
서 론
송이버섯(Tricholoma matsutake, T. magnivelare, etc.)은 전 세계적으로 주로 소나무과(Pinaceae) 수목과 공생하는 균근성 버섯인데, 우리나라 송이(T. matsutake)는 대부분 소나무(Pinus densiflora)와 공생관계를 맺으며 자실체를 형성한다. 즉, 우리나라 송이는 소나무와 밀접한 관련이 있는데, 우리나라 산림에서 소나무림은 줄어들고 있다. 국가산림자원조사 결과를 분석한 바에 따르면, 소나무가 대표 수종이었던 곳이 10년 후에도 그대로 유지된 비율은 75%이었고, 나머지 25%는 혼효림으로 전환된 것으로 나타났다(Kim et al., 2024). 또한, 향후 10년간 현재의 소나무 순림 14.2%가 혼효림으로 전환될 것으로 추정되었다. 이러한 변화는 수분 조건이 좋으며 임분 교란 요인이 많은 지역에서 더 뚜렷하게 나타나는데, 지역적으로 보면 소나무림 변화율은 제주와 경기가 각각 42.8%와 26.9%로 높고, 경북과 강원이 각각 8.8%와 13.8%로 낮았다. 즉, 미래에는 한반도 남서쪽 지역 소나무림은 거의 소멸하는 것으로 예측되었는데, 이러한 현상은 송이 생산지의 변화를 낳을 것으로 예상된다.
최근에는 산불로 인하여 소나무림이 소실되는 경우도 많아졌는데, 특히, 2000년과 2022년에는 송이 주산지인 강원도와 경상북도 일대에서 20,000 ha가 넘는 대형 산불 피해를 겪었다. 더구나 활엽수림에 비하여 소나무림이 산불에 더 민감한 것으로 평가되므로, 최근의 빈번한 산불은 우리나라 송이 생산량 감소에 직접적인 원인이 될 수 있을 것으로 추정된다. 아울러, 산불 피해를 겪은 송이 생산림의 복원을 위해 산불에 민감한 소나무를 재조림하는 것에 대한 비판적인 시각도 있다. 이러한 측면에서 산불이 송이 생산량 변화에 미치는 영향이나 산불 피해 송이 생산림의 복원 방법에 대한 검토가 필요하다.
최근에는 채취방식에 의존하는 송이 생산을 재배 체제로 전환하려는 노력이 활발히 전개되고 있다. Ka et al. (2006; 2018)은 기존 송이 균환을 이용한 감염묘(感染苗) 생산기법으로 송이 미발생 소나무림을 송이 발생림으로 변화시키는 것이 가능함을 보고한 바 있다. 하지만, 산업적인 활용을 위해서는 송이 감염 묘목 생산기법의 한계를 극복할 수 있는 접종묘(接種苗) 생산 및 활용이 필요함이 지적되고 있다. 그런데 송이 생장이 매우 느려서 접종묘 생산 및 육성단계에 어려움이 많으므로, 이를 극복하기 위한 시도가 전개되고 있다. 특히, 송이 균환 부위에 존재하는 다양한 생물의 상호관계 연구가 활발히 진행되고 있는데, 송이 균환에서 분리·배양한 균이 송이 균사 생장을 촉진한다는 보고(Oh and Lim, 2018; Oh et al., 2018; Choi et al., 2023; 2024)는 인공 재배에 청신호가 되고 있다. 이와 같은 송이 균환부 마이크로바이옴(microbiome) 연구의 진척도나 나갈 방향에 대한 검토는 송이 재배를 실현하는 초석이 될 것이다.
본 논문은 앞서 제기한 각 항목에 대하여 통계자료 및 문헌조사를 통해 우리나라 송이의 생산성 변화와 그 원인을 검토하고 앞으로 나아갈 방향에 대하여 고찰한 것이다. 소나무림의 분포 변화가 송이 생산성에 미치는 영향을 살펴보고, 송이 생산성을 유지·증진하기 위해서 어떤 조치가 필요한지 검토하였다. 아울러, 최근의 송이 연구를 고찰하며 미래 송이 연구의 방향성을 제시하고자 한다.
