Article
포천지역 초기 조림지 탄소저장량 추정을 위한 소나무 조림목 바이오매스 상대생장식 개발
송규홍
1, 장지원
1, 이세희
1, 이상태
2, 정상훈
2, 노남진
1,3,*
Development of Biomass Allometric Equations for Pinus densiflora Saplings to Estimate Carbon Storage of Early-Stage Plantations in Pocheon Region
Kyu Hong Song
1, Ji Won Jang
1, Se Hee Lee
1, Sang Tae Lee
2, Sang Hoon Chung
2, Nam Jin Noh
1,3,*
1Department of Forestry and Environmental Systems, Kangwon National University, Chuncheon 24341, Korea
2Forest Technology and Management Research Center, National Institute of Forest Science, Pocheon 11187, Korea
3Department of Forest Resources, Kangwon National University, Chuncheon 24341, Korea
© Copyright 2025 Korean Society of Forest Science. This is an Open-Access article distributed under the terms of the
Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits
unrestricted non-commercial use, distribution, and reproduction in any
medium, provided the original work is properly cited.
Received: Mar 28, 2025; Revised: Jun 27, 2025; Accepted: Jul 19, 2025
Published Online: Sep 30, 2025
요 약
본 연구는 소나무(Pinus densiflora) 초기 조림목의 바이오매스 상대생장식을 개발하고 초기 조림지의 탄소 저장량을 추정하기 위해 수행되었다. 2024년 4월, 7년생 소나무 조림지 내에 정방형 조사구 9개를 설치하고 조사구 내 조림목들을 대상으로 매목조사를 실시하였다. 10월에 소나무 조림목 16본을 선정하여 근원경, 흉고직경, 수고, 수관폭을 재측정한 뒤, 뿌리를 포함하여 수확하였다. 소나무 조림목의 바이오매스 추정 모델의 적합성은 근원경 기반(R2>0.96), 수관폭 기반(R2>0.91), 흉고직경 기반(R2>0.88) 모델 순으로 높게 나타났다. 소나무 조림목의 부위별 바이오매스 구성비는 줄기가 17.5%, 가지 28.5%, 잎 36.4%, 그리고 뿌리 17.6%로, 성숙목에 비해 가지와 잎이 높은 비율을 차지했다. 본 연구에서 개발된 바이오매스 상대생장식과 매목조사를 바탕으로 추정된 소나무 초기 조림지의 탄소저장량은 5.25±0.42 Mg C ha−1로 나타났다. 본 연구에서 개발된 소나무 어린나무의 바이오매스 상대생장식은 소나무 재조림지와 산림 복구지의 초기 탄소 저장량과 흡수량을 보다 정확히 추정하는 데 기여하며, 이를 바탕으로 입지 생산성을 정량적으로 평가하는 데 중요한 자료를 제공한다.
Abstract
This study aimed to develop biomass allometric equations for Pinus densiflora saplings to assess carbon storage in young plantations. In April 2024, nine sample plots were established in a 7-year-old pine stand, and a tree census was carried out within each plot. In October, 16 saplings were selected for destructive sampling. Prior to harvesting, measurements of root collar diameter (RCD), diameter at breast height (DBH), height, and crown width (CW) were repeated for each sapling, and the entire tree, including its root system, was collected. Among the developed models for biomass estimation, the RCD-based model demonstrated the highest explanatory power (R2 > 0.96), followed by the CW-based model (R2 > 0.91) and the DBH-based model (R2 > 0.88). The biomass allocation by component was 17.5% in stems, 28.5% in branches, 36.4% in foliage, and 17.6% in roots, with branches and foliage accounting for a higher proportion compared to mature trees. Using the biomass equations and census data, carbon storage in early-stage plantations was estimated at 5.25 ± 0.42 Mg C ha−1. These equations support more accurate estimations of carbon storage and sequestration in pine reforestation and restoration efforts, offering critical data for evaluating forest productivity.
