서 론
난대 상록활엽수인 동백나무(Camellia japonica)는 우리나라를 포함한 동아시아의 온대 및 아열대 지역에 자생하고 있다(Kang et al., 2020). 동백나무는 1월부터 3월까지 개화하며, 9월부터 10월 사이에 열매가 성숙하고, 각 열매에는 3~9개의 종자가 포함된다. 전통적으로 동백나무는 생화, 조경수, 정원수, 차 원료 등 다양한 원료로 활용되고 있다(Vela et al., 2013). 특히 동백나무의 종자는 화장품 원료, 식용유 생산, 약용 소재 등 여러 산업 분야에서 활용되고 있는 중요한 경제적 자원이다(Pereira et al., 2022).
동백나무 종자는 불포화 지방산이 풍부하여 ‘동양의 올리브유’로 불릴 만큼 그 가치를 인정받고 있으며, 최근에는 기능성 화장품과 약용 소재로서의 수요가 급증하고 있다(Ji et al., 2022; Pereira et al., 2022). 동백나무 종자의 활용 가치는 최근 연구들을 통해 더욱 주목받고 있다. Kim et al.(2020)은 동백나무 종자 초임계 추출물이 항노화, 항산화, 항염 효과를 나타낸다고 보고하였으며, 이는 화장품 산업에서 요구되는 천연 기능성 소재로서의 가능성을 보여준다. 또한 동백나무 종자는 흑색종 세포의 전이 억제, 항염증, 항바이러스 등 다양한 생리활성 효과를 나타내는 것으로 밝혀졌으며, 바이오연료로서의 활용 가능성도 제기되고 있다(Chung, 2010; José et al., 2016). 추가적으로, 종자유 추출 후 부산물로 발생하는 압착박(seed cake)에서도 폴리페놀, 사포닌, 단백질, 다당류 등 다양한 생리활성 물질이 확인되었다(Li et al., 2015). 이는 기존에 폐기물로 여겨지던 동백나무 종자 부산물이 고부가가치 자원으로 활용될 수 있음을 보여준다.
그러나 동백나무 종자에 대한 산업적 수요가 증가하고 있음에도 불구하고, 안정적이고 지속 가능한 공급은 여전히 해결해야 할 과제로 남아 있다(Park and Choi, 2019). 이러한 수급 불균형을 해소하기 위해 새로운 재배지를 조성하는 것도 하나의 방안이 될 수 있으나, 기존 임분에서 종자 생산성을 향상시키는 접근이 더 효율적일 수 있다. 기존 임분은 이미 성숙기에 도달한 개체들로 구성되어 있어, 종자 생산에 필요한 생리적, 생태적 잠재력이 높으며, 추가적인 토지 확보 없이도 생산량 증대가 가능하다(Viglas et al., 2013). 이에 따라 기존 동백나무 임분에서 종자 생산성을 효과적으로 증진시킬 수 있는 방법을 모색할 필요가 있다.
임분밀도 조절은 종자 생산량을 증대시킬 수 있는 효과적인 임업적 관리 방법 중 하나이다. 낮은 임분밀도는 개체목이 이용할 수 있는 빛, 수분, 영양분을 증가시켜 나무의 활력과 생장을 향상시킬 수 있다(Mäkinen and Isomӓki, 2004; Inagaki et al., 2008). 임분밀도 조절이 종자 생산에 미치는 긍정적인 효과는 여러 선행 연구에서도 보고된 바 있다. 40년생 잣나무 임분을 대상으로 한 연구에서는 낮은 임분밀도에서 나무의 생육 상태가 개선되고, 구과 및 종자 생산량이 증가한 것으로 나타났다(Nguyen et al., 2019). 또한, 침엽수종을 대상으로 한 선행 연구에서도 임분밀도 감소가 나무의 생장과 종자 생산에 긍정적인 영향을 미친다는 결과가 확인되었다(Otto et al., 2012; Ayari and Khouja, 2014). 그러나 임분밀도 조절에는 신중한 접근이 필요하다. 과도한 임분밀도 조절은 생물학적⋅비생물학적 피해를 유발할 수 있으며, 임분 내 온도 상승으로 인한 증산율 증가는 나무의 생산성과 수분 가용성을 저하시킬 수 있다(Niccoli et al., 2020). 이는 임분의 건전성과 생산성을 모두 고려한 적정 임분밀도 설정의 중요성을 시사한다. 현재까지 다양한 수종을 대상으로 임분밀도 연구가 진행되어 왔으나, 동백나무에 대한 연구는 부족한 상황이다. 특히 동백나무 종자 생산과 관련하여 임분밀도의 영향을 규명한 연구는 전무한 실정이다.
