Article
서울시 남산 신갈나무림의 식물 군집 및 개체군 동태(2017년~2023년)
하민선
1, 이정은
1, 최훈기
2, 한상학
3, 박정수
4, 이재연
4, 윤충원
1,*
Plant Populations and Community Dynamics in a Quercus mongolica Forest in Mount Namsan, Seoul, Over 6 Years (2017-2023)
Min-Seon Ha
1, Jeong-Eun Lee
1, Hun-Gi Choi
2, Sang-Hak Han
3, Jeong-Su Park
4, Jae-Yeon Lee
4, Chung-Weon Yun
1,*
1Department of Forest Science, Kongju National University, Yesan 32439, Korea
2Division of Forest Biodiversity Conservation Research, Korea National Arboretum, Pocheon 11186, Korea
3Team of Climate Carbon Research, National Institute of Ecology, Seocheon 33657, Korea
4Team of Climate Ecology Observation, National Institute of Ecology, Seocheon 33657, Korea
© Copyright 2025 Korean Society of Forest Science. This is an Open-Access article distributed under the terms of the
Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits
unrestricted non-commercial use, distribution, and reproduction in any
medium, provided the original work is properly cited.
Received: Jan 19, 2025; Revised: Mar 19, 2025; Accepted: May 27, 2025
Published Online: Jun 30, 2025
요 약
우리나라에서 가장 인구밀도가 높아 인위 및 자연 교란이 지속되는 서울시 남산은 비교적 자연성이 높은 신갈나무림이 넓은 면적을 차지하고 있다는 점에서 생태 가치, 보전 가치 등이 크다. 본 연구는 도시림의 생태적 관리와 유지 방안의 생태학적 근거 자료를 확보하고 이를 활용하고자 서울시 남산의 신갈나무림을 대상으로 6년간(2017년~2023년) 식물 군집 및 개체군의 동태를 구명하고자 하였다. 따라서 국가장기생태연구를 통해 구축된 서울시 남산 신갈나무림 1 ha (100 m×100 m)를 대상으로 밀도, 고사율과 이입률, 흉고단면적, 흉고직경급 분포, 중요치, 수간건강상태 및 임목 쇠퇴율 분석을 실시하였다. 측정 기간동안, 5개체의 진계목과 23개체의 고사목이 발생한 신갈나무의 연고사율과 연이입율은 각각 0.5% 및 2.2%로 계산되었으며, 2019년 발생한 개체 밀도 감소에 따라 흉고단면적은 감소 후 다시 증가하였다. 우점종으로서 중요치가 가장 높은 신갈나무는 20-30 cm의 직경급에서 개체목 밀도가 가장 높은 정규분포를 나타냈고, 꾸준하게 임목쇠퇴율이 증가하고 있었다. 하지만, 작은 직경급에서 개체목 밀도가 높았고, Alive Standing (AS) 유형이 대다수를 차지하여 조사 장소의 우점종이 다른 수종으로 대체될 가능성은 낮은 것으로 판단되었다. 결론적으로, 지속적인 교란에도 불구하고 남산 신갈나무림의 주요 목본 개체군들은 높은 지속가능성을 나타내는 것으로 평가되었다. 앞으로도 도시림 내의 신갈나무림이 잘 유지될 수 있도록 지속적인 모니터링이 요구되는 바이다.
Abstract
Namsan, Seoul, is located at the most densely populated area in Korea and is subject to ongoing anthropogenic and natural disturbances. The large Quercus mongolica forest in Namsan holds significant ecological and conservation value. This study monitored a 1-ha (100 m × 100 m) plot within this forest from 2017-2023 as part of the Korea Long-Term Ecological Research, which aims to provide baseline data for the ecological management and maintenance of urban forests. We analyzed tree density, mortality rate, recruitment rate, breast height area (BHA), diameter distribution, importance values, trunk health status, and tree decline rate. Over the study period, 23 trees died and 5 young individuals were recruited, yielding annual mortality and recruitment rates of 2.2% and 0.5%, respectively. BHA decreased in 2019 due to decrease in density but subsequently recovered. The dominant species, Q. mongolica, was normally distributed, with the highest tree density in the 20-30 cm diameter class. Although the tree decline rate was steadily increasing, the high density of the small diameter class of trees and the prevalence of “alive and standing” trees indicate that Q. mongolica is likely to remain dominant. In conclusion, despite ongoing disturbances, the Q. mongolica forest in Namsan demonstrates a high potential for sustainability. However, continuous monitoring is essential to ensure the long-term maintenance of Q. mongolica forests in urban areas.
Keywords: urban ecosystem; urban forest; importance value; breast height area; dbh analysis
서 론
도시 생태계는 참나무 시들음병과 같은 수목 병해, 이용객들에 의한 인위적 교란과 훼손에 놓였기 때문에 도시림(urban forest) 고유의 생태계 기능을 유지하기 위한 식생 군집의 동태 연구는 필수적으로 이루어져야 한다(Kim et al., 2011; Cho et al., 2011, Seoul, 2023; Kim et al., 2023). 이것은 수십 년 이상 진행되는 장기생태연구(Long-Term Ecological Research, LTER)를 통해 이루어질 수 있으며(Son, 1997; Cho et al., 2011; Cho et al., 2020; NIER, 2013; Lee, 2023), 장기생태연구에서 획득되는 정보는 지구 생태계의 구조와 기능의 현황과 변화를 이해하는 데 기여할 뿐만 아니라, 관측 생태계의 유지 및 효과적인 관리에도 중요한 정보를 제공한다(Cho et al., 2006; Kim et al., 2011).
우리나라의 도시생태계를 대표하는 서울시의 남산은 일제 강점기 이후 도시화 및 산업화 과정에서 발생한 부정적인 인위 영향으로 경관구조의 조각화 및 파편화가 진행되었고(Lee et al., 2006; Seoul, 1992), 2020년 기준 우리나라의 평균 산림 면적 비율은 62.7%이지만, 서울은 전국에서 가장 낮은 25%에 불과하다(Lim et al., 2021). 1990년대 후반, 서울시는 남산 산림생태계 보전의 중요성을 인식하였으며, 그곳의 신갈나무림 일부를 생태경관보전지역으로 지정하였다(Seoul, 2008). 이러한 점에서 남산이 갖는 생태적 가치와 그것의 변화를 기록하는 장기생태연구는 도시림의 생물다양성과 생태 기능 보전 관리에 매우 중요한 관점을 제공할 수 있다(Lee et al., 2006; Lim, 2021; Han et al., 2022).
개체목의 고사와 이입을 통해 진행되는 개체수 통계량(demographic rates)의 변동 과정으로서, 임분 동태의 측정은 그들의 장기 변화 예측의 기초 활동이며, 종조성과 다양성의 변화 양상을 이해하는데 필요한 핵심 정보를 제공한다(Condit, 1998; Park et al., 2024). 다양한 스트레스 요인에 노출되어 있는 남산의 식생 조건을 고려하면, 선구종(pioneer species)과 천이 후기종의 교대 양상을 통해 임분의 스트레스 상태, 그리고 장기적인 변화 방향을 이해할 수 있다. 우점하는 신갈나무와 당단풍나무와 같이 천이 후기 수종들의 개체군 성장 양상은 남산 생태계의 기능 변화에 관한 핵심 요인이다. 또한, 흉고직경 계급별 개체 밀도를 나타내는 직경급 분포는 임분의 구조를 분석할 때 가장 흔히 이용하는 방법으로, 군집 또는 개체군의 유지 가능성을 판단하는 데 중요한 정보를 제공한다(Austin, 1976; Barbour et al., 1987; Lee et al., 2007; Ko et al., 2013). 서울시에서 발생하는 부정적인 인위 교란압이 지속되고 있는 남산의 신갈나무림은 천이후기 식생으로서 상대적으로 안정성이 높다. 따라서, 남산 신갈나무림과 같은 도시 생태계의 낙엽활엽수림의 동태를 이해하는 것은 우리나라 식생 변화 양상에 관한 중요한 보고가 될 것이며, 도시림 보전 관리에 필요한 기초 자료를 제공할 수 있다.
본 연구는 국가장기생태연구(KLTER, Korean Long Term Ecological Research)가 실시되고 있는 남산 신갈나무림에서 최근 6년간(2017-2023)의 식물 군집과 개체군의 동태를 분석하여 그들의 변화 양상을 이해하고, 이를 바탕으로 도시림의 생태 관리에 필요한 근거 자료를 제공하고자 수행되었다.
재료 및 방법
1. 조사지 개황
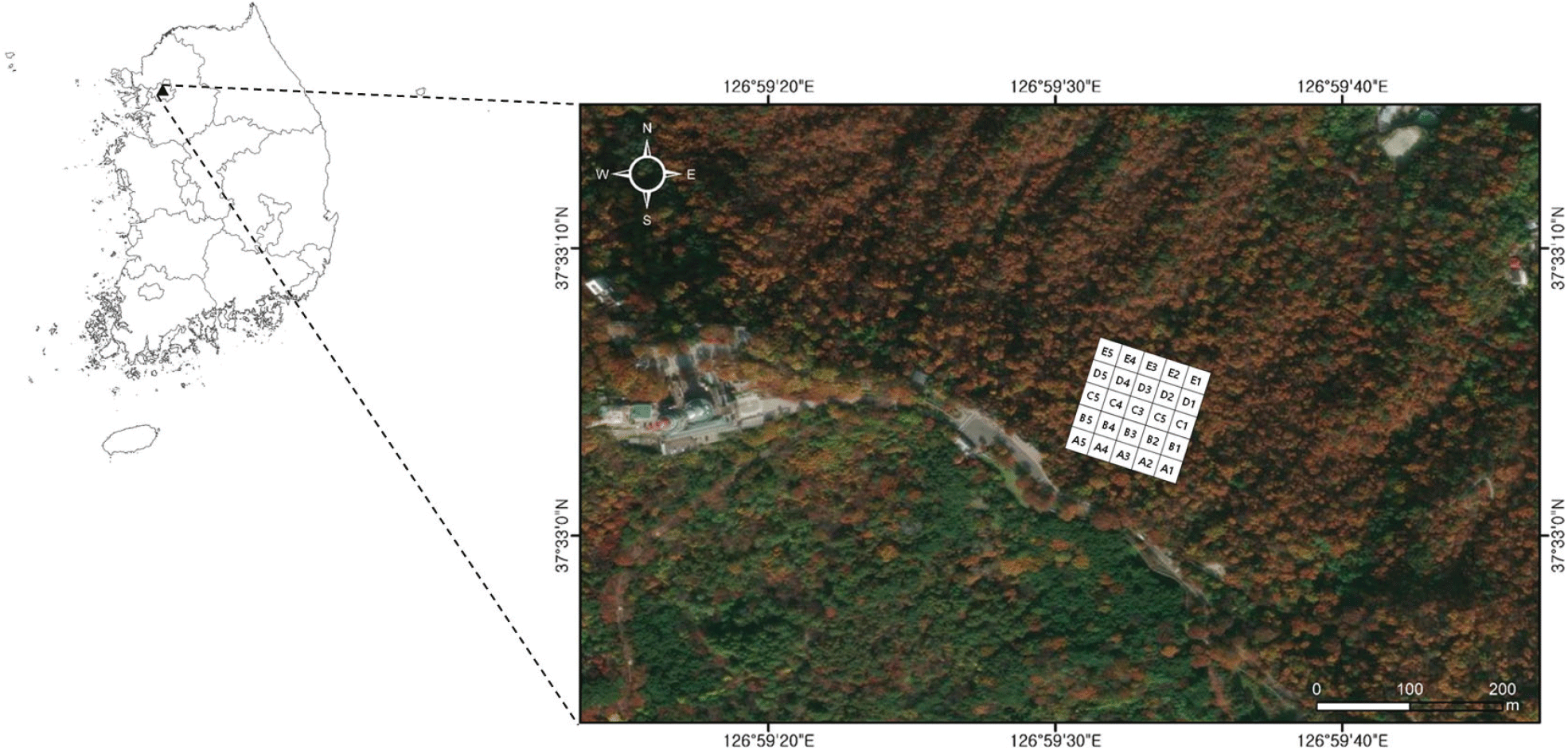
남산은 서울시의 중심부로서 북위 37° 32′~33′ 사이, 동경 126° 58′~127° 00′ 사이에 위치한다(Figure 1). 한강의 북쪽에 위치하므로 백두대간 마루금의 식개산에서 분기하여 경기도 운악산, 도봉산, 남산으로 이어지는 한북정맥에 속한다(Kim et al., 2023; Park, 2004; Son et al., 2016). 산림대는 온대중부(Yim and Kira, 1976), 식물구계는 중부아구(Lee and Yim, 1978), 생태권역은 중부산야권역(Shin and Kim, 1996)에 각각 속한다(Son et al., 2024).