Keywords: biomass; carbon uptake; pine; root collar diameter; sapling
서 론
우리나라는 산림 탄소흡수 기능 증진, 영급 불균형 개선, 수종 갱신, 산불피해지 복원 등을 위해 재조림 면적이 점차 확대되고 있다(KFS, 2024). 재조림은 단순히 목재 공급을 넘어, 산림부문 탄소중립 이행을 위한 핵심 전략으로 자리 잡고 있다(KFRI, 2016). 산림은 대기 중 이산화탄소를 흡수하고 저장하며, 이를 통해 기후변화 완화에 중요한 역할을 한다(Hasler et al., 2024). 그러나 벌채 이후 재조림된 산림의 초기 탄소저장량 및 흡수량에 대한 정보는 여전히 매우 부족하며, 이는 산림의 생산성과 탄소저장 잠재력을 효과적으로 평가하는 데 장애가 되고 있다(Chan et al., 2018). 따라서, 초기 조림지에서의 생산성과 탄소 저장량을 정확히 추정하고 지속적인 모니터링이 필요한 시점이다(Song et al., 2025).
바이오매스 상대생장식은 산림의 생산성 및 탄소저장량을 추정하기 위해 가장 널리 사용되고 있으며, 일반적으로 흉고직경(Diameter at breast height, DBH)이나 수고와 같은 현장에서 측정할 수 있는 생장 지표를 매개변수로 활용하여 개발한다(Pati et al., 2022). 국내에서는 2000년 초부터 산림 부문 국가 온실가스 인벤토리 구축을 위해 바이오매스 상대생장식 및 탄소배출계수가 개발된 바 있다(KFRI, 2014). 그러나 기존의 바이오매스 상대생장식은 성숙림에서 생육하는 개체들을 대상으로 개발되었기 때문에, 초기 조림목의 형태적 특성을 충분히 반영하지 못한다. 또한 초기 조림목은 성숙목에 비해 수고 및 부피생장이 미발달된 상태이므로, 흉고직경보다 근원경(Root collar diameter, RCD)을 이용한 바이오매스 상대생장식이 더 높은 적합성을 갖는다(Blujdea et al., 2012, Thippawan et al., 2023; Song et al., 2025). 한편, 최근에는 수관폭(Crown width, CW)을 이용한 바이오매스 상대생장식 개발 연구 사례가 증가하고 있으며, 다양한 수종과 생육 환경에서 그 적용 가능성이 보고되고 있다(Mclntire et al., 2022; Menéndez-Miguélez et al., 2022; Juan-Ovejero et al., 2023; Madalcho et al., 2023).
초기 조림목의 생장 특성은 성숙림에서 생육하는 치수와 뚜렷한 차이를 보인다. 조림지에서 생육하는 초기 조림목은 개벌 이후 인위적으로 식재되어 광 자원이 상대적으로 풍부한 환경에서 생육되며, 이로 인해 잎과 가지 등 수관 부위로의 바이오매스 할당 비율이 높아지는 경향이 있다(Chave et al., 2005; Nogueira et al., 2014). 반면 성숙림 내에서 자연적으로 발생하는 치수는 우세목들에 의해 광이 제한된 환경에서 생육하기 때문에 이러한 할당 특성이 상대적으로 미약하게 나타난다. 또한 초기 조림목은 바이오매스 할당에 있어 풀베기, 어린나무 가꾸기와 같은 조림 초기 단계에서 실시되는 산림 관리 작업의 직접적인 영향을 받지만, 성숙림 내의 치수는 이러한 관리의 영향을 받지 않는 경우가 대부분이다. 이러한 임분의 구조적⋅생태적 특성 차이는 개체목의 부위별 바이오매스 할당 방식 및 수준에 영향을 미치며, 이를 고려하지 않은 바이오매스 추정은 초기 조림지의 탄소저장량 및 탄소흡수량 추정의 불확실성을 초래할 수 있다(Bjarnadóttir et al., 2007; Menéndez-Miguélez et al., 2022). 또한 수종에 따라 생장 특성 및 조림지에 대한 환경적응 능력이 상이하기 때문에 주요 조림 수종을 대상으로 한 초기 조림목 바이오매스 상대생장식이 필요하다(Fradette et al., 2021; Song et al., 2025).