따라서 본 연구는 동백나무림에서 임분밀도가 종자 생산량 및 종자의 형태 특성에 미치는 영향을 분석하기 위해 수행되었다. 이를 위해 4년간 수집한 자료를 바탕으로 임분밀도에 따른 종자 생산량과 형태 특성 변화를 분석하였다. 또한, 나무의 생장 특성과 기후 요인이 종자 생산량에 미치는 상호작용을 분석하여 종자 생산에 영향을 미치는 복합적인 요인을 파악하고자 하였다. 이를 통해 동백나무림의 지속 가능한 관리 및 종자 생산성 향상을 위한 기초 자료를 제공하고, 종자의 안정적인 공급 체계 구축에 기여하고자 한다.
재료 및 방법
본 연구는 전라남도 완도수목원 내 산1-74번지에 위치한 동백나무 군락지(34°20'54"N, 126°39'54"E)에서 수행하였다(Figure 1). 연구대상지는 남서사면에 위치하며, 평균 해발고도는 134 m, 경사는 18°~20°이다. 임령은 Ⅲ-Ⅳ 영급이며, 산림토양은 갈색약건산림토양형(B2)이 주로 분포한다. 최근 10년간(2015-2024) 완도군의 연평균 기온은 15.2°C, 연 강수량은 1,488.7 mm로 온난 습윤 기후를 나타낸다(KMA, 2025).

2019년 8월, 임분 밀도에 따른 종자 생산량 및 형태 특성을 조사하기 위해 처리구를 조성하였다. 임분밀도 처리는 대조구(non-thinning, Control)와 약도구(light thinning, LT), 중도구(moderate thinning), 강도구(heavy thinning, HT)로 설정하였다. 열악목과 피압목을 우선적으로 제거하였으며, 잔존목 간의 간격을 고려하여 간벌 작업을 수행하였다. 처리구 조성 후의 임분밀도는 대조구(2,500본 ha−1), LT(1,750본 ha−1), MT(1,250본 ha−1), HT(750본 ha−1)이다. 각 처리구에는 20 m × 10 m 크기의 조사구를 3개씩 배치하였으며, 각 조사구의 상, 중, 하부 위치에 4 m2(2 m × 2 m) 크기의 정방형 종자 트랩을 각각 1개씩 설치하여, 총 36개의 트랩을 설치하였다.
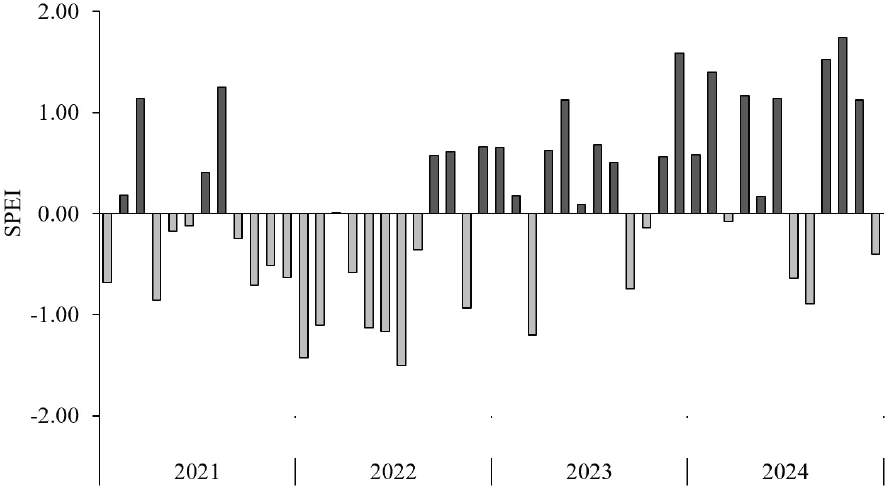
연구 기간 동안 완도 지역의 가뭄 상태를 평가하기 위해 1개월 단위의 표준강수 증발산지수(SPEI)를 산출하였다. SPEI는 강수량과 잠재증발산량의 차이를 바탕으로 가뭄 정도를 평가하며, 기온의 영향까지 고려하기 때문에 기후 조건을 보다 정밀하게 파악할 수 있다(Tirivarombo et al., 2018). 2022년 완도군의 연강수량은 최근 10년 평균의 약 47% 수준인 704.4 mm에 불과하였으며, SPEI 분석 결과, 3월을 제외한 1월부터 8월까지는 경미한 가뭄에서 보통 가뭄에 해당하는 건조한 상태가 지속되었다(Figure 2).