자료수집 및 분석방법
우리나라에서 송이버섯은 1970년부터 2006년까지 임산물 유통 제한 고시에 의거하여 산림조합에서만 수매, 유통될 수 있었다. 따라서 산림조합중앙회가 보유한 송이 공판량 자료는 우리나라 송이 생산량을 추론할 수 있는 매우 정확한 통계자료이었다. 그러나 규제 완화와 더불어 현재는 산림조합 이외의 유통망을 통해 자유롭게 유통될 수 있으므로 정확한 실태를 파악하기 힘들다. 국가산림통계로 제시되는 송이 생산량은 임산물 생산 조사 결과를 활용하는데, 송이는 산림조합 수매 이외에 일반 수집상에 의하여 수집·유통되는 양이 상당히 많으므로 산림조합 공판장과 송이 수집상을 대상으로 구분 조사한다. 그런데, 때로는 전국 생산량이 산림조합의 공판실적보다 적게 보고되기도 하여 통계의 신뢰도가 높지 않은 편이다. 이에 따라 본 연구에서는 정확한 수치를 파악하기 어렵다고 판단하여 생산량 변화 경향만 파악하기로 하였으며, 산림조합의 공판실적만 한정하여 자료로 활용하였다.
산불에 의한 소나무림 감소가 송이 생산량 변화에 직접적인 영향을 미치는지에 대한 검토는 2020년대 산불 피해를 겪은 강원도 홍천, 인제, 삼척 및 경상북도 울진의 산불 피해지와 송이 생산량의 상관성을 분석하였다. 산불 피해 이전 송이 수매량과 산불 피해 당년 및 이후의 송이 수매량을 비교하며 산불이 송이 생산량에 직접적인 영향이 있었는지 파악하였다.
국가과학기술정보센터의 SCIENCE ON (https://scienceon.kisti.re.kr/main/mainForm.do) 시스템을 활용하여 2024년 5월 현재 송이(Tricholoma matsutake)로 검색되는 논문 171개를 찾았다. 논문 제목을 확인하며 송이버섯의 화학적 성분이나 식·약용 가치 및 활용과 관련되는 논문은 제외하고, 송이산 및 송이균의 생태, 버섯 생산 관련 논문을 일일이 검토하였다.
본 논문은 학술지에 게재되는 것임을 고려하여 문헌은 30편 이내로 한정하여 국내 학회지에 발표된 논문 위주로 검토하였으며, 연구의 당위성이나 관련성을 설명하는 필수적인 외국 논문은 추가로 확인하며 고찰하였다. 특히, 최근 우리나라의 송이 연구 동향을 파악하기 위해서는 국내에서 연구가 진행된 연구자료로 한정하여 논문을 검토하였다.
결과 및 고찰
우리나라에서 송이는 1970년대에는 남한의 거의 전 지역, 2000년대 초반에는 태백산맥과 소백산맥, 그리고 노령산맥 주변의 49개 시·군 산림조합에서 수매되었다. 하지만, 이제 한반도 남서 지역에서는 송이 생산이 급격히 줄어 해당 지역 산림조합에서는 수매조차 하지 않고 있다(Figure 1). 산림조합 중앙회에서 제공하는 최근 5년간의 송이 수매 기록에 따르면, 강원 7개 지역(홍천, 인제, 고성, 양양, 삼척, 양구, 강릉), 경북 12개 지역(영덕, 울진, 봉화, 문경, 안동, 의성, 청송, 청도, 상주, 예천, 영주, 포항), 경남 1개 지역(거창)이 현재의 송이 주산지로 평가된다. 경북이 강원, 경남 지역보다 송이 생산량이 많았는데, 높은 수매량을 기록한 시·군 산림조합 1~5위가 모두 경북이었다(Table 1). 영덕은 5년간 송이 총 수매량이 107톤으로 가장 높았으며, 이어서 청송과 청도, 울진, 안동 순으로 송이 수매량이 많았다.