소나무(Pinus densiflora)는 일본잎갈나무, 편백 등과 함께 한국의 경제림 조성을 위한 주요 조림 수종으로, 2020년 기준, 혼효림을 제외한 침엽수림과 활엽수림 내에서 가장 넓은 산림 면적(36.6%)과 가장 높은 입목축적(43.8%) 비율을 차지하고 있다(KFS, 2024). 하지만 재선충병, 대형 산불 등 소나무 수종에 대한 피해가 커지면서 조림 비율 및 묘목 생산은 감소하는 추세이나, 2023년 기준 소나무의 조림본수(7,661,000본)는 전체 조림 실적의 3번째(16%)로 여전히 많은 비중을 차지하고 있다(KFS, 2024). 따라서 본 연구는 흉고직경 6 cm 이하의 포천지역 소나무 초기 조림목을 대상으로 부위별 바이오매스 상대생장식을 개발하고 초기 조림지의 탄소저장량 추정을 위해 수행되었다.
재료 및 방법
1. 연구대상지
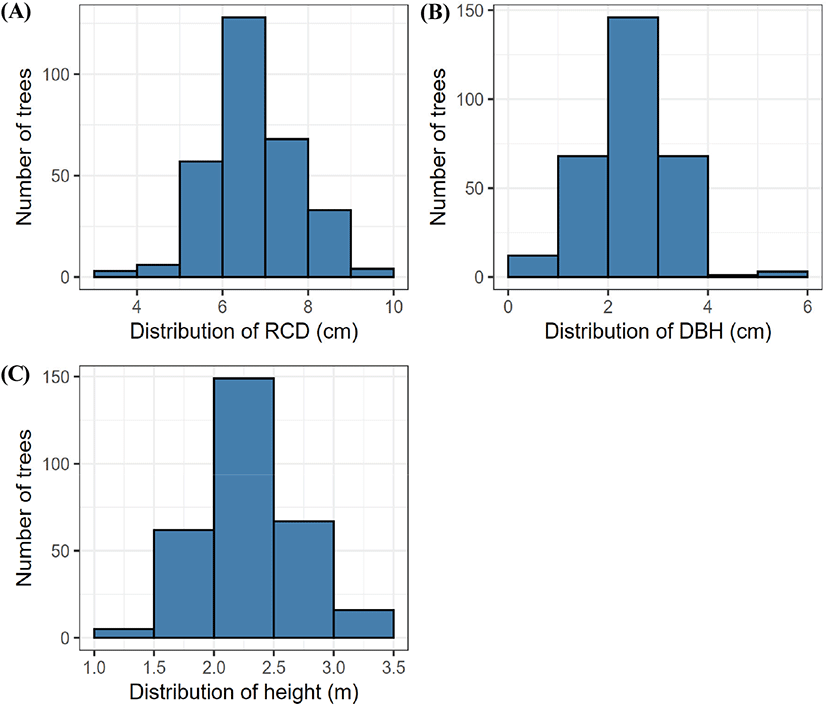
본 연구는 경기도 포천시 소흘읍 산림기술경영연구소에 위치한 7년생 소나무 조림지(37°46’03” N, 127°10’38” E, 180 m a.s.l.)를 대상으로 수행되었다. 2024년 4월, 소나무 조림지 내 10 m×10 m 크기의 정방형 조사구 9개를 설치하고 조사구 안에 있는 조림목들의 근원경, 흉고직경, 수고를 측정하였다(Figure 1). 매목조사 결과, 임분밀도는 2,767±66 trees ha−1이었으며, 조림목의 평균 근원경, 흉고직경, 수고는 각각 6.62±0.12 cm, 2.26±0.09 cm, 2.17±0.04 m로 나타났다. 연구대상지의 평균 경사는 20°, 사면 방위는 북서사면(NW 318°)이었으며 2020년에서 2024년 동안 연평균 기온과 강수량은 각각 11.2°C와 1442.6 mm로 나타났다(Korea Meteorological Administration, 2024). 토양 용적밀도는 약 1.11±0.05 g cm−3이었으며, 토성은 모래 41.6%, 미사 34.0%, 점토 24.4% 함유되어 있는 양토로 나타났다(PARIO, METER Group, Germany). 토양 표층의 전질소 및 총탄소 농도는 각각 0.12±0.01%, 2.26±0.17%로 나타났으며(Vario Max Cube, Elementar, Germany), 토양 pH는 4.50±0.04로 나타났다(Lab 855 pH Meter, SI Analytics, Germany).
Figure 1.
Distributions of (A) root collar diameter (RCD), (B) diameter at breast height (DBH), (C) height of P. densiflora saplings measured within the study plots in April 2024. The histograms show the number of trees grouped by size class in each growth variable.