연구대상지의 하층식생은 장딸기(Rubus hirsutus), 수까치깨(Corchoropsis tomentosa), 파리풀(Phryma leptostachya), 송악(Hedera japonica), 동백나무(Camellia japonica), 산기장(Phaenosperma globosa), 마삭줄(Trachelospermum asiaticum), 주름조개풀(Oplismenus undulatifolius)이 우점하였다. 연구대상지의 토양 특성을 파악하기 위해 토양 분석을 실시하였다. 유기물층을 제거한 후, 토심 15 cm까지 고르게 토양 시료를 채취하였다. 채취한 시료는 실험실로 운반하여 풍건한 뒤, 2 mm 체로 걸러진 부분을 이용하여 이화학적 특성을 분석하였다. 입도 분석 결과, 토성은 미사질양토로 분류되었으며, 모래 34.6%, 미사 50.3%, 점토 15.1%로 구성되었다. 토양의 pH는 6.0, 유기물 함량은 8.64%, 전질소 함량은 0.468%로 나타났다. 유효인산 농도는 19.2 mg kg−1이었으며, 양이온치환용량 (CEC)은 15.40 cmolc kg−1로 분석되었다. 치환성 양이온의 농도는 K+; (0.64 cmolc kg−1), Na+; (0.13 cmolc kg−1), Ca2+ (7.47 cmolc kg−1), Mg2+ (2.03 cmolc kg−1)로 나타났으며, 전기전도도 (EC)는 0.36 dS m−1, NaCl 함량은 0.002%로 측정되었다.
2021년 12월과 2024년 12월에 각각 흉고직경과 수고를 조사하였으며, 수관 변수는 2024년에만 측정하였다. 흉고직경(diameter at breast height, DBH)은 직경테이프(Shinill Science, South Korea)를 이용하여 지면으로부터 1.2 m 높이에서 측정하였다. 흉고단면적(basal area, BA)은 조사구 내 모든 수목의 단면적을 합산하여 계산하였으며, 이를 ha당 m2로 환산하였다. 수고(height, H)와 지하고는 수고 측정기(Vertex Laser Geo 2, Haglöf Sweden, Sweden)를 이용하여 측정하였으며, 수고와 지하고의 차이를 수관 길이(crown height, CH)로 정의하였다. 수관 직경(crown width, CW)은 직경 테이프를 이용하여 수관의 장축과 이에 직각으로 교차하는 단축의 직경을 측정한 후, 두 값의 평균으로 계산하였다. 수관 부피(crown volume, CV)는 동백나무 수관의 형태가 타원형인 것을 고려하여 Miranda-Fuentes et al.(2015)에서 제시한 식을 사용하였다(식 1).
식 1에서 Ea는 수관 길이의 절반(m), Eb와 Ec는 각각 수관 직경의 장축과 단축의 절반(m)을 의미한다. 수관투영면적(crown projection area, CPA)은 수관 직경의 장축과 단축을 이용하여 타원 면적 공식으로 계산하였다. 흉고직경은 조사구 내의 모든 나무를 대상으로 측정하였으며, 수고와 수관 변수는 트랩 주변의 개체목을 대상으로 조사하였으며, 대조구는 10본, LT와 MT는 8본, HT는 6본을 측정하였다.
종자 생산량 및 종자 형태 특성을 분석하기 위해 2021년부터 2024년까지 매월 첫째 주에 트랩에 낙하한 종자를 수집하였다. 이 중 동백나무 종자가 낙하하는 시기인 9월부터 12월까지의 종자를 분석 대상으로 하였다. 수집한 모든 종자는 미량 정밀저울(HS-1100A, HANSUNG, Korea)을 사용하여 소수점 둘째 자리까지 측정하였으며, 이를 ha로 환산하여 kg 단위로 표현하였다. ha당 종자 생산량은 각 처리구의 ha당 본수로 나누어 개체목당 생산량을 추정하였다. ha당 생산량은 임분 단위에서의 종자 생산 수준을 파악하여 산림 경영 및 수확 전략 수립에 활용할 수 있으며, 개체목당 생산량은 임분밀도 변화에 따른 개별 나무의 생산성 변화를 분석하는 데 유용하다.
종자 형태 특성 분석을 위해 수집된 종자는 종피를 제거하고 건전, 충해, 부후 종자로 분류하였다. 이후 각 조사구 내 3개 트랩에서 건전 종자를 트랩당 20개씩 임의로 선정하여 측정하였다. 단, 조사구의 총 건전 종자가 60개 미만일 경우, 확보된 모든 건전 종자를 분석에 포함시켰다. 선정된 종자에 대해서는 디지털 캘리퍼(CD-15CP, Mitutoyo, Japan)를 이용하여 길이, 너비를 각각 측정하였으며, 미량 정밀저울을 사용하여 무게를 함께 측정하였다. 측정된 값들은 처리구별로 평균을 산출하여 비교하였다.