Figure 1.
The location of LTER (Long Term Ecological Research) site of Q. mongolica community in Mt. Namsan, Seoul city.
Download Original Figure
행정구역으로는 서울특별시 중구와 용산구에 걸쳐 위치해 있으며, 중구의 6개 동(회현동, 남산동, 예장동, 필동, 장충동, 신당동)과 용산구의 4개 동(후암동, 이태원동, 용산동, 한남동)을 포함하고 있다(Seoul, 1992).
남산은 지형적으로 동서로 달리는 능선을 경계로 하며, 북사면은 경사가 급하고 남사면은 비교적 경사가 완만한 남고북저의 지형을 이루고 있다. 본 연구의 조사구 경사는 5~15°로 완경사지, 평균 경사도는 7°로 각각 나타났다. 남산 북사면은 주로 화강암이며, 남사면의 모암은 주로 편마암으로 이룬 화강편마암 지대이다. 남산의 토양은 2000년 pH 4.49로 낮은 산도에서 2023년 pH 4.79로 개선되었으나 밭토양 pH 5.80과 미경작지 토양 pH 5.48보다 다소 낮은 산성토양을 보였으며, 토양배수가 양호한 사질토양 또는 식토양의 특성을 지니고 있다(Park, 2024).
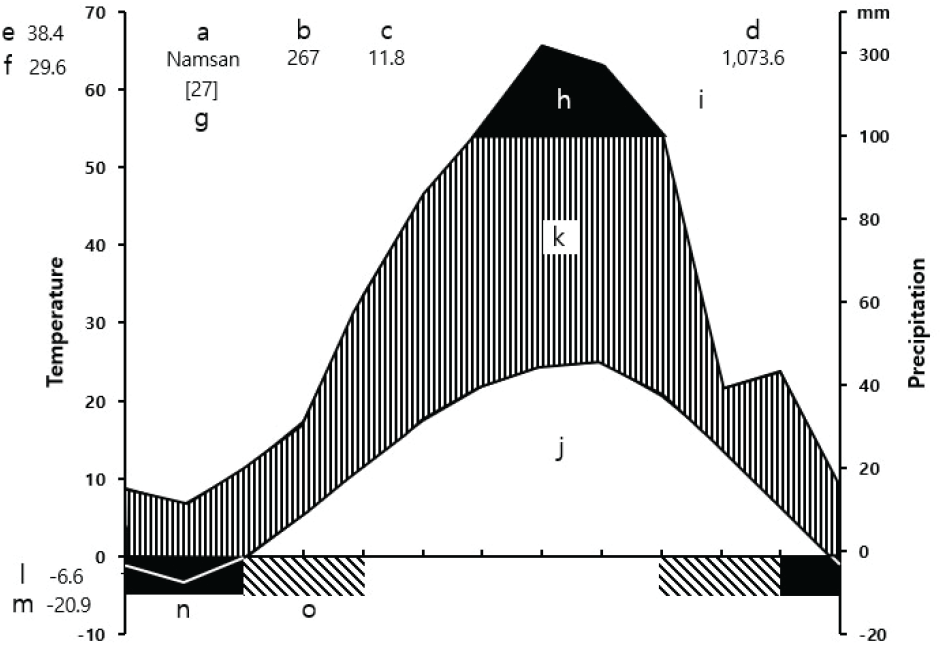
조사지와 인접한 서울특별시 중구(419) 지점의 기상청 자동기상관측장비(AWS: Automatic Weather System)에 기록된 기상자료(KMA, 2024)를 이용하여 Walter et al.(1975)이 제시한 기후도를 작성한 결과, 최근 27년간(1997년~2023년) 중구에서의 연평균 기온은 11.8°C이며, 연평균강수량은 1,073.6 mm이었다. 강수량이 집중된 달은 7월과 8월이었다. 가장 더운 달의 일평균 최고기온은 29.6°C, 가장 추운 달의 일평균 최저기온은 −6.6°C로 각각 나타났고, 절대 최고기온은 38.4°C, 절대 최저기온은 –20.9°C로 각각 나타났다(Figure 2).
Figure 2.
Climate diagram of Jung-gu, Seoul(419).
a: Station name, b: Height above sea level (m). c: Mean annual temperature(°C), d: Mean annual amount of precipitation (mm), e: Absolute maximum temperature (°C), f: Mean daily maximum of the hottest month (°C), g: Number of years observation, h: Average monthly precipitation exceeding 100 mm (black area), i: Monthly means of precipitation (mm), j: Monthly means of temperature (°C), k: Humid period (lined), l: Mean daily minimum temperature of the coldest month (°C), m: Absolute minimum temperature (°C), n: Months with a mean, daily minimum temperature below 0°C, o: Months with an absolute minimum below.
Download Original Figure
2. 연구 방법
1) 조사구 설치 및 조사 방법
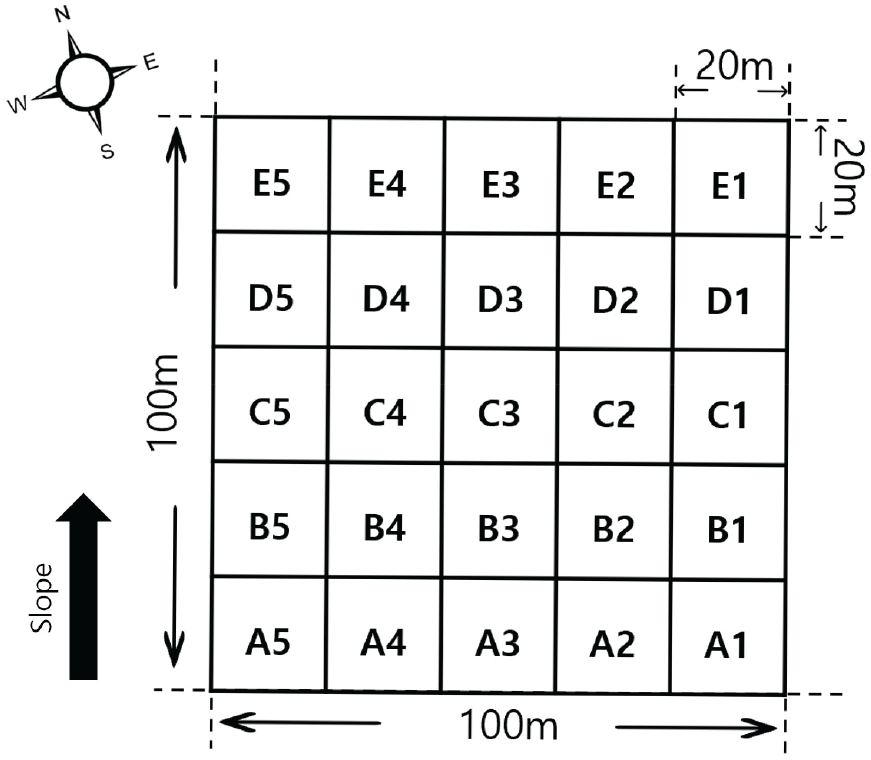
본 조사는 2017년 5월부터 2023년 5월까지 남산의 장기생태모니터링 구간 신갈나무림 1 ha를 조사구로 선정하였다. 장기생태연구사업의 영구방형구 설치방법에 따라 각 조사구마다 동일한 등고선 방향을 유지하도록 영구방형구를 설치하였으며 영구방형구의 끝모서리 네 군데를 각각 GPS를 사용하여 동서남북의 방향을 잡아주었다(NIER, 2013). 영구방형구 내 한쪽 모서리로부터 20 m ×20 m의 소방형구를 설치한 후 계속 연결하여 총 25개의 소방형구를 설치하였다(Figure 3). 각 소방형구 내의 개체목에 고유번호가 적힌 테그를 흉고높이(1.2 m) 지점에 부착하였으며, 매목조사는 영구방형구 내 층위에 따른 구분과 함께 흉고직경 2 cm 이상의 개체목에 대한 종명, 수고, 흉고직경, 수간건강상태, 수관활력도 등을 측정하여 기록하였다.
조사구 내 출현하는 모든 수종에 대한 동정은 원색대한식물도감(Lee, 2003), 나무생태도감(Yun, 2022)을 기준으로 하였으며, 국명과 학명은 국가생물종목록(NIBR, 2023)을 기준으로 하여 작성하였다.
수고의 경우 A~E등급(A: 5 m 미만, B: 5-10 m, C: 10-15 m, D: 15-20 m, E: 20 m 이상)을 기준으로 하여 기록하였으며, 직경테이프를 사용하여 개체목 마다 흉고직경을 측정하였다. 수간건강상태는 사목과 생목을 구분하고 수간상태에 따라 9개 수간건강상태 유형(AS: Alive Standing; AB: Alive Broken; AD: Alive Deadtop; AF: Alive Fallen; AL: Alive Leaning; DS: Dead Standing; DB: Dead Broken; DL: Dead Leaning; DF: Dead Fallen)을 기준으로 하여 기록하였다. 수관활력도는 개체목의 수관에 달린 잎의 양을 조사하여 5개의 등급으로(활력도 1: 식물체에 달린 잎의 양이 10% 미만, 활력도 2: 식물체에 달린 잎의 양이 10-40%, 활력도 3: 식물체에 달린 잎의 양이 40-60%, 활력도 4: 식물체에 달린 잎의 양이 60-90%, 활력도 5: 식물체에 달린 잎의 양이 90% 초과) 기록하였다.