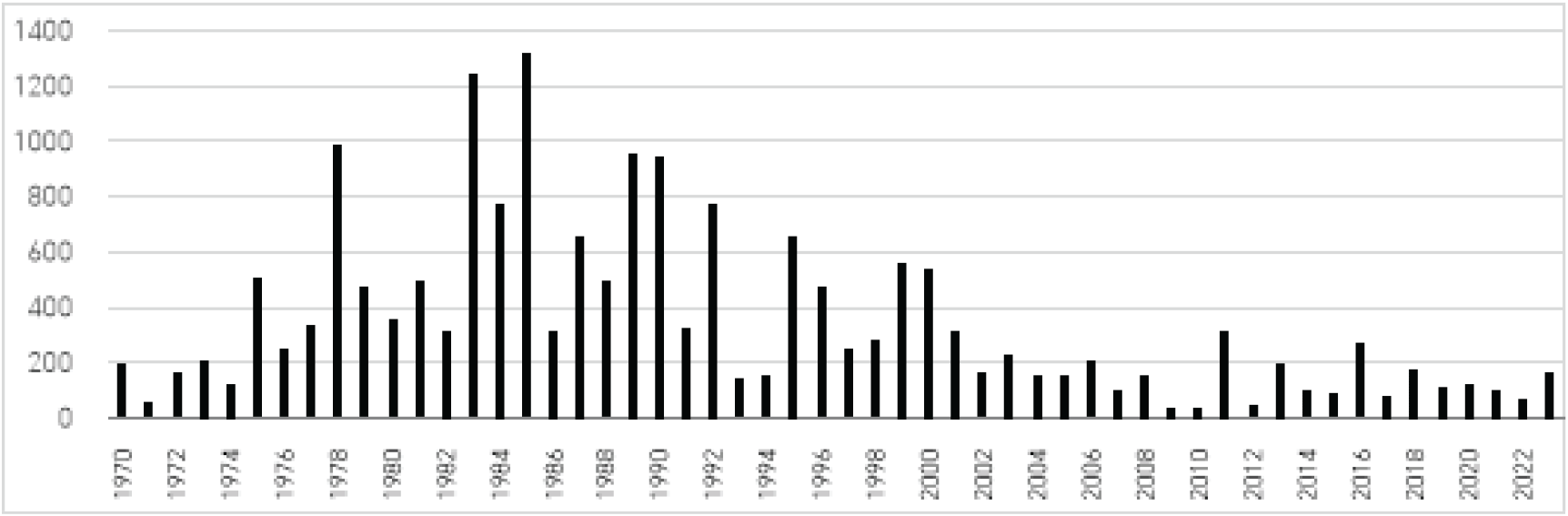
지역에 따라 산림조합에서 수매되는 송이의 비율이 다르므로 이 자료가 우리나라 송이 생산에 대한 명확한 정보를 제공한다고 할 수는 없다. 특히, 제1호 지리적표시제 임산물로 등록하여 송이를 유통하고 있는 강원도 양양은 산림조합 이외의 경로를 통해 송이가 유통되고 있으므로 생산량이 훨씬 더 많은 것으로 추정된다. 이러한 점을 고려하더라도 현 시점의 대한민국 송이 주산지는 태백산맥과 소백산맥 주변에 한정되는 추세임을 알 수 있다.
최근에는 산불로 인하여 소나무림이 소실되는 경우가 많아졌는데, 특히, 21세기에 들어서면서 송이 주산지인 강원도와 경상북도 지역에서는 대형 산불 피해를 겪었다. 이에 따라 해당 지역의 송이 생산량에 직접적인 영향이 있었는지 파악하기 위하여 강원도 5개 시·군(홍천, 인제, 고성, 양양, 삼척)의 최근 자료를 검토하였다. 강원도 지역의 경우, 영서(홍천, 인제)와 영동(고성, 양양, 삼척)의 지역 특성 또는 산불 규모에 따라 영향력을 검토하였는데, 일정한 경향성을 찾기 어려웠다.
영서 지역인 홍천의 경우, 다른 해에 비하여 상대적으로 넓은 면적의 산불 피해를 입은 2021년에는 송이 수매량이 전무하였으나, 다음 해에는 다소 회복되는 모습을 보였다[Figure 3(a); r=-0.41, p=0.24]. 영동 지역인 삼척의 경우에도 765 ha가 소실된 대형 산불이 발생한 2017년에는 수매량이 극히 적었지만, 다음 해에는 수매량이 급증하고 이후 5년간 다소 낮아진 수매량을 유지하였다[Figure 3(e); r= -0.29, p=0.42]. 즉, 영동, 영서 지역을 막론하고 산불이 그 해의 송이 수매량에는 분명히 영향을 미치지만, 다음 해 이후에는 그 영향이 뚜렷하게 나타나지 않음을 확인할 수 있었다.
한편, 고성의 경우에는 2019년, 1,268ha에 달하는 대형 산불을 겪었지만 당년과 다음 해에도 송이 수매량이 많은 모습을 보였다[Figure 2(c); r=0.54, p=0.11]. 그리고, 인제와 양양의 경우에는 홍천에 비해서는 산불 피해 면적이 컸지만 송이 수매량은 전년도와 비교할 때 큰 차이가 없는 것으로 나타났다[Figure 2(b), (d); 인제 r=0.33, p=0.55, 양양 r=0.30, p=0.41].