처리구에 따른 나무의 생장 특성, 종자 생산량 및 형태 특성의 차이를 평가하기 위해 one-way ANOVA를 실시하였다. 처리구 간 유의한 차이가 나타난 경우, 5% 유의 수준에서 Tukey’s range test를 수행하여 각 처리 간 평균의 차이를 비교하였다. 임분밀도, 나무 생장 변수 및 종자 생산량 간의 상관성을 분석하기 위해 Pearson 상관분석을 수행하였다. SPEI는 R의 SPEI 패키지(ver 1.8-1)를 사용하여 분석하였다(Begueria and Vicente-Serrano, 2023). 모든 통계 분석은 R (ver. 4.2.2)을 사용하여 수행하였다.
결 과
임분밀도 처리 2년 후 2021년도의 평균 흉고직경은 대조구(14.2 ± 0.68 cm), LT(14.0 ± 1.20 cm), MT(15.3 ± 1.63 cm), HT(14.3 ± 3.11 cm)였다(data not shown). 2024년도는 대조구(14.8 ± 0.90 cm), LT(14.7 ± 1.36 cm), MT(16.2 ± 1.67 cm), HT(15.2 ± 3.30 cm)로 나타났다. 상대 흉고직경 생장률은 대조구보다 처리구에서 더 높은 값을 보였다(Table 1). 특히, HT(6.24%)에서는 대조구(3.83%)와 유의미한 차이를 보이며 가장 높은 상대 생장률을 나타내었다(P<0.05). 2021년도 처리구별 평균 수고는 대조구(6.99 ± 0.76 m), LT(6.51 ± 0.19 m), MT(6.75 ± 0.23 m), HT(6.69 ± 0.63 m)였다(data not shown). 2024년도는 대조구(7.16 ± 0.80 m), LT(6.67 ± 0.13 m), MT(6.93 ± 0.32 m), HT(6.77 ± 0.61 m)로 나타났다. 상대 수고 생장률은 대조구(2.33%), LT(2.44%), MT(2.67%), HT(1.30%)로, 대조구와 모든 처리구 간에 유의적인 차이는 없었다(Table 1). 2021년도 처리구별 흉고단면적은 대조구(34.64 ± 8.44 m2 ha−1), LT(21.69 ± 2.16 m2 ha−1), MT(20.76 ± 1.31 m2 ha−1), HT(17.73 ± 4.23 m2 ha−1)였고, 2024년도는 대조구(37.12 ± 8.51 m2 ha−1), LT(24.17 ± 2.53 m2 ha−1), MT(23.63 ± 2.12 m2 ha−1), HT(20.09 ± 4.77 m2 ha−1)였다(data not shown). 4년간의 흉고단면적 생장률은 MT(13.69%)에서 가장 높았으며, 그 다음으로 HT(13.37%), LT(11.42%), 대조구(7.39%) 순으로 나타났다. 처리구는 대조구보다 높은 흉고단면적 생장률을 보였으며, 특히 MT와 HT는 대조구에 비해 유의미하게 높았다(P<0.05).
DBH = Diameter at breast height; H = Tree height; BA = Basal area; CH = Crown height; CW = Crown width; CV = Crown volume; CPA = Crown projection area. Values are presented as mean ± standard error. Different letters within columns indicate significant differences among treatments (P<0.05).
2024년도의 처리구별 수관 특성을 분석한 결과, 수관 길이는 LT(5.27 m)에서 가장 길었고, 대조구(5.11 m), MT(5.08 m), HT(4.89 m) 순으로 나타났다(Table 1). 그러나 처리구 간 통계적으로 유의미한 차이는 없었다(P>0.05). 한편, 수관 직경, 수관 부피, 그리고 수관투영면적은 모두 HT에서 가장 큰 값을 보였다. HT의 평균 수관 직경은 4.41 m로, 대조구(3.53 m)보다 컸으며, 평균 수관 부피는 52.53 m3, 수관투영면적은 15.58 m2로 각각 대조구(33.42 m3, 9.92 m2)에 비해 높은 값을 보였다. 처리 강도가 증가할수록 수관 크기가 증가하는 경향이 관찰되었으나, 모든 변수에서 처리구 간 통계적으로 유의한 차이는 나타나지 않았다(P>0.05).
2021년부터 2024년까지의 처리구별 연평균 종자 생산량을 비교한 결과, 대조구(116.8 kg ha−1 year−1), LT(156.7 kg ha−1 year−1), MT(108.1 kg ha−1 year−1), HT(145.9 kg ha−1 year−1)이었다(data not shown). 또한, 각 연도에서 처리구별 종자 생산량의 평균값을 비교한 결과, 2023년(270.6 kg ha−1)에 가장 많은 종자가 생산되었으며, 2024년(40.8 kg ha−1)에 가장 적은 종자가 생산되었다. 2021년(106.7 kg ha−1)과 2022년(109.4 kg ha−1)의 종자 생산량은 비교적 유사한 수준을 보였다(data not shown).