Download Original Figure
2. 바이오매스 상대생장식 및 확장계수 개발
2024년 10월, 조림목의 근원경 및 흉고직경 분포를 고려하여 총 16본을 선정하였다. 일반적으로 바이오매스 상대생장식을 개발할 경우 최소 30본 이상의 표본이 요구되나, 어린나무의 경우에는 최소 10본 이상의 표본으로도 모델의 높은 설명력과 유의성을 확보할 수 있음이 보고된 바 있다(Roxburgh et al., 2015; Han and Park, 2020; Vorster et al., 2020). 선정된 개체목들을 대상으로 근원경, 흉고직경, 수고, 수관폭을 재측정한 후, 지하부 뿌리까지 굴취하여 수확하였다. 수확한 소나무 조림목의 부위별 바이오매스 측정을 위해 줄기, 가지, 잎, 뿌리를 분리하여 65°C 오븐에 넣고 항량에 도달할 때까지 건조시킨 후 건중량을 측정하였다.
바이오매스 상대생장식은 부위별 건중량과 근원경, 흉고직경, 수고, 수관폭을 이용하여 개발되었다(Pajtík et al., 2008; Zianis et al., 2011; Cutini et al., 2013; Mclntire et al., 2022) (식 1–3).
(Y=부위별 바이오매스, X=흉고직경 또는 근원경, H=수고, Z=수관폭)
이후, 개발된 바이오매스 상대생장식의 추정 회귀선과 우리나라 중부지방 소나무 성숙림에서 개발된 바이오매스 상대생장식(Noh et al., 2013)의 추정 회귀선을 부위별로 비교하였다.
줄기 밀도는 수확한 소나무에서 약 1 m 간격으로 10 cm 두께의 원판을 채취한 후, 각 원판의 원구와 말구 직경을 측정하고 Smalian 식을 적용하여 재적을 산출하였다(식 4).
(V=원판 재적, L=원판 길이, B=원구 단면적, S=말구 단면적)
이후, 원판 시료를 65°C에서 오븐 건조하여 건중량을 측정하였으며, 이를 이용해 줄기 재적 대비 건중량의 비율로 밀도를 계산하였다. 바이오매스 확장계수(Biomass Expansion Factors, BEF)는 줄기 건중량 대비 지상부 건중량의 비율로 계산되었다.
3. 조림지 입목 탄소저장량 추정
매목조사에 의한 근원경과 흉고직경 자료를 본 연구에서 개발한 바이오매스 상대생장식과 선행 연구에서 개발된 바이오매스 상대생장식(Noh et al., 2013)에 각각 적용하여 임분 수준의 부위별 바이오매스 현존량을 산정한 후, 탄소전환계수 0.5를 적용하여 탄소저장량을 추정하였다.
4. 통계 분석
본 연구에서는 소나무 조림목의 근원경, 흉고직경, 및 수관폭을 매개변수로 하여 바이오매스 상대생장식을 도출하였으며, 근원경과 흉고직경을 이용한 모델은 비선형회귀분석(Non-linear regression analysis), 수관폭을 이용한 모델은 일반선형회귀분석(Simple linear regression analysis)을 통해 각각 개발되었다. 바이오매스 추정 모델의 적합성 평가는 결정계수(R2)와 평균제곱근오차(Root Mean Square Error, RMSE)를 기준으로 실시하였다.
각 모델의 R2는 독립변수가 종속변수의 변동을 설명하는 정도를 나타내며(식 5), RMSE는 모델의 예측값과 관측값 간의 차이를 정량적으로 평가하는 지표로, 값이 낮을수록 모델의 예측 정확성이 높은 것으로 간주하였다(식 6).
(yi=관측값, =모델 예측값, =종속변수의 평균값)
회귀계수의 통계적 유의성은 p-value를 기준으로 평가하였으며, 유의수준은 p<0.05, p<0.01, p<0.001에서 각각 *, **, *** 기호로 표시하였다. 모든 통계 분석은 R version 4.4.1에서 제공하는 ‘stats’ 패키지를 이용하여 수행되었다.
