 광합성을 통한 동화작용이 가장 활발하게 일어났음을 알 수 있다.
또한 55% 상대 광량 조건에서는 기공전도도, 증산 속도, 암호흡 속도 역시 가장 높게 나타났다. 기공전도도와 증산 속도가 높다는 것은 기공을 통한 가스 교환과 증산이 활발하게 일어났다는 것을 뜻한다. 또한 증산으로 인한 수분 손실이 있었음에도 불구하고 수분이용효율이 높았던 것으로 보아, 광합성을 위한 이산화탄소 공급이 원활하게 이루어진 것으로 보인다. 암호흡은 생명 유지와 대사에 필요한 에너지를 공급하는 과정이며, 특히 어린 식물의 생장에 기여하는 바가 크다(Ryan et al., 1994). 암호흡이 감소하면 광합성 능력과 생장 속도 역시 저하될 수 있다(Kim et al., 2001). 따라서 55% 상대 광량에서 박달목서 유묘의 암호흡 역시 가장 활발하게 일어났기 때문에 광합성 능력과 물질 합성에 이용할 수 있는 에너지를 충분히 생산하였음을 알 수 있다.
반면, 전광 조건에서는 10%, 20% 상대 광량에서보다 광합성 속도, 암호흡 속도가 높았으나, 55% 상대 광량보다는 낮은 값을 보였다. 일반적으로 햇빛이 강해지면, 식물의 잎은 수분 손실을 막기 위해 크기가 작아지는 대신 책상조직을 치밀하게 발달시키고, 단위 면적 당 엽록소 함량을 늘리는 것으로 알려져 있다(Lee, 2011; Morales and Kaiser, 2020). 그러나 박달목서 유묘의 경우, 전광 조건에서 잎은 작고 두꺼워졌으나 엽록소 a, b의 함량은 모든 처리구 중에 가장 낮게 나타났는데, 이는 잎이 너무 강한 햇빛에 노출되어 엽록소의 광산화가 일어났기 때문으로 보인다(Lee et al., 2014; Sato et al., 2015; Luo et al., 2022). 더불어 전광 조건에서 엽록소/카로테노이드 비율이 가장 낮게 나타났는데, 이는 엽록소에 대한 카로테노이드의 상대적인 함량이 높았다는 것을 의미한다. 카로테노이드는 세포에서 에너지 소산, 자유라디컬 소거, 원형질막 손상 방지 등을 통해 광산화를 방지하는 역할을 한다(Bramley, 2013; Sun et al., 2022). 즉, 높은 광 조건에서 광계 II와 엽록소의 손상을 방지하기 위해 카로테노이드의 상대적인 함량이 높아진 것으로 보인다(Izuhara et al., 2020). 따라서 전광 조건에서 생육한 박달목서 유묘는 강한 햇빛에 의해 엽록소에 손상을 입어 55% 상대 광량 조건에 비해 낮은 광합성 능력을 보였고, 이에 최종 산물인 건중량도 더 낮았던 것으로 보인다.
또한, 전광에 노출된 박달목서 유묘는 기공전도도와 증산 속도가 55% 상대 광량보다 낮아지면서 수분이용효율이 가장 높게 나타났다. 이것은 전광 조건에서 증산에 의한 수분 손실을 막기 위해 잎의 기공을 55% 상대 광량 조건보다 더 많이 닫았기 때문인 것으로 보인다. 식물은 햇빛에 반응하여 기공을 열지만(Inoue and Kinoshita, 2017), 세포 내 CO2 농도가 높거나 광이 너무 강하면 수분이이용효율을 높이기 위해 기공을 닫기도 한다(Han et al., 2011; Lee, 2016; Yang et al., 2020).
차광 처리에 따른 생리적 특성 변화는 해당 수종의 내음성에 대한 정보를 제공한다. 기존 연구들을 살펴보면, 양수인 이태리포플러의 경우 약광 조건에서 엽록소 함량과 광합성 효율이 감소한 것에 비해, 음수로 알려진 음나무와 졸참나무는 이와 반대로 엽록소 함량과 광합성 효율을 높이는 적응 반응을 보였다(Kim et al., 2001). 극음수로 알려진 굴거리나무 유묘 역시 피음 수준이 높아질수록 엽록소 함량이 높아졌으며, 약한 피음 수준에서 광합성 속도가 가장 높았다(Song and Kim, 2017). 이와 같이 내음성이 강한 수종들은 그늘에서 생장할 때 엽록소 함량 및 카로테노이드와 같은 광합성 보조 색소의 함량을 증가시킴으로써 광합성 효율을 높인다(Han et al., 2011).