개체목 당 종자 생산량의 경우, 모든 연도에서 대조구 대비 처리구에서 종자 생산량이 높았다[Figure 3(a)]. 그 중 2024년을 제외한 모든 연도의 HT에서 종자 생산량이 대조구와 유의적인 차이를 보이며 가장 많은 종자 생산량을 보였다(P<0.05). 반대로 모든 연도에서 대조구가 가장 낮은 종자 생산량을 나타내었다. 4년간의 축적량에서 개체목 당 종자 생산량은 HT(0.78 kg tree−1), LT(0.36 kg tree−1), MT(0.35 kg tree−1), 대조구(0.19 kg tree−1) 순으로 종자를 생산하였으며, HT는 대조구 대비 약 4배 높은 수준이었다.
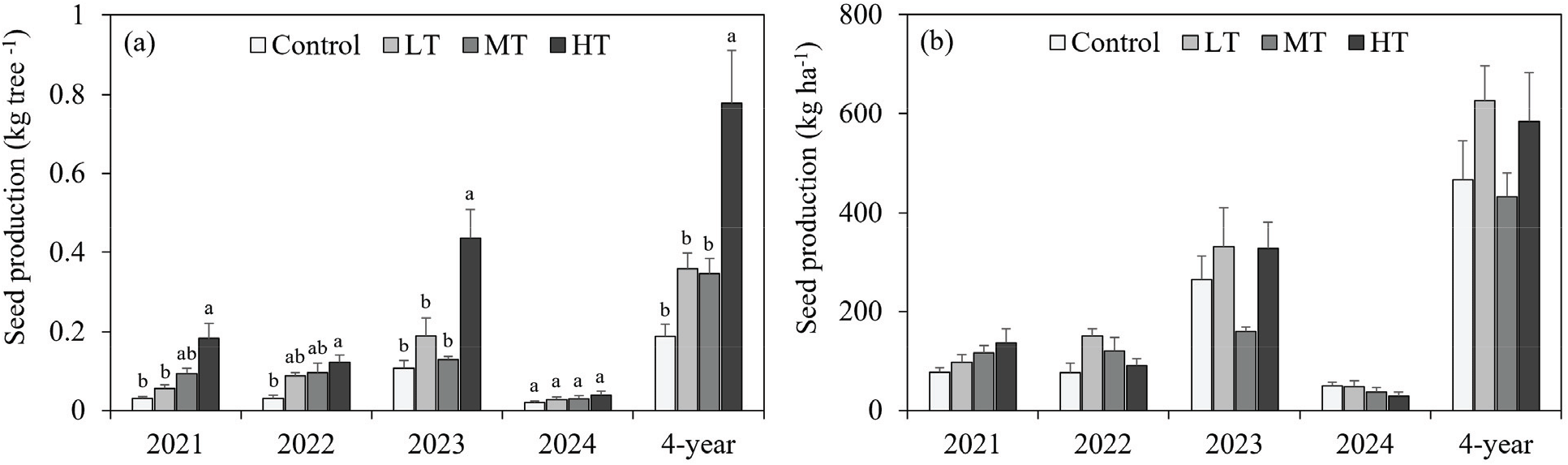
ha당 연간 종자 생산량은 연도와 처리구에 따라 변동을 보였다[Figure 3(b)]. 2021년에는 HT(137.3 kg ha−1)에서 가장 많은 종자가 생산되었으며, MT(115.9 kg ha−1), LT(96.8 kg ha−1), 대조구(76.7 kg ha−1)의 순으로 나타났다. 2022년에는 LT(151.6 kg ha−1)에서 최대 생산량을 기록했으며, MT(119.19 kg ha−1), HT(90.66 kg ha−1), 대조구(76.0 kg ha−1) 순이었다. 2023년에는 모든 처리구에서 종자 생산량이 급격히 증가하여 LT(330.7 kg ha−1), HT(326.8 kg ha−1), 대조구(264.7 kg ha−1), MT(160.3 kg ha−1)로 확인되었다. 그러나 2024년에는 전반적으로 종자 생산량이 감소하여 대조구(49.56 kg ha−1), LT(47.76 kg ha−1), MT(36.86 kg ha−1), HT(28.87 kg ha−1)로 관찰되었다. 4년 누적 종자 생산량은 LT(626.9 kg ha−1)에서 가장 높았으며, 이어서 HT(583.6 kg ha−1), 대조구(470.0 kg ha−1), MT(432.2 kg ha−1) 순으로 나타났다. LT와 HT는 대조구 대비 각각 34.2%, 24.9% 증가한 반면, MT 처리구는 대조구보다 7.4% 감소하였다. 그러나 모든 연도에서 처리구 간 종자 생산량은 통계적으로 유의하지 않았다(P>0.05).