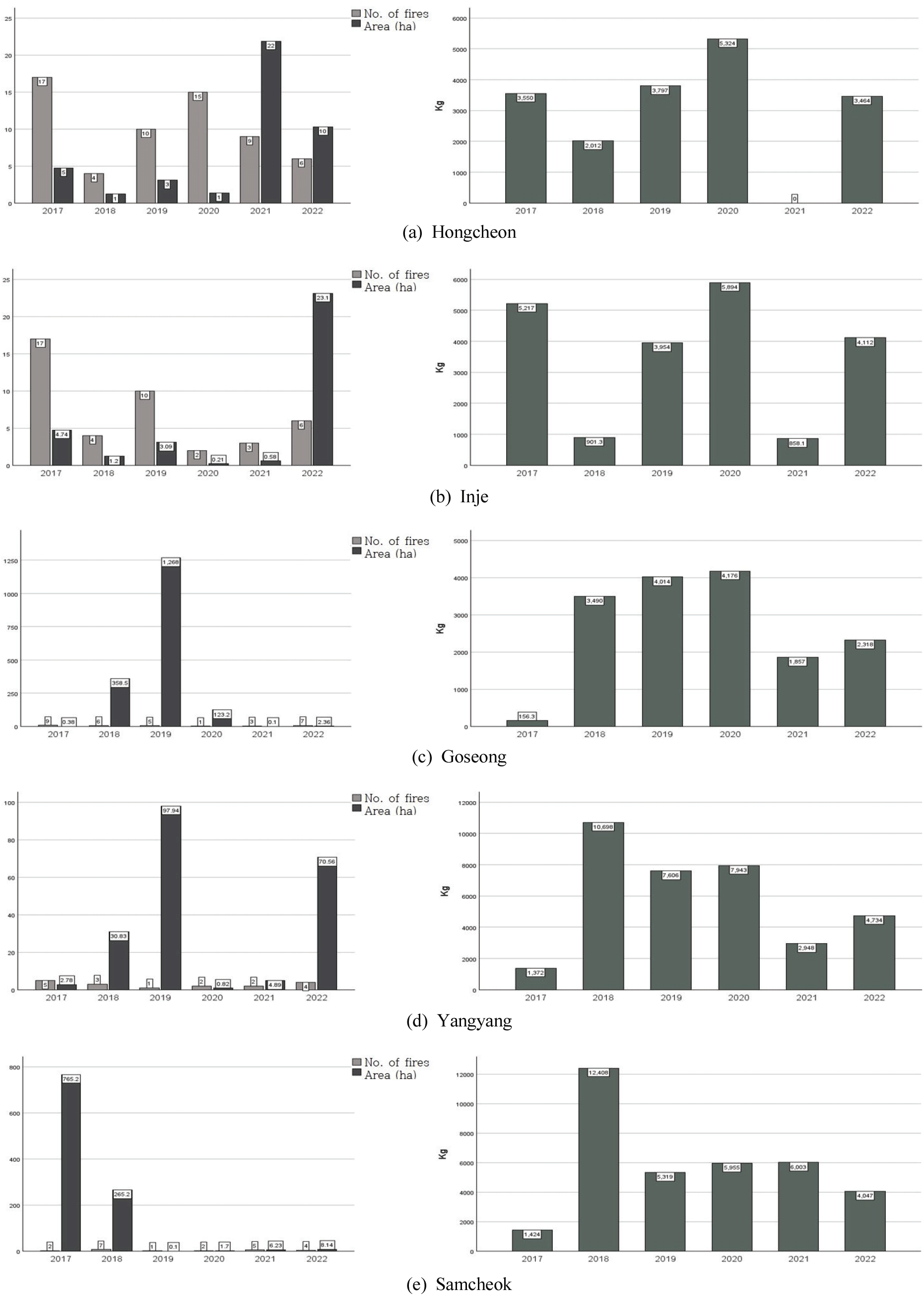
강원도 5개 시·군의 자료를 살펴보면, 산불이 송이 생산량 감소의 직접적 감소 요인이라고 평가하기에는 다소 무리가 따른다. 산불 피해지의 많은 부분이 소나무림이며, 이 중 송이 생산림도 많이 포함되어 있을 수 있다. 하지만, 산불이 송이 생산성을 낮추는 가장 중요한 원인이라고 단정하기는 곤란하다는 의미이다.