그러나 박달목서 유묘의 경우 차광 처리구 내에서 상대 광량이 낮아질수록 엽록소 a, 엽록소 b, 카로테노이드의 함량이 낮아졌으며, 광포화점에서의 광합성 속도와 순양자수율 역시 감소하였으므로 이와 같은 적응이 나타나지 않은 것으로 보인다. 뿐만 아니라 암호흡 역시 차광 처리구 내에서 광량이 낮아짐에 따라 감소하는 경향을 보였는데, 기존 연구에 따르면 식물이 그늘에서 자랄 때 낮아진 대사 활성에 대한 적응 반응으로 암호흡을 감소시키는 경우가 있다(Loach, 1967; Villar et al., 1995; Lusk and Reich, 2000). 따라서 박달목서 유묘는 광이 적은 환경에서 광합성 뿐 아니라 암호흡 속도를 낮추어 대사 활성과 생장 속도가 느려진 것으로 보인다.
이처럼 박달목서 유묘는 차광 수준이 높아질수록 광합성 색소 함량과 광합성 효율이 감소하는 경향을 보였으므로, 내음성이 비교적 약한 것으로 보인다. 그러나 일부 그늘에 대한 적응 반응을 보인 경우도 있었다. 먼저 광-광합성곡선에서 광합성량이 호흡량과 일치하여 순광합성량이 0이 되는 광도인 광보상점은 광량이 낮을수록 감소하는 경향을 보였는데, 낮은 광 조건에서 광합성을 할 수 있는 최소 광량인 광보상점을 낮추는 것은 그늘에서 자라기 위한 식물의 적응 반응 중 하나로 볼 수 있다(Lee, 2011). 또한 광량이 적어짐에 따라 엽록소 a/b 값이 감소하는 경향이 있었다. 이는 낮은 광도에서 생육하였을 때 엽록소 a, b의 절대적인 함량은 감소했지만, 그 감소폭은 엽록소 b에서 더 완만했으며, 결과적으로 엽록소 a에 대한 엽록소 b의 상대적인 비율은 증가하였다는 것을 보여준다. 엽록소 b는 광계 II의 반응단백질인 light harvesting chl-protein complex II의 함량과 상관성이 높은 색소로, 음수의 경우 낮은 광도에서 엽록소 b 함량의 상대적인 비율이 높아져 엽록소 a/b 값이 감소하는 것으로 알려져 있다(Terashima and Hikosaka, 1995; Kim et al., 2001; Luo et al., 2022). 그럼에도 불구하고 낮은 광 조건에서 실제 광합성 능력은 증가하지 않았으므로, 이러한 특징들이 광합성 효율을 높이는 데 실제적인 영향을 미치지는 않은 것으로 보인다.
한편, 20% 상대 광량에서 잎의 광합성 능력은 10%보다 높았음에도 불구하고 수고, 근원경, 건중량이 가장 낮게 나타났다. 이것은 20% 상대 광량에서 개별 잎의 광합성 속도를 포함한 생리적 특성은 광량이 높아짐에 따라 증가하였지만, 개체 당 잎 생산이 10% 상대 광량보다 적었기 때문에 총 광합성량이 적어 바이오매스 생산량에 영향을 미친 것으로 보인다.
결론적으로 박달목서 유묘의 경우, 수고와 근원경은 광량에 영향을 받지 않으며, 잎은 광량에 따라 형태적인 적응을 보였다. 그러나 그늘에서 광합성 효율을 높이는 생리적인 적응 반응은 뚜렷하지 않았다. 또한, 유묘 단계의 생육에 가장 적절한 광량은 전광의 55% 수준이며, 이 조건에서 광합성 활성이 가장 높고 최종적으로 건중량 생산이 최대가 되는 것으로 나타났다. 따라서 희귀 수종인 박달목서의 복원 또는 양묘를 위한 생육 환경 조성 시 유묘 단계에서는 중간 수준의 차광 처리를 하는 것이 가장 좋으며, 너무 강한 광이나 피음은 묘목의 생장과 광합성을 저해할 수 있을 것으로 판단된다.