2) 분석 방법
(1) 중요치
중요치는 개체목들의 상대적인 생태적 지위를 파악하기 위해서 조사지 내 수종을 대상으로 Curtis and McIntosh의 방법을 이용하여 상대밀도, 상대빈도, 상대피도를 합산하여 중요치(Importance value; I.V)를 산출하였다(Curtis and McIntosh, 1950, 1951).
식에서, I.V = 중요치(Importance value) : 상대밀도(RDi), 상대빈도(RFi), 상대피도(RCi)를 합산하여 평균 낸 비율
RDi = ni/∑n : 출현한 모든 종의 총개체수(∑n)에 대한 i종 개체수(ni)의 비율,
RCi = Ci/∑C : 출현한 모든 종의 총피도의 합(∑C)에 대한 i종의 피도(Ci)의 비율,
RFi = Fi/∑F : 출현한 모든 종의 빈도의 합(∑F)에 대한 i 종의 빈도(Fi)의 비율
2) 밀도 및 흉고단면적
조사구 내 구성목의 밀도는 2017년, 2019년, 2021년, 2023년에 살아있는 개체군별 개체수를 나타내었으며, 신갈나무림 내 기타 수종을 제외한 대표적인 15종의 밀도 데이터를 가지고 연도별로 신갈나무림의 밀도 변화가 유의미한지 알아보고자 비모수통계법인 Friedman-test (Friedman, 1937)를 R 4.4.2 통계프로그램을 통해 통계검증을 실시하였다.
한 개체목의 흉고단면적(Breast height area; BHA)은 아래의 식과 같이 계산하였고 조사년도에 따른 개체군별 흉고단면적 합계를 나타내었다.
식에서, BHA: 흉고단면적, d: 흉고직경(cm)
3) 흉고직경급 분포
직경급 분포는 살아있는 목본을 대상으로 2-5 cm, 5-10 cm, 10-20 cm, 20-30 cm, 30-40 cm, 40-50 cm, 50-60 cm, 60-70 cm, 70 cm 이상 총 9개의 직경급으로 분류하여 임목 본수를 각 조사 연도별로 나타내었다.
4) 고사율과 이입률
조사구 내 각 군락별 주요 수종의 고사율(mortality rate)과 이입률(recruitment rate)은 고사목과 진계목을 구분하여 분석하였으며, 이 방법은 개체목 집단의 사망률이 시간이 지남에 따라 기하급수적으로 감소하는 것을 가정하고 있다(Sheil, 1995).
Nt= 이전 측정에서 살아 있는 개체목 수 중 현재 측정에서 살아 있는 개체목 수
No= 이전 측정에서 살아있는 개체목 수
t = 측정사이의 기간
Nt= 현재 측정에서 추가된 개체목 수
No= 이전 측정에서 살아있는 개체목 수
t = 측정사이의 기간
5) 임목 쇠퇴율
임목쇠퇴율은 임목 쇠퇴도를 조사지 내 각 입목(생육목, 고사목)을 대상으로 수관활력도와 수간건강상태를 수관활력지표(Ha), 수간건강지표(Hb)로 변환하여 산출한 후(Lim et al., 2019; Park et al., 2022), 임목 쇠퇴도를 Percentage(%)로 환산하여 임목 쇠퇴율을 구하였다.
(Ha: 수관활력지표, Hb: 수간건강지표)
결과 및 고찰
1. 밀도 및 고사율과 이입률
남산 신갈나무림의 2017년, 2019년, 2021년, 2023년 밀도 및 고사율과 이입률은 다음과 같다(Table 1,2, Figure 4). 개체군 별 밀도는 당단풍나무, 때죽나무, 신갈나무, 팥배나무 등의 순으로, 당단풍나무의 밀도가 가장 높게 나타났다. 전체 구성 종의 밀도는 2017년 1,117 stems ha−1에서 96 stems ha−1(이입률 4.4%)의 진계목과 47 stems ha−1(고사율 2.1%)의 고사목 발생으로 인하여 2019년 1,166 stems ha−1로 변화하였다. 2017년에서 2019년 사이 5 cm 미만의 당단풍나무 진계목이 다수 발생함에 따라 전체 이입률이 크게 증가하였다. 2019년에서 2021년 사이 1,170 stems ha−1로 변화하였으며, 이는 30 stems ha−1(1.3%)의 진계목과 26 stems ha−1(1.1%)의 고사목 발생으로 인한 것으로 나타났다. 2021년에서 2023년은 1,108 stems ha−1로 변화하였으며, 34 stems ha−1의 진계목(1.5%)과 96 stems ha−1(4.2%)의 고사목 발생으로 인하여 변화하였다.
Table 1.
Density for living stems in Q. mongolica community and Friedman-Test Statistics
|
Species |
|
Density (stems ha−1) |
|
Scientific name (Korean name) |
|
2017yr
|
2019yr
|
2021yr
|
2023yr
|
|
Quercus mongolica (신갈나무) |
|
187 |
176 |
170 |
169 |
|
Acer pseudosieboldianum (당단풍나무) |
|
350 |
381 |
396 |
405 |
|
Styrax japonicus (때죽나무) |
|
255 |
261 |
259 |
248 |
|
Sorbus alnifolia (팥배나무) |
|
177 |
177 |
175 |
162 |
|
Alnus hirsuta (물오리나무) |
|
16 |
22 |
23 |
22 |
|
Q. serrata (졸참나무) |
|
20 |
20 |
20 |
20 |
|
Euonymus oxyphyllus (참회나무) |
|
52 |
61 |
57 |
12 |
|
Robinia pseudoacacia (아까시나무) |
|
6 |
8 |
10 |
10 |
|
Kalopanax septemlobus (음나무) |
|
6 |
7 |
7 |
9 |
|
Cornus controversa (층층나무) |
|
7 |
8 |
9 |
9 |
|
Fraxinus rhynchophylla (물푸레나무) |
|
4 |
4 |
5 |
5 |
|
Maackia amurensis (다릅나무) |
|
2 |
2 |
2 |
4 |
|
Magnolia kobus (목련) |
|
2 |
4 |
4 |
4 |
|
M. obovata (일본목련) |
|
2 |
3 |
3 |
3 |
|
Q. aliena (갈참나무) |
|
1 |
1 |
1 |
1 |
|
Others(10) |
|
30 |
31 |
29 |
25 |
|
Total |
|
1,117 |
1,166 |
1,170 |
1,108 |
|
Friedman Test Statistics |
|
χ2(3) = 8.2703 (p<0.05) |
Download Excel Table
Table 2.
The biannual change of recruitment rate, mortality rate in Q. mongolica community.
|
Species |
Mortality rate (% year−1) |
|
Recruitment rate (% year−1) |
|
’17-’19 |
’19-’21 |
’21-’23 |
’17-’23 |
|
’17-’19 |
’19-’21 |
’21-’23 |
’17-’23 |
|
Quercus mongolica (신갈나무) |
3.5 |
1.7 |
1.2 |
2.2 |
|
0.5 |
0.0 |
0.9 |
0.5 |
|
Acer pseudosieboldianum (당단풍나무) |
0.4 |
0.0 |
0.3 |
0.2 |
|
5.0 |
2.0 |
1.4 |
3.1 |
|
Aria alnifolia (팥배나무) |
1.1 |
0.9 |
3.8 |
2.0 |
|
1.1 |
0.3 |
0.0 |
0.5 |
|
Styrax japonicus (때죽나무) |
4.6 |
1.7 |
5.2 |
4.2 |
|
5.9 |
1.4 |
2.9 |
3.7 |
|
Q. serrata (졸참나무) |
0.0 |
0.0 |
0.0 |
0.0 |
|
0.0 |
0.0 |
0.0 |
0.0 |
|
Alnus hirsuta (물오리나무) |
0.0 |
0.0 |
2.2 |
1.1 |
|
20.9 |
2.3 |
0.0 |
9.1 |
|
Prunus spp. (벚나무류) |
7.8 |
8.2 |
6.1 |
8.2 |
|
5.1 |
2.7 |
0.0 |
2.7 |
|
Kalopanax septemlobus (음나무) |
0.0 |
0.0 |
0.0 |
0.0 |
|
8.7 |
0.0 |
15.5 |
10.9 |
|
Robinia pseudoacacia (아까시나무) |
0.0 |
0.0 |
0.0 |
0.0 |
|
18.4 |
13.4 |