결과 및 고찰
1. 바이오매스 상대생장식
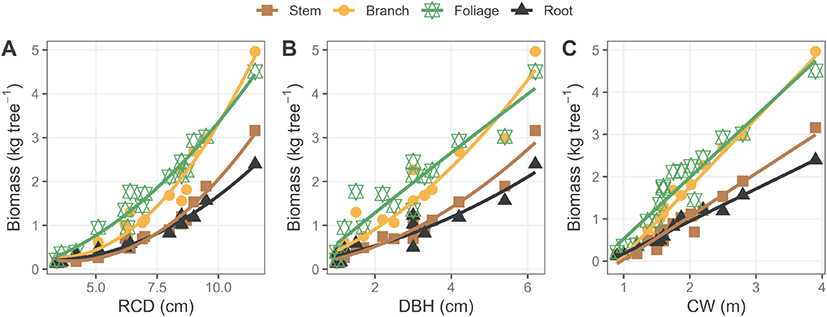
소나무 초기 조림목의 근원경, 흉고직경, 수관폭 및 수고에 대한 부위별 바이오매스 상대생장식의 조제 결과는 Table 1과 같다. 소나무 조림목의 근원경, 흉고직경, 수관폭 증가에 따른 부위별 바이오매스의 증가율은 잎과 가지가 줄기나 뿌리에 비해 높았으며, 이러한 상대적 경향성은 모든 모델에서 유사하게 나타났다(Figure 2). 소나무 초기 조림목의 바이오매스 추정 모델의 적합성은 근원경(R2>0.96, RMSE=0.22), 수관폭(R2>0.91, RMSE=0.31), 흉고직경(R2>0.88, RMSE=0.38) 기반 모델 순으로 높게 나타났다(Table 1, Figure 3). 또한 줄기와 뿌리에서 근원경 또는 흉고직경 하나의 독립변수로 개발한 모델에 비해 수고를 포함한 두 개의 독립변수로 개발한 모델이 비교적 더 높은 설명력을 보였다(Table 1).
Table 1.
Regressions of dry weights (kg) on different components in P. densiflora saplings. Y: dry weights of tree component (kg), RCD: root collar diameter (cm), DBH: diameter at breast height (cm), H: height (m), CW: crown width (m).
|
Model |
Component |
a |
b |
c |
R2
|
RMSE |
|
Y = a × RCDb
|
Stem |
0.0016**
|
3.1010***
|
|
0.9774 |
0.1136 |
|
Branch |
0.0044*
|
2.8775***
|
|
0.9673 |
0.2181 |
|
Foliage |
0.0270**
|
2.0946***
|
|
0.9693 |
0.1924 |
|
Root |
0.0061*
|
2.4351***
|
|
0.9777 |
0.0941 |
|
Total |
0.0342**
|
2.4866***
|
|
0.9863 |
0.4737 |
|
Y = a × RCDb × Hc
|
Stem |
0.0221 |
1.3960**
|
1.1180***
|
0.9907 |
0.0727 |
|
Branch |
0.0113 |
2.2549**
|
0.4142 |
0.9693 |
0.2112 |
|
Foliage |
0.0228 |
2.2111***
|
−0.0847 |
0.9694 |
0.1918 |
|
Root |
0.0166 |
1.7787*
|
0.4443 |
0.9807 |
0.0877 |
|
Total |
0.0748 |
1.9713**
|
0.3471 |
0.9878 |
0.4459 |
|
Y = a × DBHb
|
Stem |
0.1720***
|
1.5360***
|
|
0.9445 |
0.1779 |
|
Branch |
0.3310***
|
1.4250***
|
|
0.9041 |
0.3733 |
|
Foliage |
0.6295***
|
1.0335***
|
|
0.8807 |
0.3790 |
|
Root |
0.2279**
|
1.2251***
|
|
0.8803 |
0.2182 |
|
Total |
1.3400**
|
1.2680***
|
|
0.9101 |
1.2123 |
|
Y = a × DBHb × Hc
|
Stem |
0.1718***
|
0.5225**
|
1.4000***
|
0.9869 |
0.0865 |
|
Branch |
0.3199***
|
0.5184 |
1.2830*
|
0.9419 |
0.2905 |
|
Foliage |
0.5769***
|
0.4725 |
0.8589 |
0.9049 |
0.3384 |
|
Root |
0.2232***
|
0.0937 |
1.5847*
|
0.9462 |
0.1463 |
|
Total |
1.3246***
|
0.3106 |
1.3367*
|
0.9550 |
0.8581 |
|
Y = a + bCW
|
Stem |
−0.9804***
|
1.0187***
|
|
0.9512 |
0.1668 |
|
Branch |
−1.5094***
|
1.6304***
|
|
0.9557 |
0.2539 |
|
Foliage |
−0.9279**
|
1.4555***
|
|
0.9192 |
0.3120 |
|
Root |
−0.5895***
|
0.7665***
|
|
0.9794 |
0.0905 |
|
Total |
−4.0194***
|
4.9261***
|
|
0.9843 |
0.5067 |
Download Excel Table
Figure 2.