종자 생산량에 영향을 미치는 요인을 파악하기 위해 임분밀도 및 나무의 생장 특성과 종자 생산량 간의 상관분석을 실시하였다(Table 2). ha당 종자 생산량은 어떤 변수와도 유의적인 상관관계를 보이지 않았으나, 개체목 당 종자 생산량은 임분밀도 및 흉고단면적과 음의 상관관계를 나타내었다(P<0.05). 그러나 다른 나무 생장 및 수관 변수와는 상관관계가 나타나지 않았다. 한편, 임분밀도(SD)는 흉고단면적(BA)과 높은 양의 상관관계(r = 0.782)를 보였으며, 수관직경(r = −0.580) 및 수관 투영 면적(r = −0.581)과는 유의미한 음의 상관관계를 나타냈다. 흉고직경(DBH)은 수관 부피(r = 0.889), 수관 직경(r = 0.783), 그리고 수관 투영 면적(r = 0.807)과 강한 양의 상관관계를 보였다(P<0.05).
SD = Stand density; DBH = Diameter at breast height; H = Tree height; BA = Basal area; CH = Crown height; CW = Crown width; CV = Crown volume; CPA = Crown projection area. * and ** indicate significance at P<0.05 and P<0.001 respectively.
동백나무 종자의 형태적 특성은 임분 밀도와 조사 연도에 따라 다양한 양상을 보였다(Table 3). 종자의 길이는 12.4115.31 mm, 너비는 8.60-11.59 mm 범위로 관찰되었다. 2022년에는 대조구(12.64 mm)와 HT(14.55 mm) 사이에서 종자 길이에 유의한 차이가 나타났으나, 다른 연도에서는 대조구와 처리구 간 종자 길이 및 너비 모두에서 유의한 차이가 발견되지 않았다. 다만, 전반적으로 처리구에서 대조구보다 종자의 길이와 너비가 증가하는 경향이 확인되었다. 종자의 무게는 0.42-0.70 g 범위로 관찰되었다. 2022년을 제외한 모든 연도에서 처리구 간에 유의한 차이는 나타나지 않았으나, 마찬가지로 대조구보다 처리구에서 종자의 무게가 증가하는 경향이 있었다. 특히 2022년에는 대조구(0.42 g)와 HT(0.58 g) 사이에서 통계적으로 유의한 차이가 확인되었다(P<0.05). 전반적으로 낮은 임분 밀도는 동백나무 종자의 형태적 특성에 긍정적인 영향을 미치는 것으로 나타났으며, 특히 HT 처리구에서 가장 큰 종자가 생산되는 경향이 있었다. 다만, 이러한 효과는 조사 연도에 따라 다르게 나타났으며, 특히 2022년에만 그 차이가 뚜렷하게 관찰되었다.
Values are presented as mean ± standard error. Different letters within the same year indicate significant differences among thinning treatments (P<0.05) by one-way ANOVA followed by Tukey’s HSD test.
고 찰
임분밀도 조절 4년 후, 동백나무림에서는 임분밀도 감소가 개체목의 생장에 전반적으로 긍정적인 영향을 미치는 것으로 나타났다. 임분밀도가 낮아질수록 흉고직경 생장률이 증가하고 수관의 수평적 크기도 확장되는 경향을 보였다. 그러나 수고의 상대 생장률은 처리구 간 유의한 차이를 보이지 않았다. 이와 유사한 경향은 너도밤나무(Fagus sylvatica), 물푸레나무(Fraxinus excelsior) 등 6종의 활엽수에서도 관찰되었다(Hawe and Short, 2016). 이들 수종은 임분밀도가 낮을수록 흉고직경이 증가하고 수관이 확장되었으나, 수고는 간벌의 영향을 비교적 적게 받는 것으로 나타났다. 앨리오도라 나무(Cordia alliodora)와 잣나무(Pinus koraiensis)에서도 임분밀도에 따른 수고의 변화는 관찰되지 않았다(Hummel, 2000; Nguyen et al., 2019). 수관 특성의 경우, 수관 길이를 제외한 직경, 부피, 투영면적은 임분밀도가 낮아질수록 증가하는 경향을 보였다. Li et al.(2022)의 연구에서도 높은 임분밀도에서는 개체 간 경쟁으로 인해 수관 확장이 제한되었으나, 간벌 이후 잔존 개체의 수관 발달이 촉진되는 양상이 관찰되어 본 연구 결과와 일치하였다. 따라서 동백나무의 경우, 임분밀도는 흉고직경과 수관 발달에는 영향을 미치지만, 수고 생장에는 유의미한 영향을 미치지 않는 것으로 판단된다.