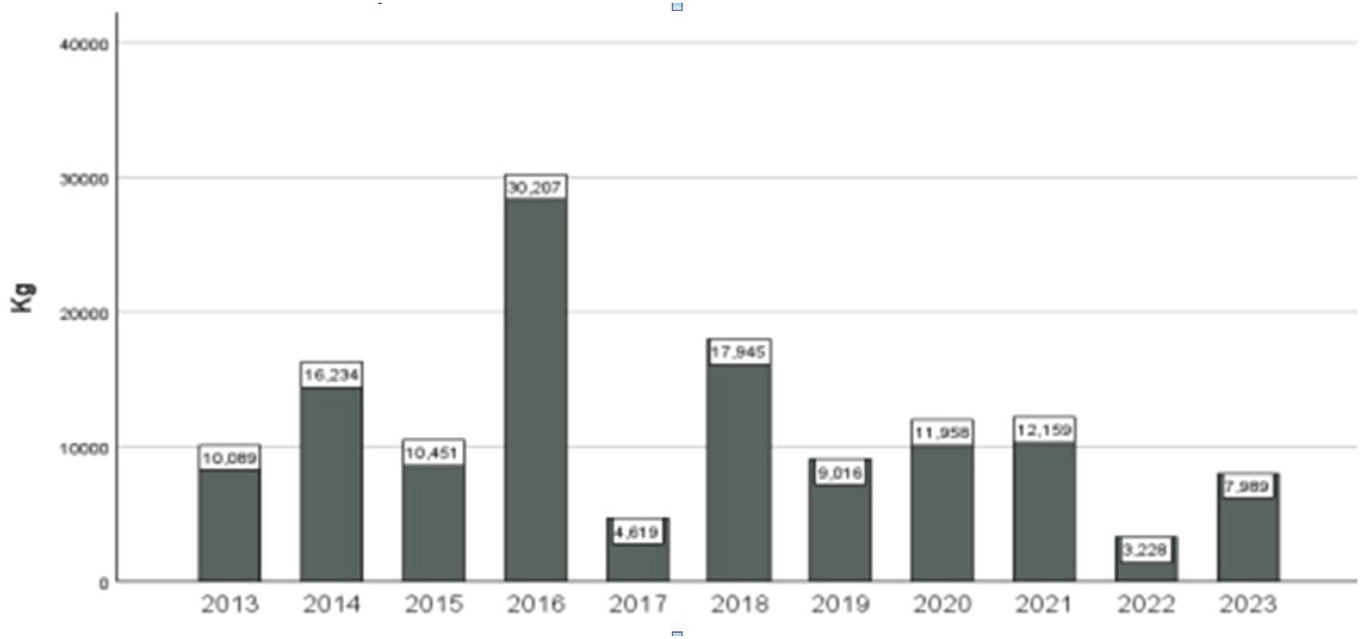
강원도의 전반적인 경향을 보다 심층적으로 분석하기 위하여 최근 대형 산불을 겪은 경상북도 울진을 대상으로 송이 생산량 변화와 관련 인자를 검토하였다. 산림조합 중앙회에서 제공하는 송이 수매 기록(2013-2023)에 따르면, 울진군 산림조합의 송이 공판량은 2016년을 정점으로 다소 감소하는 모습을 보였다(Figure 3; r=-0.43, p=0.22). 산불 피해와의 연관성을 살펴보았을 때, 2022년 대형 산불이 발생한 해의 송이 공판량은 상대적으로 적었지만, 2023년에는 다소 회복된 모습을 보였다. 즉, 강원도의 사례와 마찬가지로 산불이 송이 생산량에 미치는 영향력을 쉽게 판단할 수 없었다. 특히, 산불 피해지가 송이를 많이 생산하던 읍·면에 집중되어 있었다는 의견이 있지만, 피해 보상 단계에서도 피해 산림 소재지가 아닌 피해자의 주거지를 기준으로 자료가 정리되어 정확한 판단이 어려웠다. 기존의 다른 지역 산불에서도 산불 피해지의 ‘실제 송이산 여부’에 대한 논란이 있었음을 감안할 때 산불로 인해 송이 생산량에 미치는 영향을 수치적으로 파악하는 것이 쉽지 않음을 시사한다.
이에 따라 울진군 산림의 임상, 임령 등 산림 정보를 분석하였다. 임상별 산림면적과 임목축적의 변화를 분석하였는데, 임령(영급)별 산림면적과 임목축적의 변화를 파악하였다. 최근 5년의 통계자료를 비교하면, 2015년에 비하여 2020년에는 침엽수림과 혼효림의 면적은 감소한 반면, 활엽수림의 산림면적과 임목축적은 2배 수준으로 증가했다(Table 2). 즉, 송이를 생산할 수 있는 숲(소나무림 또는 소나무 위주 혼효림)이 많이 감소하였음을 시사한다.
세부적으로 숲의 영급 분포변화를 살펴보면, Ⅳ영급 이하의 면적은 줄어들고 Ⅴ영급 이상의 면적이 늘어났다(Table 3). Ogawa(1981)를 비롯한 많은 학자들이 언급한 우리나라 송이 생산성이 최성기가 Ⅳ~Ⅴ영급이라는 것을 감안하면, 경상북도 울진군의 산림은 송이 생산성이 떨어지는 숲으로 변화되고 있음을 시사한다. 특히, 평균 임목축적과 더불어 Ⅴ영급 이상의 임목축적이 많이 증가하고 있어서 숲이 계속 성숙된 모습으로 변화되고 있음을 나타내었다.