0.0 |
16.7 |
|
Euonymus oxyphyllus (참회나무) |
0.0 |
4.2 |
54.1 |
41.9 |
|
9.1 |
0.8 |
0.0 |
3.5 |
|
Cornus controversa (층층나무) |
0.0 |
0.0 |
5.7 |
2.5 |
|
7.4 |
6.5 |
5.7 |
8.9 |
|
Fraxinus rhynchophylla (물푸레나무) |
0.0 |
0.0 |
0.0 |
0.0 |
|
0.0 |
13.4 |
0.0 |
4.7 |
|
Magnolia kobus (목련) |
0.0 |
0.0 |
0.0 |
0.0 |
|
100.0 |
0.0 |
0.0 |
100.0 |
|
M. obovata (일본목련) |
0.0 |
0.0 |
0.0 |
0.0 |
|
29.3 |
0.0 |
0.0 |
10.9 |
|
Maackia amurensis (다릅나무) |
0.0 |
0.0 |
0.0 |
0.0 |
|
0.0 |
0.0 |
100.0 |
100.0 |
|
Q. aliena (갈참나무) |
0.0 |
0.0 |
0.0 |
0.0 |
|
0.0 |
0.0 |
0.0 |
0.0 |
|
Picrasma quassioides (소태나무) |
0.0 |
0.0 |
18.4 |
10.9 |
|
29.3 |
0.0 |
0.0 |
10.9 |
|
Pinus densiflora (소나무) |
0.0 |
0.0 |
0.0 |
0.0 |
|
0.0 |
0.0 |
0.0 |
0.0 |
|
P. padus (귀룽나무) |
0.0 |
0.0 |
0.0 |
0.0 |
|
0.0 |
0.0 |
0.0 |
0.0 |
|
P. koraiensis (잣나무) |
0.0 |
0.0 |
0.0 |
0.0 |
|
0.0 |
0.0 |
0.0 |
0.0 |
|
Paulownia coreana (오동나무) |
0.0 |
0.0 |
0.0 |
0.0 |
|
0.0 |
0.0 |
0.0 |
0.0 |
|
Morus bombycis (산뽕나무) |
29.3 |
0.0 |
0.0 |
10.9 |
|
0.0 |
0.0 |
0.0 |
0.0 |
|
Ailanthus altissima (가죽나무) |
0.0 |
0.0 |
29.3 |
10.9 |
|
0.0 |
0.0 |
0.0 |
0.0 |
|
Castanea crenata (밤나무) |
0.0 |
0.0 |
0.0 |
0.0 |
|
0.0 |
0.0 |
0.0 |
0.0 |
|
Juniperus rigida (노간주나무) |
0.0 |
0.0 |
0.0 |
0.0 |
|
0.0 |
0.0 |
0.0 |
0.0 |
|
Total |
2.1 |
1.1 |
4.2 |
2.7 |
|
4.4 |
1.3 |
1.5 |
2.5 |
Download Excel Table
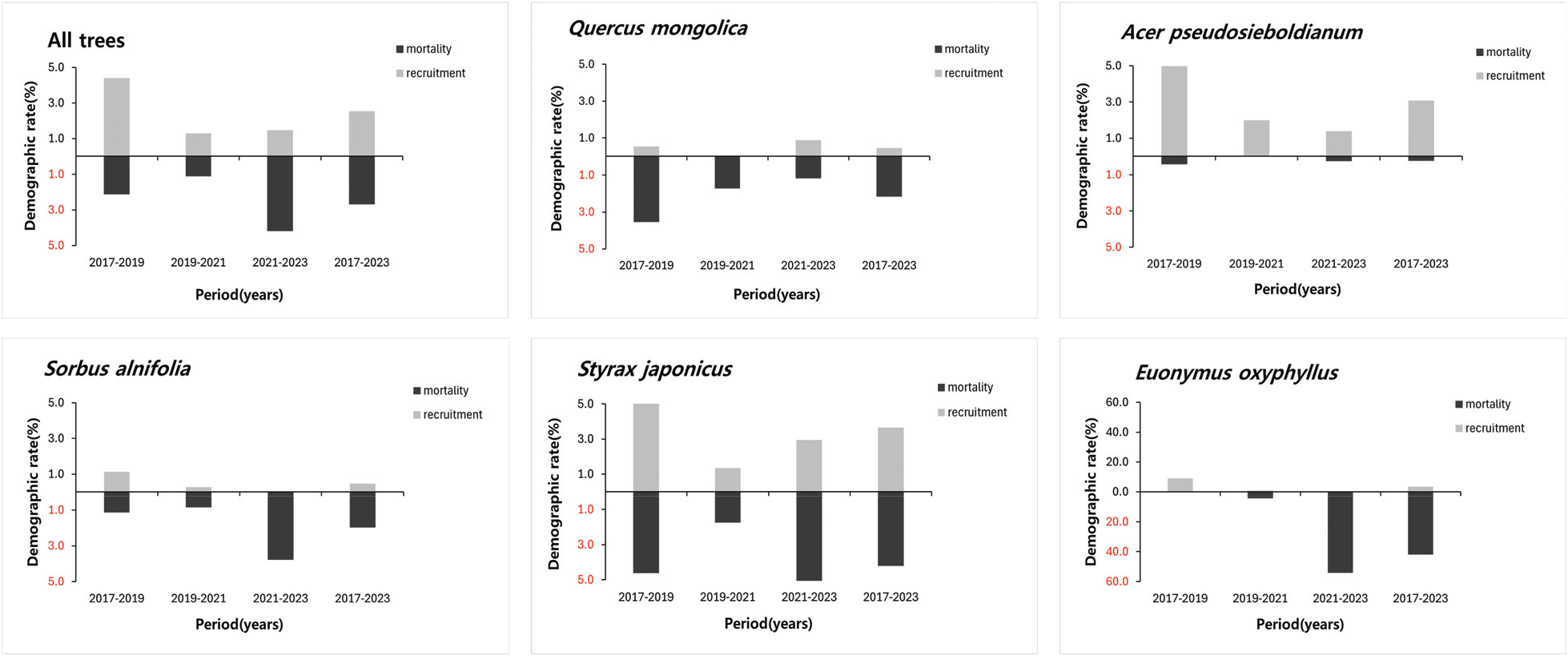
Figure 4.
Mortality and recruitment of Q. mongolica community and major tree species of the community.
Download Original Figure
신갈나무림 내 신갈나무 밀도는 2017년 187 stems ha−1에서 2023년 169 stems ha−1로 2017년 대비 2023년 5 stems ha−1의 진계목과 23 stems ha−1의 고사목(이입률 0.5%, 고사율 2.2%)이 발생하였으며, 팥배나무는 177 stems ha−1에서 162 stems ha−1으로 2017년 대비 2023년 5 stems ha−1의 진계목과 20 stems ha−1의 고사목(이입률 0.5%, 고사율 2.0%) 발생하였으며 두 개체군 모두 비교적 세력이 유지되고 있는 것으로 판단되었다. 당단풍나무는 2017년 350 stems ha−1에서 2023년 405 stems ha−1로 2017년 대비 2023년 60 stems ha−1의 진계목과 5 stems ha−1의 고사목(이입률 3.1%, 고사율 0.2%)이 발생하여 개체목이 지속적으로 증가하고 있었다. 때죽나무는 255 stems ha−1에서 248 stems ha−1로 2017년 대비 2023년 51 stems ha−1의 진계목과 58 stems ha−1의 고사목(이입률 3.7%, 고사율 4.2%)이 발생하였다. 때죽나무는 비교적 다른 수종에 비하여 내한성과 내공해성이 높은 특성(Woo et al., 2000; Yun, 2022)과 벌도목에서 근주 맹아가 빠르게 발달 특성(Jee and Oh, 2001)이 있다. 이러한 특성으로 인해 벌채목들이 있는 소방형구(Figure 5)는 임상(林床)에 광량이 증가하여 때죽나무 진계생장이 유리했던 것으로 판단되었다. 때죽나무 고사목의 경우 DBH 5 cm 미만인 고사목이 대부분이며, 수간건강상태가 DS유형(서서 고사한 개체목)과 DF유형(쓰러져서 고사한 개체목)으로, 당단풍나무와의 종간 경쟁에 의하여 생육상태가 저조하였던 개체목 일부가 고사한 것으로 판단되었다. 반면, 졸참나무는 2017년부터 2023년까지 밀도(20 stems ha−1)가 유지되었다. 일본목련과 같은 외래수종의 경우 밀도가 증가 후 유지되고 있었다. 남산 북사면 신갈나무림에서는 참나무시들음병과 인위적 훼손이 발생한 지역을 중심으로 가중나무, 참오동나무, 일본목련 등의 외래종 치수가 생육하고 있었다(No, 2014). 본 연구에서도 일부 외래수종이 유입되었으나 밀도가 크지 않고 증가보다는 유지되는 경향을 보여, 당분간 외래수종으로 인한 신갈나무림의 개체군 변화는 크지 않을 것으로 판단되었다.
Figure 5.
Photos (a) and (b) show traces of logged trees resulting from forest management within the Q. mongolica community in Mt. Namsan.
Download Original Figure
Friedman-test를 R 4.42 프로그램을 통해 통계 검정을 실시하였으며, Friedman Test 결과, 연도별 밀도의 변화는 통계적으로 유의미하게 나타났다(p-value = 0.04074).
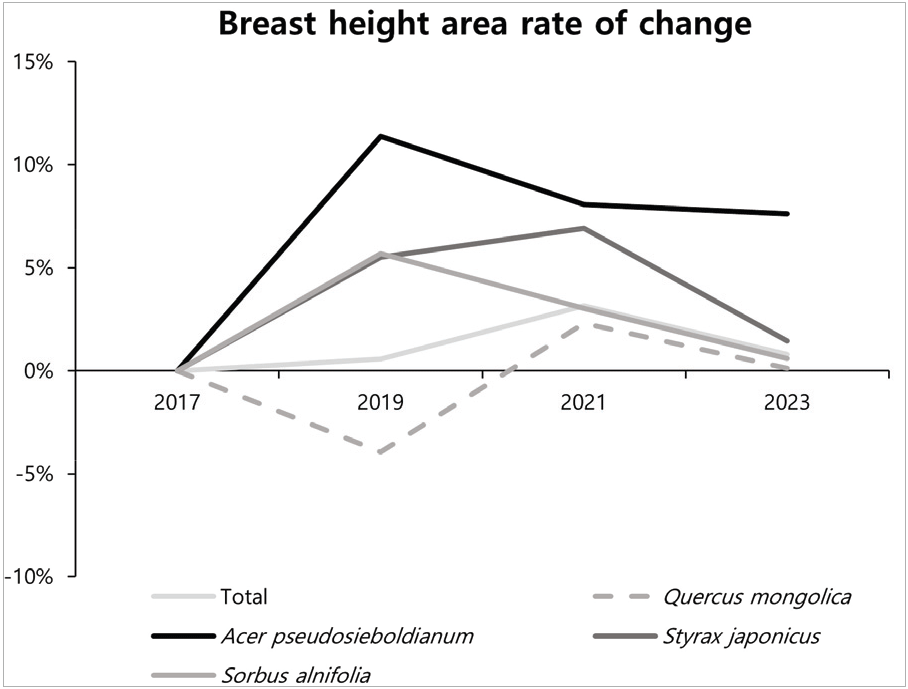
2. 흉고단면적 생장 변화
신갈나무림 전체 구성종의 흉고단면적은 2017년 21.0 m2·ha−1, 2019년 21.1 m2·ha−1(+0.6%), 2021년 21.8 m2·ha−1 (+3.1%), 2023년 21.9 m2·ha−1(+0.8%)로 증가하는 경향이었다. 신갈나무 개체군의 흉고단면적 변화는 2017년 10.0 m2·ha−1, 2019년 9.6 m2·ha−1(−3.9%), 2021년 9.8 m2·ha−1 (+2.3%), 2023년 9.8 m2·ha−1(+0.1%)로 나타났으며, 연도별 증감율은 2019년 감소 후 다시 증가하였다. 이는 2019년에 전년도의 태풍 영향에 따른 쓰러져서 고사한 개체(DF), 부러져서 고사한 개체(DB) 증가와 몇몇 개체목이 벌채됨에 따라 밀도가 감소되었다. 당단풍나무는 2017년 1.3 m2·ha−1, 2019년 1.5 m2·ha−1(+11.3%), 2021년 1.6 m2·ha−1 (+8.1%), 2023년 1.7 m2·ha−1(+7.7%)로, 팥배나무는 2017년 2.8 m2·ha−1, 2019년 3.0 m2·ha−1(+5.6%), 2021년 3.1 m2·ha−1(+3.0%), 2023년 3.1 m2·ha−1(+0.6%)로, 2017년부터 2023년까지 증가하였다. 때죽나무는 2017년 0.6 m2·ha−1, 2019년 0.7 m2·ha−1(+5.6%), 2021년 0.7 m2·ha−1(+6.8%), 2023년 0.7 m2·ha−1(+1.6%)로 졸참나무는 2017년 2.9 m2·ha−1, 2019년 3.0 m2·ha−1(+2.7%), 2021년 3.1 m2·ha−1(+4.4%), 2023년 3.2 m2·ha−1(+3.1%)로 아교목성 수종인 당단풍나무, 팥배나무, 때죽나무 모두 흉고단면적이 증가하고 있는 것으로 보아 신갈나무림 내 아교목성 수종들의 생육이 양호함에 따라 직경이 증가하여 흉고단면적이 증가한 것으로 판단되었다. 참회나무의 흉고단면적은 2021년 참회나무 개체수의 대부분인 5 cm 미만의 직경급들이 다량 고사함에 따라 흉고단면적도 크게 감소하는 것으로 나타났다(Table 3, Figure 6).