종자 생산량과 임분 및 나무 생장 특성 변수와의 상관 분석에서, ha당 종자 생산량은 어떤 변수와도 유의한 상관관계를 보이지 않았다. 개체목당 종자 생산량은 임분밀도 및 흉고단면적과 유의한 음의 상관관계가 있었으나(P<0.05), 흉고직경, 수고, 수관 특성과는 상관관계가 나타나지 않았다. 반면 Ayari et al.(2012)은 알레포 소나무(Pinus halpensis) 임분에서 흉고직경, 수관 크기, 임분밀도가 구과 및 종자 생산량과 상관관계를 보였고, 나무의 직경과 수관의 크기가 클수록 구과 및 종자 생산량이 증가한다고 설명하였다. 이와 유사한 결과는 여러 선행연구들에서도 보고된 바 있다(Moreno-Fernández et al., 2013; Calama et al., 2008). 동백나무의 경우, 나무의 크기보다 임분 내 빛의 도달 정도나 토양 수분과 같은 환경적 요인이 종자 생산량에 더 큰 영향을 미쳤을 가능성이 있으며, 이에 대한 추가적인 연구가 필요할 것으로 판단된다.
임분밀도 조절은 동백나무의 종자 생산 능력을 향상시키는 것으로 나타났다. 특히 개체목의 4년간 종자 생산량은 HT에서 대조구에 비해 약 4배 이상 증가하였다. 간벌은 개체목 간 생육 공간을 확장시키고, 빛, 수분, 무기 양분과 같은 자원의 가용성을 높여 종자 생산량을 증가시키는 효과가 있다(Mäkinen and Isomӓki, 2004; Inagaki et al., 2008; Ayari et al., 2012). 20년생 우산소나무(Pinus pinea) 임분에서도 간벌 처리구의 구과 생산량이 대조구에 비해 두 배 이상 증가한 것으로 나타났으며, 다른 선행 연구들에서도 유사한 결과가 보고되었다(Moreno-Fernández et al., 2013; Ayari and Khouja, 2014).
단위 면적(ha)당 종자 생산량은 대조구와 처리구 간에 유의한 차이를 보이지 않았다. HT에서는 개체목당 종자 생산량이 증가하였음에도 불구하고, 간벌로 인한 총 입목 수의 감소로 인해 단위 면적당 총 생산량은 다른 처리구와 유사한 수준을 나타내었다. 이는 낮은 임분 밀도에서 개체목의 생산성 증대 효과가 입목 수 감소로 상쇄되었음을 의미하며, 백송(Pinus strobus)을 대상으로 한 선행 연구와도 유사한 결과이다(Parker et al., 2013). 이러한 점은 적정 간벌 강도 설정 시 개체목 단위 생산성과 단위 면적당 총 생산성 간의 균형을 고려할 필요가 있음을 시사한다.
동백나무는 낮은 임분밀도 조건에서 종자의 크기와 질량이 증가하는 경향을 보였으며, 특히 HT에서 가장 큰 종자 길이와 무게가 관찰되었다. 이러한 경향은 잣나무, 삼나무(Cryptomeria japonica), 편백(Chamaecyparis obtusa) 등 침엽수종을 대상으로 한 선행 연구에서도 보고된 바 있다(Lee et al., 2006; Nguyen et al., 2019). 반면, 붉가시나무(Quercus acuta)를 대상으로 한 연구에서는 간벌 처리구와 대조구 간 종자 길이 및 무게에 유의미한 차이가 발견되지 않았으며, 구주소나무(Pinus sylvestris)의 경우에도 종자의 품질은 연도에 따라 차이는 있었으나 처리구 간 차이는 크지 않은 것으로 보고되었다(Almqvist and Jansson, 2015; Kim and Park, 2021). 이는 동백나무 종자의 크기와 질량이 임분밀도의 영향을 받을 수 있음을 보여주는 한편, 이러한 임분밀도 효과는 수종에 따라 상이하게 나타날 수 있는 것으로 판단된다.
종자의 크기와 질량은 영양분 배분과 밀접한 관련이 있으며, 일반적으로 크고 무거운 종자는 더 많은 영양분을 포함하고 있다(Souza and Fagundes, 2014; Bu et al., 2019). 유칼립투스속(Eucalyptus), 하케아속(Hakea), 뱅크시아속(Banksia) 수종을 대상으로 한 연구에서도, 큰 종자가 더 많은 무기 영양소와 탄소 기반 저장 물질을 포함하고 있음이 보고되었다(Milberg et al., 1998). 본 연구에서는 낮은 임분 밀도에서 동백나무 종자의 크기와 질량이 증가하는 경향이 확인되었지만, 이러한 크기 증가가 실제로 종자 내 기름 및 생리활성 물질 함량의 증가로 이어지는지는 명확하지 않다. 따라서 향후 해당 성분들에 대한 분석을 통해 종자 크기와의 관계를 규명할 필요가 있다.