모든 소나무림에서 송이가 생산되지는 않는다는 것은 송이 생산량이 소나무림의 존재 여부로만 결정되는 것은 아님을 말한다. 소나무와 송이의 균근 형성을 통한 공생관계 유지가 송이 생산을 위한 필수조건인데, 균근 관계의 유지는 토양이 척박한 조건에서 더 유리하다고 할 수 있다. 왜냐하면, 토양이 비옥해지면 공생기주인 소나무 입장에서는 송이 균근의 도움이 없이도 생활이 가능한 반면, 공생관계를 유지하며 광합성 산물의 20% 수준을 균에게 나눠 주는 것(An et al., 2020)은 비용 효율적인 생활방식이 아니라고 판단할 수 있기 때문이다. 이에 따라 성숙한 숲에서는 공생관계가 약화되는 경향이 나타나는데, 울진을 비롯한 우리나라의 숲은 성숙하면서 균근성 버섯인 송이가 공생관계를 유지하며 생존하기에 부적합한 환경으로 변화되고 있음을 시사한다.
Kim et al.(1975)의 발표를 필두로 우리나라 송이 발생림의 생태적인 특성에 대한 연구가 많이 보고되었다. 하지만, 대부분 논문은 송이가 생산되는 소나무림의 밀도나 수령, 수고, 토양 등에 대한 일반적인 보고로 그 변이가 크고 송이 미발생림과의 비교가 제대로 이루어지지 못하였다. 일본인 학자로 우리나라 학술지에 송이 발생림의 생태적 특성을 소개한 Ogawa(1981)에 따르면, 송이 발생림은 화강암 또는 화강편마암 모재의 척박한 15~30년생 소나무림에서 형성되어 40~50년생 때 최성기에 이르며, 60~80년생이 되면 균환이 소실된다. 하지만, 이는 임분밀도 등 숲의 여건이나 토양 상태에 따라 변화될 수 있다.
21세기의 송이 관련 논문을 살펴보면, Ka et al.(2002)은 65년생 소나무림에서 송이 균환이 매년 평균 11 cm 정도로 성장을 하며 여전히 송이 생산이 왕성히 이루어졌다고 보고하였으며, 관리가 이루어진 숲은 90년생이 된 시점에도 여전히 송이가 생산되고 있다(미발표 자료). Ha et al. (2004)은 경기도에서도 송이가 생산되고 있는데 송이 발생림 소나무는 500~2,700본/ha, 수령은 15~50년생, 수고는 4~11 m로 변이가 컸다. 한편, Chung et al.(2008)은 경북 봉화와 강원도 간성(고성) 지역의 비교를 통해 송이 균사가 토양 수분과 철 이온의 농도가 높은 곳에 주로 분포하는 것으로 추정하였다. 그러나 이는 임목 밀도와 토양 수분함량이 상대적으로 높은 봉화지역이 간성(고성)에 비해 송이 생산량이 많음을 단순 비교하며 추정한 오류라고 할 수 있다. 균근성 버섯인 송이는 배수가 잘되는 토양에서 원활히 생장하는 반면 수분이 많은 곳에서는 부후균의 활성이 더 높아지는 것이 일반적인 사례이다.
송이 균환의 활력은 임분밀도 등 숲의 여건이나 토양 상태에 따라 변화한다. Park et al.(1997)은 식생 정리나 지피물 관리를 통해 송이 생산을 관리할 수 있는데, 이러한 처리는 단기적인 효과를 낳을 뿐이므로 지속적인 관리를 통해서만 송이 생산성을 유지할 수 있다고 하였다. 또한, 지나치게 척박한 토양조건에서는 낙엽을 제거하는 것이 역효과를 낼 수도 있으므로 토양 상황을 고려한 관리가 필요함을 지적하였다.
일반적으로 균근균과 부후균은 생태적 영역에서 경쟁관계를 지닌다. Gadgil and Gadgil(1971)은 라디아타소나무림에서 살아있는 뿌리, 특히 균근을 형성하고 있어서 낙엽 분해균과 경쟁 관계를 띠는 소나무 뿌리는 낙엽의 분해를 저해하여 거친 부식층(mor type)을 형성하며 잎의 분해가 제대로 이루어지지 않도록 하는 효과가 있다고 평가하였다. 즉, 낙엽 분해가 잘 이루어지는 곳에서는 균근의 활성이 낮고, 잘 분해되지 않는 낙엽이 존재하는 경우에는 그 반대 현상이 나타난다고 하였다. 숲이 성숙하면서 낙엽이 많아지고, 이들의 분해가 활성화되면 송이의 활력은 떨어지게 되는데, 낙엽이 분해가 제대로 이루어지지 않는 곳에서는 송이가 계속 활력을 유지할 수 있음을 시사한다.