Table 3.
Breast height area for living stems of Q. mongolica community.
|
Species |
|
Breast height area (m2 ha−1) |
|
Scientific name (Korean name) |
|
2017yr
|
2019yr
|
2021yr
|
2023yr
|
|
Quercus mongolica (신갈나무) |
|
9.996 |
9.601 |
9.821 |
9.834 |
|
Acer pseudosieboldianum (당단풍나무) |
|
1.345 |
1.497 |
1.618 |
1.742 |
|
Styrax japonicus (때죽나무) |
|
0.626 |
0.661 |
0.706 |
0.717 |
|
Sorbus alnifolia (팥배나무) |
|
2.835 |
2.996 |
3.086 |
3.104 |
|
Alnus hirsuta (물오리나무) |
|
1.238 |
1.288 |
1.327 |
1.388 |
|
Q. serrata (졸참나무) |
|
2.901 |
2.980 |
3.112 |
3.208 |
|
Euonymus oxyphyllus (참회나무) |
|
0.078 |
0.093 |
0.078 |
0.013 |
|
Robinia pseudoacacia (아까시나무) |
|
0.044 |
0.052 |
0.063 |
0.073 |
|
Kalopanax septemlobus (음나무) |
|
0.738 |
0.751 |
0.759 |
0.775 |
|
Cornus controversa (층층나무) |
|
0.051 |
0.055 |
0.063 |
0.069 |
|
Fraxinus rhynchophylla (물푸레나무) |
|
0.144 |
0.147 |
0.152 |
0.155 |
|
Maackia amurensis (다릅나무) |
|
0.106 |
0.109 |
0.119 |
0.122 |
|
Magnolia kobus (목련) |
|
0.002 |
0.006 |
0.012 |
0.019 |
|
M. obovata (일본목련) |
|
0.008 |
0.015 |
0.023 |
0.031 |
|
Q. aliena (갈참나무) |
|
0.163 |
0.187 |
0.189 |
0.199 |
|
Others(10) |
|
0.720 |
0.675 |
0.648 |
0.500 |
|
Total |
|
20.995 |
21.113 |
21.776 |
21.949 |
Download Excel Table
Figure 6.
The changes in the Breast height area growth rate of the Q. mongolica community and its major species (Q. mongolica, A. pseudosieboldianum, S. japonicus, and S. alnifolia).
Download Original Figure
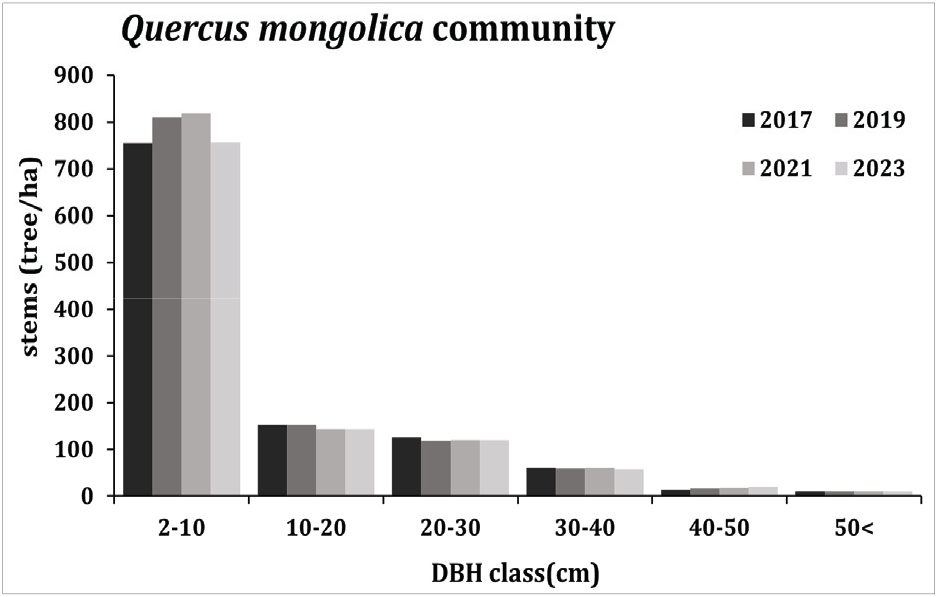
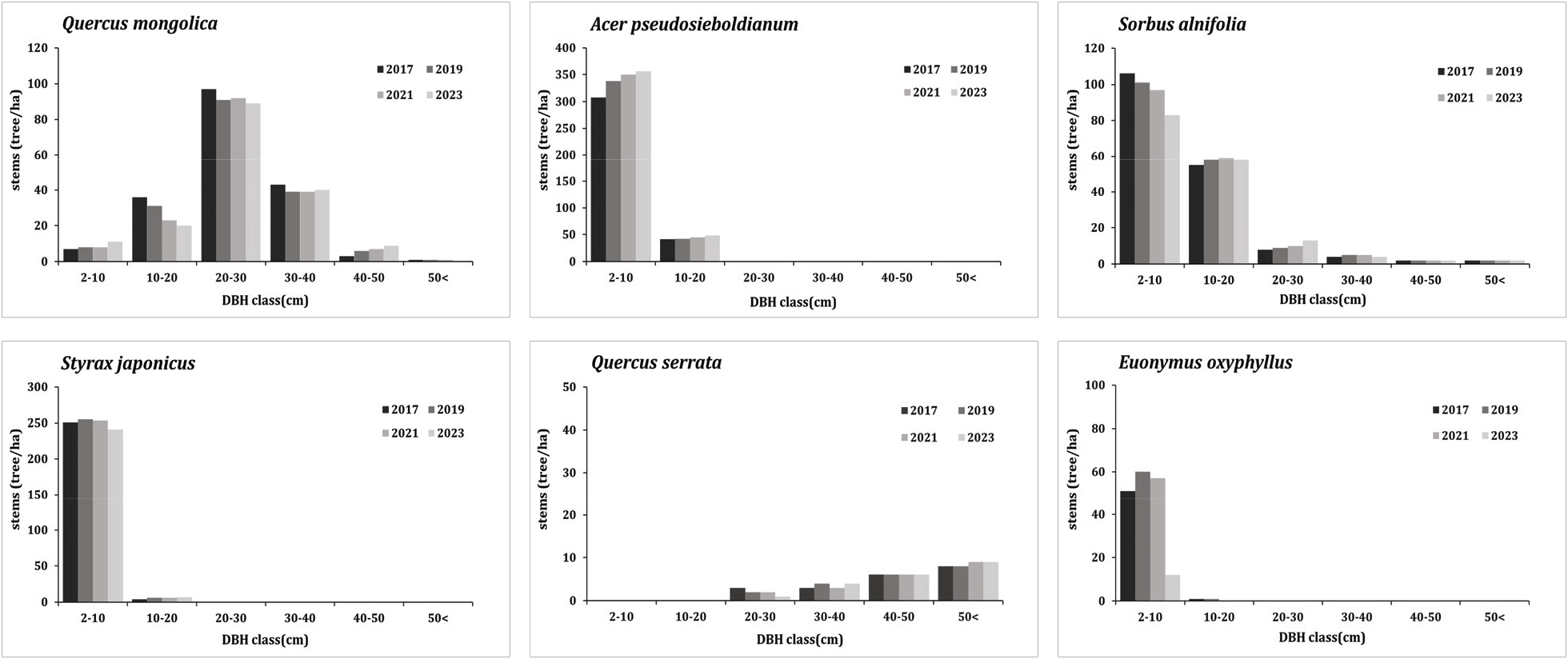
3. 흉고직경급 분포 변화
신갈나무림의 조사구에서 매목조사를 실시하여 얻어진 자료를 기초로 하여 직경급 구조를 나타낸 결과(Figure 7, 8), 신갈나무림 전체 구성종은 5 cm 미만과 5-10 cm의 직경급이 다른 직경급보다 개체수가 많고 직경급이 높아질수록 밀도가 낮아지는 역 J자 구조를 가지고 있었다. 직경급분포그래프에서 역 J자형태를 보이고 있음은 동령림의 경우 경쟁이 일어나고 있음을 의미하며 동령림이 아닌 이령림의 경우 침입집단 또는 극상림 집단의 특징으로서 지속적으로 유지될 수 있는 집단을 의미한다(Barbour et al., 1987; Lee et al., 2007). 5 cm 미만의 직경급에서는 연도별로 증가와 감소를 반복하고 있었으며 5-10 cm 직경급에서는 연도별로 점차 증가하고 있었는데 이러한 흉고직경급 패턴은 활엽수 혼효림에서 주로 발견되는 구조로 다양한 층위로 구성된 임분 형태를 나타내고 있는 것으로 해석할 수 있다(Smith and Hawley, 1986). 신갈나무림 전체 구성종의 5 cm 미만 직경급에서 2021년에서 2023년 사이 다소 크게 감소한 것은 참회나무, 팥배나무, 때죽나무 5 cm 미만 직경급의 개체목 다량 고사 때문으로 판단되었다.
Figure 8.
The biannual change of DBH class distribution for major species population(5) and Euonymus oxyphyllus Population in Mt. Namsan.
Download Original Figure
신갈나무 개체군의 직경급 구조는 극상림으로서 역 J자의 구조를 보이는 형태가 아닌 20-30 cm 직경급에서 모든 조사연도를 통틀어 가장 높은 정규분포(단봉형)의 형태를 보였다. 이는 후계목 발생에 있어 불리한 것으로 판단되나(Park, 2019) 5 cm 미만 5-10 cm 직경급에서 개체 밀도가 비교적 안정적으로 나타나고 있어 특별한 외부적 간섭이 없는 신갈나무가 우점하는 상태로 유지될 것으로 판단되었다(Kim, 2016). 신갈나무의 10-20 cm 직경급의 경우 연도별 개체 밀도가 감소하는 추세로 보이지만 이는 2017년 조사부터 생육상태 및 수관활력이 좋지 못했던 몇몇 개체들이 고사하여 개체 밀도가 감소한 것으로 이를 제외한 대부분의 개체목의 생육상태는 특별한 변동이 없었다. 20 cm 이하의 직경급에서 역 J자 구조를 보이는 아교목성 수종의 개체군은 당단풍나무, 때죽나무, 팥배나무이며 그중 당단풍나무와 팥배나무는 추후 아교목층의 신갈나무와의 경쟁 우위관계에 있을 것으로 판단되었다.