모든 처리구에서 동백나무의 종자 생산량은 연도별로 큰 변동성을 보였으며, 이는 해거리(masting) 현상과 관련이 있는 것으로 판단된다(Greene and Johnson, 2004; Pearse et al., 2016). 해거리란 식물이 매우 가변적이면서도 동기화된 방식으로 종자를 대량 생산하는 생태적 현상을 의미한다(Vacchiano et al., 2018; Koenig, 2021). 해거리의 주기는 수종에 따라 다르게 나타나며, 기온과 강수량 등 기후 조건에 따라 달라질 수 있다(Bogdziewicz et al., 2020).
SPEI 분석에 따르면, 2022년 완도군은 낮은 강수량으로 경미한 수준에서 보통 수준의 가뭄이 발생하였다. 이러한 기후 조건은 동백나무의 결실 주기에 영향을 미쳐, 일반적으로 격년 결실로 알려진 주기와는 다른 양상을 나타내었다(Uemura, 2011). 해거리를 하는 수종은 기후변화의 영향으로 결실 주기에 변화가 생기기도 한다. 일본의 북부 온대림에 분포하는 물참나무(Quercus mongolica)는 기후변화로 인한 기온 상승에 의해 기존의 3~4년 주기의 매스팅 주기가 약 2년으로 짧아졌다고 보고되었다(Bogdziewicz, 2022). 또한, 유럽의 너도밤나무(Fagus sylvatica)는 일부 지역에서 6년간의 연구 기간 동안 뚜렷한 격년 결실 경향을 보였으나, 다른 지역에서는 극단적인 기상 현상으로 인해 그 주기가 교란되었다는 연구 결과가 있다(Gavranović Markić et al., 2024). 이러한 선행 연구들은 기후 변화로 인한 외부 환경 변화가 수종의 결실 주기에도 영향을 미칠 수 있음을 시사한다.
한편, 2023년에 동백나무 종자가 대량으로 생산된 이후, 2024년에는 종자 생산량이 현저히 감소하는 양상이 나타났다. 해거리 현상을 보이는 수종의 경우, 대량 결실로 인해 내부 자원이 고갈되면서 그 결과로 다음해의 종자 생산이 감소하는 경향이 있다(Sork et al., 1993; Pearse et al., 2016). 12종의 참나무를 대상으로 한 선행 연구에서 한 해의 대량 결실 이후 다음 해에는 종자 생산량이 크게 감소하였으며, 이는 자원 고갈에 따른 결과로 해석되었다(Moreira et al., 2019). 동백나무에서도 이와 유사한 경향이 관찰되었으나, 해거리 현상은 자원 가용성 뿐만 아니라 기후 요인 등 다양한 요소가 복합적으로 작용한 결과이므로 수종에 따라 그 양상이 달라질 수 있다(Smaill et al., 2011). 따라서 동백나무 종자 생산의 연도 간 변동성을 이해하기 위해서는 자원 가용성과 기후 등 다양한 환경 요인을 함께 고려한 분석이 필요하다.
해거리에 따른 임분밀도 효과를 분석한 결과, 대량 결실이 발생한 2023년에는 처리구 간 종자 생산량에 유의미한 차이가 나타났으나, 생산량이 급감한 2024년에는 이러한 차이가 관찰되지 않았다. 이는 결실이 많은 해에는 간벌에 따른 임분 밀도 조절이 개체목 종자 생산에 긍정적인 영향을 미치지만, 결실이 적은 해에는 그 효과가 제한적일 수 있음을 시사한다. 이러한 경향은 더글라스 전나무에 대한 연구에서도 관찰된 바 있다(Moreno-Fernández et al., 2013). 따라서 동백나무는 종자 생산이 저조한 해에는 간벌 처리의 효과가 제한적인 것으로 판단된다.
결 론
본 연구는 동백나무림의 임분밀도가 종자 생산량, 종자의 형태 및 나무 생장에 미치는 영향을 4년간의 자료를 바탕으로 분석하였다. 임분밀도에 따른 개체목당 종자 생산량과 종자의 크기는 낮은 임분밀도에서 높아지는 경향을 보였으나, 단위면적당 종자 생산량은 대조구와 처리구간 유의한 차이가 없었다. 이는 임분밀도 조절 시 개체목의 생산성과 단위면적 당 생산성 간의 균형을 고려해야 함을 시사한다. 본 연구 결과는 동백나무 종자의 산업적 수요 증가에 대응하여 지속 가능한 공급 체계를 구축하는 데 있어 유용한 기초 자료로 활용될 수 있다. 그러나 연구가 한정된 지역에서 수행된 만큼 다양한 지역과 환경 조건에서의 장기적인 연구가 필요하다. 또한, 종자의 질적 특성, 미기상, 토양환경, 생리적 반응 등에 대한 추가적인 연구를 통해 동백나무림의 생산성 향상을 위한 체계적인 관리방안을 제시할 수 있을 것으로 기대된다.