Relationships between growth measurement variables (A: root collar diameter, B: diameter at breast height, C: crown width) and tree component biomass (kg tree−1) for P. densiflora saplings. Symbols represent observed values, and lines indicate fitted regression models. Power-law models (Y = a·Xb) were applied for panels A and B, while linear models (Y = a + b·X) were used for panel C.
Download Original Figure
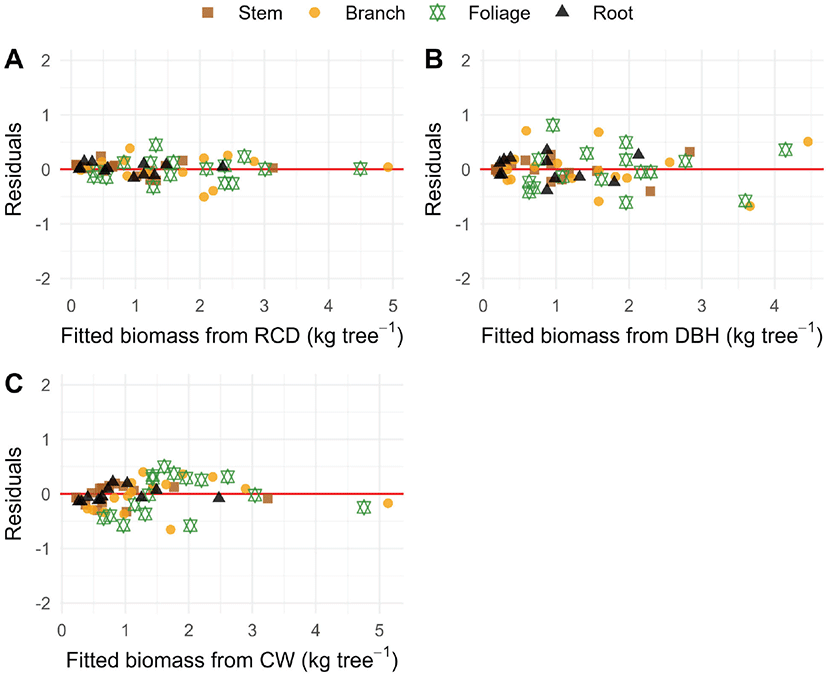
Figure 3.
Residual plots for regression models fitted to the biomass components of P. densiflora saplings, using root collar diameter (RCD) and diameter at breast height (DBH) as predictors in non-linear models (Y = a·Xb) for panels A and B, and crown width (CW) as a predictor in a simple linear regression model (Y = a + b·X) for panel C. Different colors and shapes represent biomass components (stem, branch, foliage, and root). The red horizontal line indicates zero residual.
Download Original Figure
흉고직경 기반 모델의 낮은 적합성은 흉고직경의 측정 높이인 1.2 m에서 충분한 비대생장이 이루어지지 않은 어린나무의 일반적인 특징에 기인하는 것으로 보인다(Thippawan et al., 2023; Song et al., 2025). 또한 수관폭과 바이오매스 간의 높은 상관성은 성숙목을 대상으로 한 연구에서도 보고된 바 있으며, 수관폭이 초기 조림지에서도 바이오매스 추정을 위한 주요 변수로 활용될 수 있는 가능성을 시사한다(Bilgili and Kucuk, 2009; Kim et al., 2009; Goodman et al., 2014; Menéndez-Miguélez et al., 2022; Madalcho et al., 2023). 특히 초기 조림목의 수관폭은 근원경과 수고에 비해 비교적 측정이 용이하고, 원격탐사 자료를 통해서도 추출이 가능하다는 점에서 활용 가능성이 높을 것으로 보인다. 최근, 무인항공기(UAV) 기반의 고해상도 영상을 활용하여 어린나무의 수관과 바이오매스 간의 관계를 분석한 연구 사례들이 보고되고 있어(Mclntire et al., 2022; Juan-Ovejero et al., 2023), 다양한 입지환경으로부터 초기 조림지 내 수관폭 기반 바이오매스 추정 모델의 정확도를 높이는 추가 연구가 필요할 것으로 보인다.