4. 군집 구성종의 중요치
수종 별 중요치는 신갈나무(24.5), 당단풍나무(18.0), 팥배나무(14.2), 때죽나무(13.0) 순으로 높게 나타났다(Table 4). 신갈나무의 중요치는 2017년 대비 2023년에 16% 감소하였으며, 2017년부터 2021년까지 지속적으로 감소한 후 다시 증가하였다. 이 중 2017년에서 2019년 사이에 가장 큰 감소가 나타났다. 이러한 감소는 조사 시기 당시 30-40 cm 직경급에 해당하는 일부 개체목이 고사함에 따른 결과로 나타났다. 고사의 원인은 태풍의 바람 교란으로 큰 개체들이 죽게 되었고, 중요치가 감소하게 되었다(Figure 9). 당단풍나무는 2017년 대비 2023년에 2.0% 증가하였으며 2017년부터 2023년까지 지속적으로 증가하고 있었다. 당단풍나무는 신갈나무와 다양한 입지 조건을 공유하는 공통 출현종이며(Kim and Kil, 2000; Park and Yun, 2009; Song et al., 2003; Yun et al., 2011; Kim et al., 2018; Kim et al., 2021), 강한 내음성을 통해 임분 하층에 많이 자라는 대표 수종이다(Yun, 2022). 이러한 당단풍나무의 생태 특성이 위의 중요치 결과를 나타낸 배경으로 판단된다. 팥배나무는 2017년 대비 2023년 0.5% 감소하였으며, 때죽나무는 2017년 대비 2023년 0.1% 감소하였다. 팥배나무는 2017년부터 2021년까지 감소 후 유지되었고 때죽나무는 2017년부터 2021년까지 감소 후 다시 증가하였다. 두 개체군 모두 2017년부터 생육 상태가 좋지 않은 개체목들이 시간이 지남에 따라 서서히 고사함에 따른 결과로, 고사한 개체목들 중 직경 10 cm이하의 비율이 가장 높았다.
Table 4.
Importance value of component species in the Q. mongolica community.
|
Species |
|
IV |
Mean |
|
Scientific name (Korean name) |
|
2017yr
|
2019yr
|
2021yr
|
2023yr
|
|
Quercus mongolica (신갈나무) |
|
26.0 |
24.5 |
24.1 |
24.4 |
24.5 |
|
Acer pseudosieboldianum (당단풍나무) |
|
17.4 |
17.8 |
18.1 |
19.4 |
18.0 |
|
Sorbus alnifolia (팥배나무) |
|
14.6 |
14.3 |
14.1 |
14.1 |
14.2 |
|
Styrax japonicus (때죽나무) |
|
13.2 |
12.8 |
12.7 |
13.1 |
13.0 |
|
Q. serrata (졸참나무) |
|
7.3 |
7.3 |
7.3 |
7.5 |
7.3 |
|
Alnus hirsuta (물오리나무) |
|
4.4 |
4.8 |
5.0 |
5.1 |
4.9 |
|
Kalopanax septemlobus (음나무) |
|
2.3 |
2.5 |
2.4 |
2.5 |
2.4 |
|
Robinia pseudoacacia (아까시나무) |
|
1.0 |
1.4 |
1.8 |
1.9 |
1.6 |
|
Euonymus oxyphyllus (참회나무) |
|
4.2 |
4.2 |
4.2 |
1.5 |
4.2 |
|
Cornus controversa (층층나무) |
|
1.2 |
1.2 |
1.4 |
1.5 |
1.3 |
|
Fraxinus rhynchophylla (물푸레나무) |
|
0.9 |
0.9 |
1.1 |
1.1 |
1.0 |
|
Magnolia kobus (목련) |
|
0.4 |
0.7 |
0.7 |
0.7 |
0.7 |
|
M. obovata (일본목련) |
|
0.5 |
0.6 |
0.6 |
0.7 |
0.6 |
|
Maackia amurensis (다릅나무) |
|
0.4 |
0.4 |
0.4 |
0.7 |
0.4 |
|
Q. aliena (갈참나무) |
|
0.5 |
0.5 |
0.5 |
0.5 |
0.5 |
|
Others (10) |
|
5.7 |
6.1 |
5.7 |
5.3 |
5.7 |
|
Total |
|
100 |
100 |
100 |
100 |
100 |
Download Excel Table
Figure 9.
Photos of dead standing and dead broken tree in the Q. mongolica community on Mt. Namsan.
Download Original Figure
팥배나무의 경우 직경 30 cm이상의 개체목들은 대부분 4~5의 활력도를 가진 건전한 생장 상태를 보였다. 이는 서울의 환경에서 도심 팥배나무들은 자연지역보다 둥글고 무거운 잎을 생산하는 형질변이 특성에 기반한 것으로 보인다(Jung et al., 2021). 때죽나무 역시 직경 10-20 cm의 개체목들은 대부분 3~4의 활력도를 가지고 있었으며 이는 때죽나무가 대기오염 등의 공해에 잘 적응하여 생존하는 특성이 비교적 성숙된 개체목들에 있어 반영된 것으로 보인다(Woo et al., 2000; Kwak, 2011). 참회나무는 2017년 대비 2023년 2.7%로 크게 감소하는 모습을 보였다. 참회나무와 당단풍나무는 서로 종간연관성이 높은데 (Cheon et al., 2014) 당단풍나무와의 지위경쟁에서 탈락된 개체들이 고사하게 됨에 따라 참회나무의 중요치에 영향이 미친 것으로 판단되었다. 중요치를 기준으로 볼 때 감소 추세를 보이는 개체군은 참회나무, 유지되는 개체군은 신갈나무, 팥배나무, 때죽나무, 증가하는 개체군은 당단풍나무인 것으로 나타났다.
임분에 발생한 자연적 및 인위적 교란은 수관구조를 변화시켜 임분 내 광환경을 변화시키는 요인이 된다(Majasalmi and Rautiainen, 2020; Castorani et al., 2021). 본 조사지에서도 이와 같은 교란은 수관 개방도를 증가시켜 하층 식생에 더 많은 빛이 도달하도록 하였으며, 이로 인해 광요구도가 높은 비료목류의 생장 조건을 유리하게 조성했을 가능성이 있다.
실제로 본 연구에서 아까시나무와 물오리나무와 같은 비료목류는 시간이 지남에 따라 중요치가 증가하는 경향을 보였다. 이들 수종은 건조, 산성화 등 불량 환경에서도 잘 적응하여 생육할 수 있는 생태적 특성(Clark, 1986; Rédei et al., 2002; Jung et al., 2011)이 있으며, 이들은 질소 고정 능력을 통해 지력 회복 및 토양 미생물 활성화에도 기여하며, 척박한 토양에서의 생태적 경쟁력을 보유하고 있다(Arnone, and Gordon, 1990; Huss-danell and Sellstedt, 1985; Lambers et al., 2008; Choi and Toda, 2012). 따라서 임분 내 수관 교란이 발생한 이후 비료목류의 중요치가 증가한 것은 광환경 개선과 이들 수종의 기능적 생태 특성이 복합적으로 작용한 결과로 해석된다.
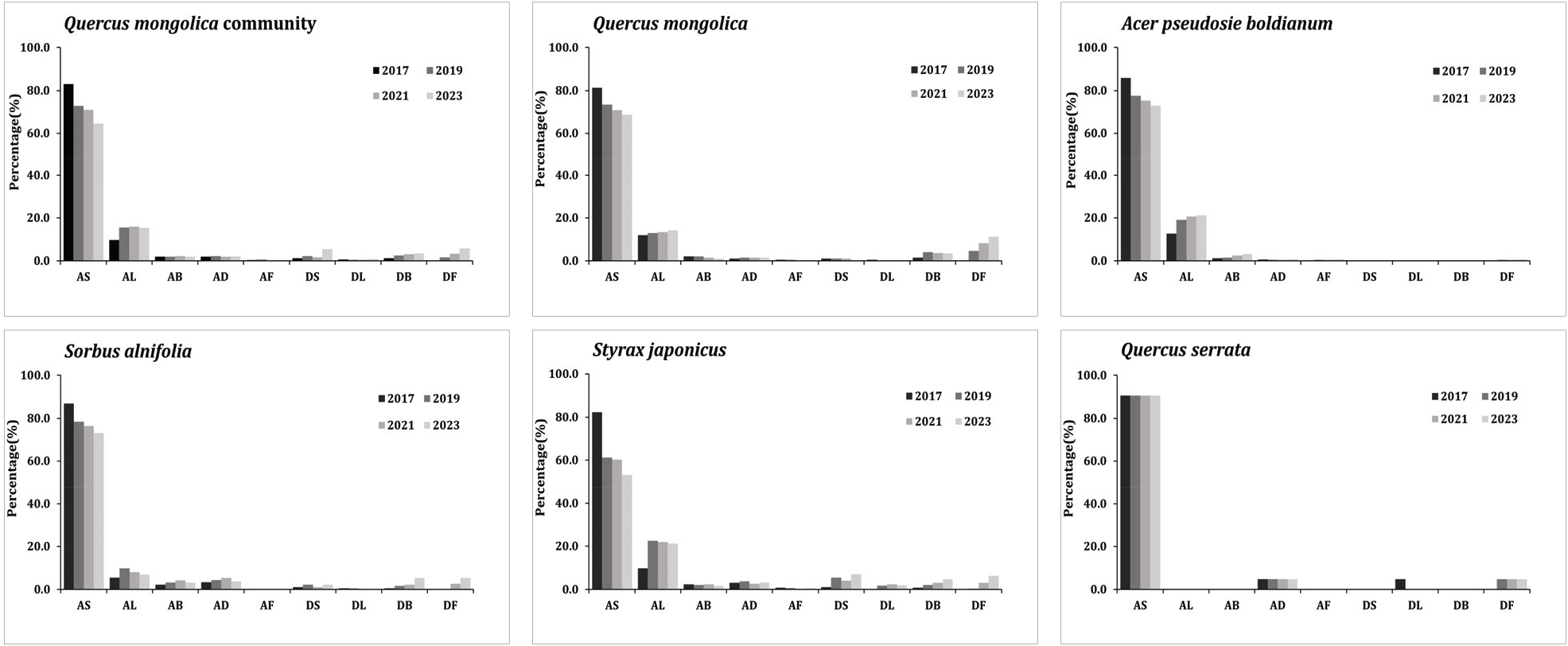
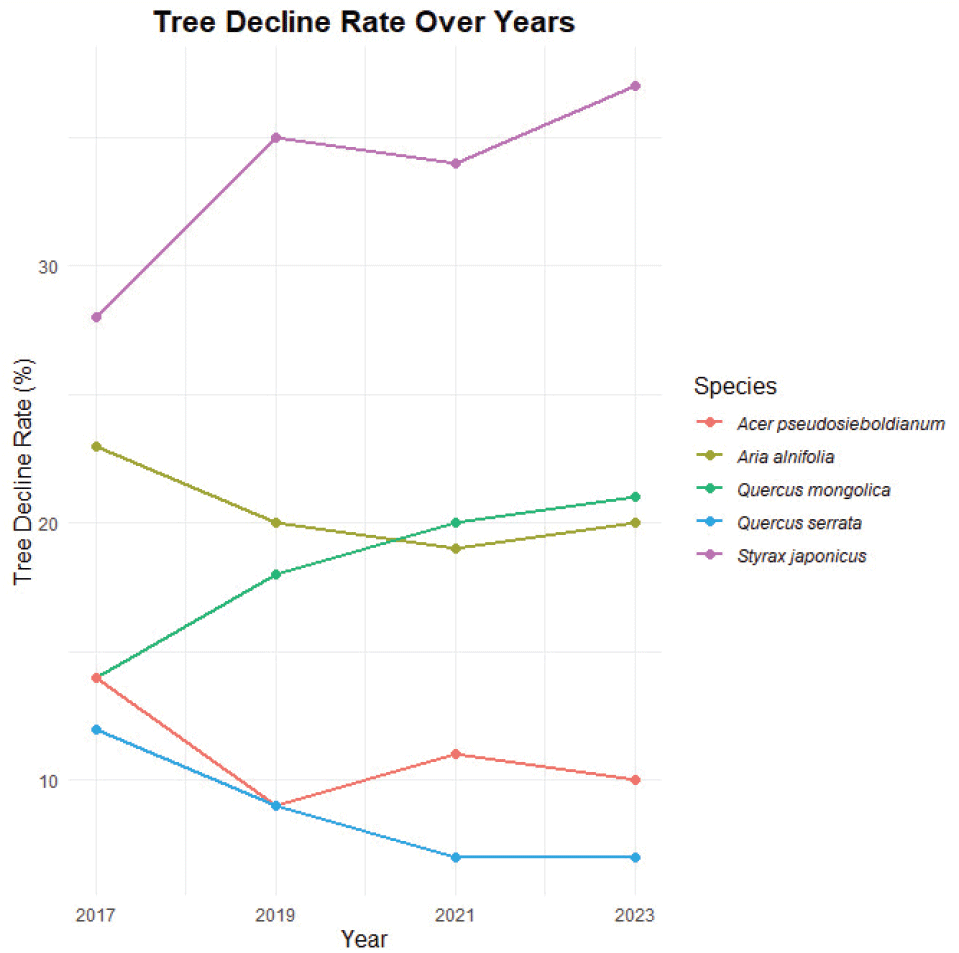
5. 수간건강상태 및 임목 쇠퇴율 변화
전체 수종의 경우 AS 유형은 2017년 959 stems ha−1(83.0%)에서 2023년 848 stems ha−1(64.6%)로 감소하고 있었으며, AL 유형은 2017년 110 stems ha−1(9.5%)에서 2023년 202 stems ha−1(15.3%)로 증가하는 것으로 나타났다(Figure 10). 전체 수종에서 2017년과 2019년 사이 AS 유형에서 AL 유형으로의 수간건강상태 변화 비율이 다른 연도와 비교해 높은 것으로 나타났는데 수관활력도와 함께 고려하였을 때 해당 개체목의 수관활력도는 같은 시기에 높아진 것으로 나타났다. 이는 개체목이 2017년에서 2018년 사이 엽량이 증가함에 따라 수관의 무게는 증가하게 되었고, 다른 시기에 비해 2018년도에 태풍 발생 빈도가 높아 수간(tree stem)의 기계적 안정성(Mechanical stability)이 떨어짐에 따라 AS 유형에서 AL 유형으로 변화한 개체목 유형이 증가한 것으로 판단되었다(Gardiner and Quine, 2000).