Coûteaux et al.(1995)이 지적한 것처럼, 낙엽 분해는 기후인자나 토양미생물과 더불어 낙엽의 질에 따라 달라진다. 특히, 낙엽의 질은 부식질을 형성하는 후기 분해 과정까지 영향을 미치는데, 침엽수와 활엽수 또는 다양한 종류의 낙엽이 섞일수록 분해가 촉진되는 것이 일반적이다(Song et al., 2010; Zhang et al., 2023). 즉, 혼효림 조건에서는 낙엽이 잘 분해되는 반면, 침엽수 위주의 낙엽이 유입될 경우에는 낙엽 분해가 원활하지 않으므로 소나무림으로 유지된다면 송이 생산이 지속될 수 있지만, 혼효림으로 변화되면 그렇지 않게 됨을 시사한다.
소나무림의 적정한 밀도 조절은 낙엽을 통한 유기물과 양분의 유입량을 줄일 수 있다. Kim(2016)은 약 40년생의 우리나라 소나무림에서 BA(basal area) 값이 커질수록 수피나 잎의 낙하량이 많아지면서 유기물(C)과 각종 식물 영양분(N, P, K, Ca, Mg) 유입량이 증가함을 보고하였다. 또한, Baek et al.(2022)은 소나무림의 간벌 후 낙엽 유입량이 현격히 줄어들었음을 보고하였다. Ogawa(1981)가 노령 소나무림에서 송이 균환이 쇠퇴한다고 지적한 것과 노령 임분이라도 관리를 하면 계속 송이 생산이 이루어질 수 있다는 논리를 뒷받침하는 보고라고 할 수 있다.
송이 균근 관계 형성과 별개로 송이버섯 생산량은 그 해의 기상 여건에 크게 좌우된다. Kang et al.(1989)은 강수량과 온도가 송이 생산량에 매우 밀접한 영향을 미치며, 지온 조사를 통해 송이 첫 발생 시기의 예측이 가능하다고 발표한 바 있다. Cho and Lee(1995)도 29개 송이 생산 시·군의 10년 자료 분석을 토대로, 가을철 강수량이 송이 생산량에 가장 큰 영향을 미치는 것으로 파악하였다. 또한, Park et al.(1995)은 송이 발생기인 9월에 관수처리를 통해 가뭄을 극복하며 송이 생산성을 증진하는 방법을 제안하였고, Chung and Kim(2005)도 살수장치를 이용한 송이산 관수가 경제적인 효용성이 있다고 보고한 바 있다.
우리나라에서 송이 인공재배 시도는 국립산림과학원을 중심으로 진행되었는데, Ka et al.(2006; 2018)은 기존 송이 균환 선단부에 소나무 묘목을 심어서 균근이 전진하며 소나무 뿌리를 감싸게 한 후 송이가 감염된 묘목을 이식하는 방식으로 송이 미발생 소나무림에서 송이버섯을 수확할 수 있었다. 하지만, 감염묘(感染苗, mycorrhizae-infected seedling) 이식기법은 기존 균환의 훼손과 이식 절차에서의 비효율성이 존재한다. 이를 극복하기 위해 접종묘(接種苗, mycorrhizae-inoculated seedling) 생산기술이 대안으로 제시되지만, 접종묘 생산과 이식 과정에서도 송이균 유지에 난관이 존재한다. 송이는 부후균에 비하여 생장이 매우 느려서 접종원을 만들기가 쉽지 않고, 접종 후 육성단계에서도 균이 사라지는 경우가 많다.경상북도 산림환경연구원에서는 송이 접종묘 생산을 계속 연구하고 있으며, 최근 일본 나가노현의 신슈대학에서는 송이 포자 발아와 이를 통한 접종묘 생산에 성공하고 균사체를 직경 10 cm 수준까지 키울 수 있었다는 보고(Horimai et al., 2021)도 있었다.