Figure 10.
The biannual change of tree health status of community and major species population(5) in Mt. Namsan.
Download Original Figure
신갈나무는 AS 유형이 2017년 157 stems ha−1(81.3%)에서 2023년 136 stems ha−1(68.7%)로 감소하였으며, AL 유형은 2017년 23 stems ha−1(11.9%)에서 2023년 28 stems ha−1(14.1%)로 증가하였다. DF 유형은 2017년 0 stems ha−1(0.0%)에서 2023년 22 stems ha−1(11.1%)로 증가하였다. 종합적으로 신갈나무의 AS 유형은 감소, DB, DF 유형은 연도가 지남에 따라 증가함으로서 임목쇠퇴율 또한 증가하였다(Figure 11). 당단풍나무는 AS 유형이 2017년 300 stems ha−1(85.7%)에서 2023년 299 stems ha−1(72.9%)로 유지되는 경향을 보였으며, AL 유형은 2017년 44 stems ha−1(12.6%)에서 2023년 87 stems ha−1(21.2%)로 증가하였다. 그 외 때죽나무와 팥배나무 모두 2017년에서 2019년 사이 AS 유형에서 AL 유형으로의 변화가 크게 나타났다. 졸참나무의 경우 교목성 수종으로서 조사구 내 평균 DBH가 40 cm이기 때문에 다른 수종과 달리 수간건강상태에 있어 자연적 영향을 덜 받은 것으로 판단되었다.
비교적 신갈나무림 내에서 수간건강상태와 수관활력도가 6년간 지속적으로 좋은 당단풍나무 개체군과 팥배나무 개체군, 졸참나무 개체군은 다른 수종에 비하여 임목쇠퇴율이 낮게 나타났으며, 신갈나무 개체군과 때죽나무 개체군은 시간이 지남에 따라 AS 유형이 감소하고 DF 유형이 증가함에 따라 임목쇠퇴율이 증가하는 추세로 나타났다.
종합고찰
도시림의 생태적 관리와 유지 방안의 생태학적 근거 자료를 확보하고 이를 활용하고자 서울시 남산의 신갈나무림을 대상으로 2017년부터 2023년까지 신갈나무군집과 그 군집을 구성하는 개체군의 변화 과정을 밝히고자 하였다(Figure 12).
연구 결과, 개체군 별 밀도는 당단풍나무, 때죽나무, 신갈나무, 팥배나무 순으로 높게 구성되어 있었으며, 교목층 우점종인 신갈나무 개체군의 밀도는 다소 감소하는 추세였지만 이는 2019년 신갈나무림 내 신갈나무의 강풍에 의한 고사 및 벌채로 인한 밀도 감소인 것으로 판단되며, 흉고단면적은 2019년 이후 다시 증가하고 있었다. 직경급 분포에서 개체목의 대부분이 20-30 cm 직경급이고, 신갈나무림 내에서 중요치가 가장 높은 개체군으로 나타난 점에서 다른 교목성 수종으로의 천이는 어려울 것으로 판단되었다. 아교목층에서는 주로 팥배나무와 당단풍나무가 우점종으로 구성되고 있었으며, 당단풍나무의 경우 밀도, 흉고단면적, 중요치, 수간건강상태 등의 결과에서 세력이 왕성한 것으로 나타나 도시림에서의 당단풍나무의 입지가 지속적으로 커져갈 것으로 판단되었다. 그러나 팥배나무 개체군은 이입률은 감소하고 고사율은 증가하는 추세로 밀도가 점점 감소하고 있다는 점에서 세력이 약화되고 있는 것으로 판단되었다. 때죽나무 개체군은 이입률과 고사율의 비율이 크지 않아 6년간의 밀도 변화가 크지는 않았으나 고사율이 다른 주요 개체군에 비해 높다는 점에서 관목층의 당단풍나무와의 종간경쟁에 의한 고사가 원인이었을 것으로 판단되며 때죽나무 개체군은 신갈나무림 내 주요 수종 중 가장 임목쇠퇴율이 높기 때문에 추후 지속적인 연구를 통해 때죽나무 고사 원인 규명을 좀 더 명확히 밝힐 필요성이 있다. 졸참나무 개체군은 밀도 변화 없이 직경 생장으로 인한 흉고단면적이 증가하는 등의 형태로 생육이 양호한 것으로 판단되었다. 앞으로도 장기생태연구를 통해 여러 가지 환경 인자가 상호작용하여 일어나는 복잡한 현상들을 해석하며, 남산 신갈나무림의 자연성이 약화되지 않도록 관리되어야 할 것이다(Son et al., 1997). 본 연구 조사구의 경우 남산 생태경관보전지역에 해당되는 만큼 하층식생정리 및 생육이 저조한 개체목이 벌채되는 등 인위적 숲 가꾸기 작업이 이루어지고 있는 것으로 판단되었다. 도시림 속의 신갈나무림 1 ha의 크기를 연구하고 조사함에 있어 그 가치가 크기 때문에 자연성이 높은 신갈나무림 조사구만큼은 인위적 개입이 최소화되어야 할 것으로 판단되었다.
감사의 글
본 연구는 국립생태원 연구과제 “국가장기생태연구-남산, 지리산 식생동태 파악을 위한 매목조사”(과제번호: NIE-B-2024-02)의 지원으로 수행되었기에 감사드립니다.
References
Arnone III, J.A. and Gordon, J.C. 1990. Effect of nodulation, nitrogen fixation and CO2 enrichment on the physiology, growth and dry mass allocation of seedlings of
Alnus rubra Bong. New Phytologist 116: 55-66.


Austin, M.P. 1976. On non-linear species response models in ordination. Vegetatio 33: 33-41.
Barbour, M.G., Burk, J.H. and Pitts, W.D. 1987. Terrestrial Plant Ecology(2nd Edition). The Benjamin/Cummings Publishing Co, Menlo Park, USA. pp. 634.
Byeon, S.Y. and Yun, C.W. 2018. Comparison of vegetation structure between natural forest(
Pinus densiflora forest and
Quercus mongolica forest) and
Larix kaempferi forest in Mt. Janggunbong. Journal of Agriculture & Life Science 52(4): 31-45.
Cheon, K.I., Byun, J.G., Jung, S.C. and Sung, J.H. 2014. Community structure of
Quercus mongolica stand in Hyangrobong area, Baekdudaegan. Journal of Agriculture & Life Science 48(1): 1-13.
Cho, H.J., Lee, J.H. and Bae, K.H. 2006. Composition and structure of the major forest stands in Mt. Worak, Korea: on the national long-tem ecological research sites. Journal of Ecology and Field Biology 29(6): 531-537
Cho, Y.C., Lee, C.S., Cho, H.J., Lee, K.S. and Park, P.S. 2011. Vegetation change and emerging research feedback for Korean National Long Term Ecological Research (KNLTER). Journal of Ecology and Environment 34(1), 87-93.
Cho, Y.C., Sim, H.S., Jung, S.H., Kim, H.G., Kim, J.S. and Bae, K.H. 2020. Mid-term (2009-2019) demographic dynamics of young beech forest in Albongbunji Basin, Ulleungdo, South Korea. Journal of Ecology and Environment 44: 27
Choi, D.S. and Toda, H.R.T. 2012. Physiological responses and growth performance of
Alnus firma and
Alnus hirsuta seedlings to artificial soil acidification. Journal of the Korea Society of Environmental Restoration Technology 15(5): 145-153.
Clark, J.S. 1986. Coastal forest tree population in a changing environment, southeastern long Island, New York. Ecological Monographs 56: 259-277.
Condit, R. 1998. Tropical Forest Census Plots. Berlin: Springer-Verlag & R.G. Landes Company pp. 211.
Curtis, J.T. and McIntosh, R.P. 1950. The interrelations of certain analytic and synthetic phytosociological characters. Ecology 31(3): 434-455.
Curtis, J.T. and McIntosh, R.P. 1951. An upland forest continuum in the prairie-forest border region of Wisconsin. Ecology 32(3): 476-496.
Friedman, M. 1937. The use of ranks to avoid the assumption of normality implicit in the analysis of variance. Journal of the American Statistical Association, 32: 675-701.
Gardiner, B.A. and Quine, C.P. 2000. The Mechanical Adaptation of Ttrees to Environmental Influences. Forestry Commission Research Agency. pp. 1-30.
Han, B.H., Park, S.C., Kim, J.Y. and Kwak, J.I. 2022. Ecological characteristics and changes of
Quercus mongolica community in Namsan (Mt.), Seoul. Journal of the Korean Institute of Landscape Architecture 50(2): 41-63.
Huss-danell, K. and Sellstedt, A. 1985. Nitrogenase activity in response to darkening and defoliation of
Alnus incana. Journal of Experimental Botany 36(9): 1352-1358.
Jee, Y.K. and Oh, K.K. 2001. Change of vegetation structure for 6 years (1994-1999) at the harvested forest area (I) - A case of (Mt.) Baegwoonsan research forest at Kwangyang city -. Journal of Korean Society of Forest Science 90(6): 673-682.
Jung, S.C., Koo, K.S. and Kim, K.H. 2011. Inhibitory factors of
Robinia pseudoacacia distribution in a
Pinus thunbergii forest at the coast. Korean, Seoul. Journal of Environment and Ecology 25(5): 717-724.
Jung, S.H., Cho, Y.C. and Lee, C.S. 2021. Changes in leaf and reproductive traits of mountain ash (
Sorbus alnifolia) as urban flourisher in the Seoul Metropolitan, South Korea. Journal of Environment and Ecology 35(6): 644-658.
Kim et al. 2023. Environmental Ecology – Conservation and Management of Ecosystem(2nd Edition). Life Science. Seoul. pp. 304.
Kim, D.H. 2017. A study on the composition characteristics of
Q. mongolica communities in the Baekdudaegan mountains. Ph. D. Dissertation, Seoul. University of Dongguk.
Kim, G.S., Son, H.K., Lee, C.H., Cho, H.J., and Lee, C.S. 2011. Ecological comparison of Mongolian oak (
Quercus mongolica Fisch. ex Ledeb.) community between Mt. Nam and Mt. Jeombong as a Long Term Ecological Research (LTER) site. Journal of Ecology and Environment 34(1): 75-85.
Kim, H.C. 2016. Population structure and dynamics of the main stand in protected area for forest genetic resource conservation of Sokwang-ri. Master’s Dissertation, Daegu. Kyungpook National University.
Kim, J.U. and Kil. B.S. 2000.
Quercus mongolica Forest in Korea. Wonkwang University Press, Iksan. pp. 511.
Kim, M.S., Cho, H.J., Kim, J.S., Bae, K.H. and Chun, J.H. 2018. The classification of forest vegetation types and species composition in the sector between danmoknyeong and guryongnyeong of Baekdudaegan. Korean Journal of Environment and Ecology 32(2): 176-184.
Kim, M.S., Yun, S.J., Park, C.W., Choi, W.I., Chun, J.H., Lim, J.H. and Bae, K.H. 2021. Sequential changes in understory vegetation community for 15 years in the long-term ecological research site in central temperate broad-leaved deciduous forest of Korea. Korean Journal of Environment and Ecology 35(3): 223-236.
Ko, S.Y., Han, S.H. and Yun, C.W. 2013. Population structure and dynamics of the
Picea jezoensis stand in Mt. Gyebangsan. Journal of Korean Society of Forest Science 102(3): 355-364.
Kwak, J.I. 2011. A Study on vegetation structure characteristics and ecological succession trends of Seoul urban forest, Korea. Ph. D. Dissertation, Seoul. University of Seoul.
Lambers, H., Chapin III, F.S. and Pons, T.L. 2008. Plant physiological ecology. 2rd Ed. Springer. New York, U.S.A. pp. 604.
Lee, C.B. 2003. Coloured Flora of Korea. Hyangmunsa. Seoul, Korea. pp. 1828.
Lee, C.S., Cho, Y.C., Shin, H.C., Lee, C.H., Lee, S.M., Seol, E.S., Oh, W.S. and Park, S.A. 2006. Ecological Characteristics of Korean Red Pine (
Pinus densiflora S. et Z.) Forest on Mt. Nam as a Long Term Ecological Research (LTER) Site. Journal of Ecology and Environment 29(3): 593-602.
Lee, H.J., Lee J.S. and Byun, D.W. 1994. Community classification and vegetation pattern of
Quercus mongolica forest in Mt. Myongji. Korean Journal of Environment and Ecology 17(2): 185-201.
Lee, J.E. 2023. Community structure and sequential change of
Abies Koreana forest in Mt. Hallasan and Mt. Jirisan. (Dissertation). Kongju. Kongju National University.
Lee, J.H., Cho, H.J., Lee, B.C., Oh, S.H. and Bae, K.H. 2007. Forest vegetation types and growth characteristics of Seonginbong in Ulleung island, Korea. Journal of Agricultural and Forest Meteorology 9(1): 37-48.
Lee, K.J., Ki, K.S., Han, B.H. and Kim, J.H. 2006. Change for eleven years (1994~2005) of plant community structure of major community in Namsan, Seoul. Korean Journal of Environment and Ecology 20(4): 448-463.
Lee, M.J. and Song, H.K. 2011. Vegetation structure and ecological restoration model of
Quercus mongolica community. Journal of the Korean Society of Environmental Restoration Technology 14(1): 57-65.
Lee, W.C. and Yim, Y.J. 1978. Studies on the distribution of vascular plants in the Korean peninsula. Korean Journal of Plant Taxonomy 8: 1-33.
Lim et al. 2019. The Current status and conservation measures of endangered high-altitude coniferous forests in Korea.
https://book.nifos.go.kr. (2019. 06.).
Lim, J.S. Moon, G.H., Lee, M.W., Kang, J.T., Won, M.S., An E.S., Jeon J.H. 2021. 2020 Korea Forest Resources.
https://book.nifos.go.kr. (2022.6.)
NIBR (National Institute of Biological Resources). 2023. National list of Korea.
https://kbr.go.kr/. (2024. 09.30)
No, T.H. 2014. The Eco-adaptive urban forest management methods of Namsan(Mt.) according to the urban environment change and management in Seoul, Korea. Ph. D. Dissertation, Seoul, University of Seoul.
Park, B.J. 2019. Characteristic for ecological community and stand dynamics of forest vegetation in Mt. Myeonbong. Ph. D. Dissertation, Daegu. University of Kyungpook.
Park, D.E., Lee, J.E., Park, G.E., Yang, H.M., Kim, H.J. and Yun, C.W. 2024. Vegetation change of
Abies koreana habitats in the subalpine zone of Mt. Jirisan over eight years. Journal of Korean Society of Forest Science 113(2): 222-238.
Park, G.E., Kim, E.S., Jung, S.C., Yun, C.W., Kim, J.S., Kim, J.D., Kim, J.B. and Lim, J.H. 2022. Distribution and stand dynamics of subalpine conifer species (
Abies nephrolepis,
A. koreana, and
Picea jezoensis) in Baekdudaegan protected area. Journal of Korean Society of Forest Science 111(1): 61-71.
Park, H.K. and Yun, C.W. 2009. A Study on forest vegetation classification in urban forest of Daejeon metropolitan city. The Journal of Korean Institute of Forest Recreation 13(4): 33-41.
Park, M.S. 2024. Study on vegetation changes following the management of oak wilt disease in
Quercus mongolica forest of Namsan(Mt.), Seoul. Master’s Dissertation, Seoul. University of Seoul.
Park, S.T. 2004. New Mountain and Stream Diagram. Chosun Ilbo. Seoul. pp. 532
Rédei, K., Osváth-Bujtás, Z. and Balla, I. 2002. Clonal approaches to growing black locust (
Robinia pseudoacacia) in Hungary: a review. Forestry 75: 547-552.
Seoul. 1992. Mt. Nam. Ecological Landscape Conservation Area Management Plan. Seoul. pp. 239. (in Korean)
Seoul. 2008. Study on Detailed Changes in Mt. Nam. Ecological Landscape Conservation Area. Seoul. pp. 132. (in Korean)
Seoul. 2023. Study on Detailed Changes in Mt. Nam. Ecological Landscape Conservation Area. Seoul. pp. 306. (in Korean)
Sheil, D. 1995. A critique of permanent plot methods and analysis with examples from Budongo forest, Uganda. Forest Ecology and Management 77: 11-34.
Shin, J.H. and Kim, C.M. 1996. Ecosystem classification of Korea (Ⅰ): Division of ecological zones. Journal of Forest Science, 54: 188-199.
Smith D.M. and Hawley, R.C. 1986. The practice of silviculture. 6rd Ed. John Wiley and Sons Inc. New York, U.S.A. pp. 514.
Son, S.Y., Kwon, K.C. and Jeong, T.S. 2002. Productive structure and net production of
Quercus mongolica forest in Mt. Taehwa (Kwangju, Kyonggi-do). Journal of Korea Forestry Energy 21(1): 76-82.
Son, Y.H. 1997. Long-term ecology research: importance and approaches. Natural Resources Research 5: 29-41.
Son, Y.H., Koo, C.D. Kim, C.S., Park, P.S., Yun, C.W. and Lee, K.H. 2016. Forest Ecology. Hyangmunsa. Seoul. pp. 346.
Son, Y.H., Koo, C.D. Kim, C.S., Park, P.S., Yun, C.W. and Lee, K.H. 2024. Samgo Forest Ecology. Hyangmunsa. Seoul. pp. 366.
Song, H.G., Lee, M.J., Lee, S., Kim, H.J., Ji, Y.W. and Kwon, O.W. 2003. Vegetation structures and ecological niche of
Quercus mongolica Forests. Journal of Korean Society of Forest Science 92(4): 409-420.
Walter, H., Harnickell, E., Müller-Dombois, D. 1975. Climate-Diagram Maps of the Individual Continents and the Ecological Climatic Regions of the Earth. Berlin. Springer-Verlag. pp. 36.
Woo, S.Y., Kim, D.G. and Lee, D.S. 2000. Effects of air pollution on physiological characteristics of styrax japonica in Yeolchon industrial complex. Journal of Korean Society for Atmospheric Environment 16(2): 121-128.
Yim Y.J. and Kira, T. 1976. Distribution of forest vegetation and climate in the Korean peninsula: Ⅱ. Distribution of climate humidity/ aridity. Japanse Journal of Ecology 26(3): 157-164.
Yun, C.W. 2022. Field Guide to Trees and Shrubs. Geobook. Korea. pp. 704.
Yun, I.S., Song, J.H., Byeon, S.Y., Kim, H.J., Lee, J.E., Kim, J.D. and Yun, C.W. 2022. Community structure comparison of fagaceae forest vegetation in Namsan, Odaesan, and Ulleungdo. Journal of Korean Society of Forest Science 111(4): 511-529.
Yun, S.W., Kim, H.J., Lee, B.C., Shin, J.H., Yang H.M. and Lim, J.H. 2011. Characteristic community type classification of forest vegetation in South Korea. Journal of Korean Society of Forest Science 100(3): 504-521.