한편, Varrio et al.(2011)은 송이가 우점하고 있는 토양에도 다양한 미생물이 공존하고 있음을 확인하였다. 즉, 송이만 분리·배양하여 다른 균이 오염이 되지 않도록 유의하며 접종묘를 만드는데, 굳이 그렇게 해야만 할 필요가 없음을 시사한다. 우리나라에서도 Oh et al.(2016)은 송이 균환 부근의 미생물상 분석을 통해 다양한 세균 및 균류가 존재하고 있음을 밝혔고, 특히 세균류가 송이 균사나 자실체 생장에 긍정적인 영향을 줄 수 있음을 실험적으로 증명했다(Oh and Lim, 2018; Oh et al., 2018). Kim(2024)은 송이와 긍정적인 상호작용을 하는 두 종류의 균류를 분리하였으며, Choi et al.(2023; 2024)도 송이 균환에서 세균을 분리, 접종하여 200% 내외의 송이 균사체 생장 촉진 효과를 거둔 것으로 보고한 바 있다. An et al.(2020)은 균근 및 근권 토양 내의 마이크로바이옴 관련 연구를 통해 송이와 같은 균근성 버섯의 재배를 위한 연구가 꾸준히 진행될 수 있을 것으로 기대하였다.
송이 생태나 마이크로바이옴 연구에서 유의할 사항은 토양 시료 채취의 정확성이다. 최근에 발표되고 있는 많은 논문은 송이 균이 우세한 곳(Tm dominant)과 그렇지 않은 곳(Tm minor)의 두 부분으로 구분하였다. 즉, 송이 균환이 있는 곳과 균환이 아닌 곳으로 구분한 것이다. 하지만, Hur et al.(1999)이 지적한 것처럼, ①송이 균이 지나간 지 2년 이상 경과된 곳과 ②최근에 자실체가 발생한 곳, 그리고 ③현재 송이 균이 왕성하게 자라고 있는 곳과 ④아직 송이 균사가 들어가지 않은 곳은 토양 이화학성 및 소나무 뿌리분포 등에서 큰 차이를 나타낸다. 활성 균환부(③)에서는 소나무 세근의 비율이 평균 60%를 넘으며 타 부위에 비하여 2배를 나타내었다고 하였는데, 근계 동태에 따라 근권의 마이크로바이옴은 큰 차이를 나타낼 수 있다. 즉, 활성 송이 균환부와 송이 균이 지나간 곳, 송이가 아직 도달하지 않은 곳 등의 토양 여건은 큰 차이가 있음에 유의하여야 한다.
Ogawa(1975)는 일본의 소나무림 송이 균환 생장을 관찰한 결과 연간 5 cm~23 cm의 변이가 있음을 보고하였다. Ka et al.(2002)은 강원도 홍천에서 4년간 송이 균환 선단(先端) 및 자실체 발생 위치 측정을 통해 65년생 송이 생산림에서의 균환 생장량은 4 cm~23 cm로 변이폭이 컸으며 평균 11.3 cm라고 보고하였다. 물론, 균환의 연령이나 주변 여건 및 기후인자가 영향을 줄 수 있으며, 균환 쇠퇴기에 이르면 그 생장량이 줄어드는 것으로 추측하였다. 아무튼, 연평균 10 cm 내외의 균환 선단부 이동을 고려하며 토양 시료를 구분하여 채취, 분석하는 것이 바람직하다.
결 론
대한민국 산림산업에서 송이가 차지하는 비중이 줄어들고 있다. 송이 생산량이 감소하고 있기 때문인데, 이는 산불 등에 의한 소나무림의 소실이 주원인이 아니라 소나무림의 쇠퇴가 더 중요한 원인이라고 할 수 있다. 분해가 쉽게 되지 않는 소나무 낙엽 위주의 임상(林床, forest floor)을 유지하는 것이 바람직한 모습이라고 평가되는데, 이를 위해서는 임상(林相, forest type)의 관리가 필요하다. 기후변화와 자연적인 천이로 소나무림이 혼효림으로 변화될 수밖에 없다고 할 수 있지만, 숲의 다양한 기능을 고려하며 태백산맥과 소백산맥 주변이라도 적극적인 임분 관리를 통해 송이 생산림으로 유지하는 것이 바람직하다.
최근에는 송이 재배를 위한 다양한 시도가 이루어지고 있으며, 이를 위한 송이 균환 생태계 연구 및 마이크로바이옴 연구도 활발히 진행되고 있다. 송이 균사의 생장에 긍정적인 영향을 미치는 세균이나 곰팡이의 분리, 배양도 성공한 상태로써 조만간 송이 재배가 이루어질 수 있으리라 기대하게 한다. 하지만, 정밀한 연구를 위해서는 송이 균환에 대한 명확한 이해와 정확한 시료 채취가 필요하다. 송이 균환이 지나간 곳과 활성부위, 그리고 송이의 전진 예정지를 잘 파악하고, 구분하여 각 부분의 특성을 제대로 평가하며 연구를 진행할 수 있어야 한다.