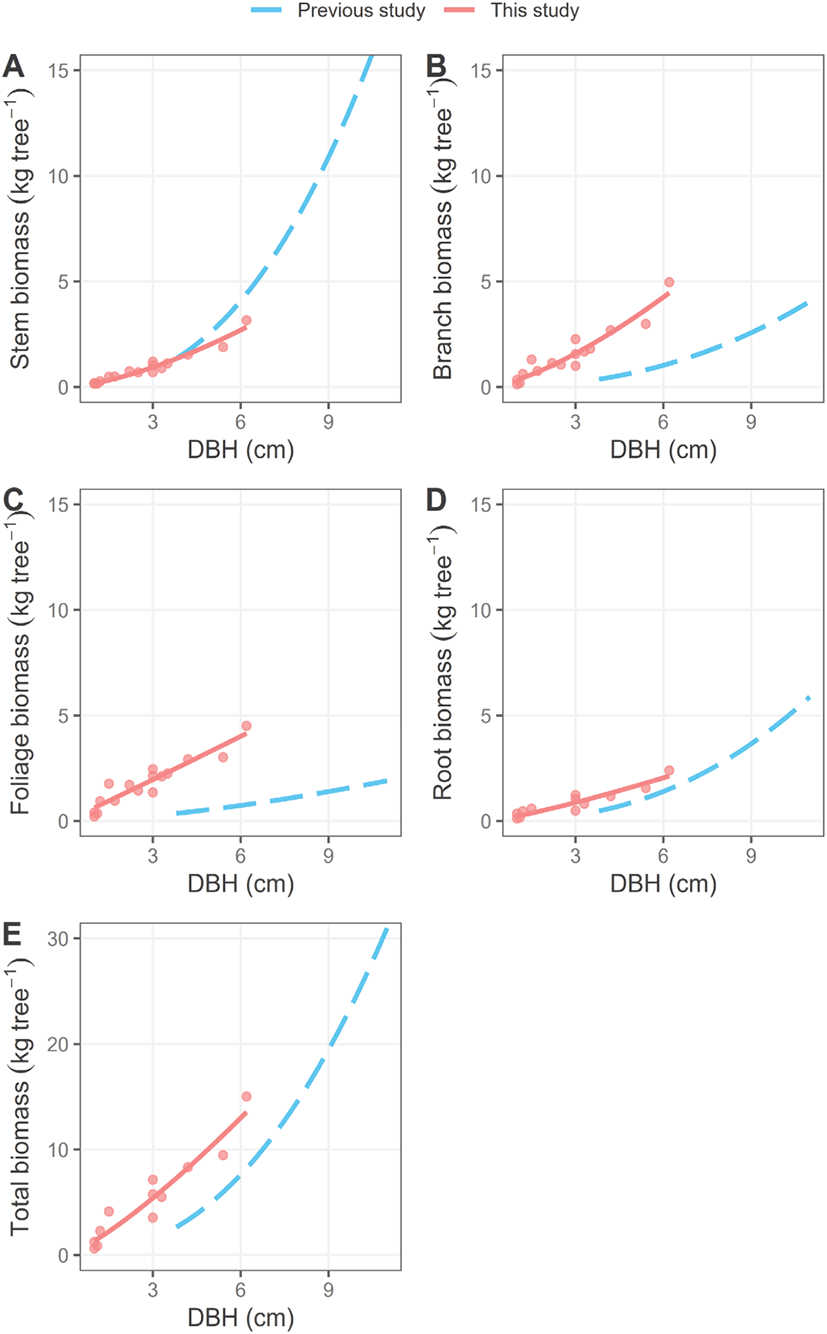
본 연구에서 개발한 소나무 초기 조림목 대상 바이오매스 추정 회귀선과 기존의 성숙임분 내에서 개발된 바이오매스 추정 회귀선을 비교한 결과(Noh et al., 2013), 줄기와 뿌리 바이오매스에서는 두 회귀선 간에 비교적 유사한 경향이 나타났다[Figure 4(A), (D)]. 반면, 잎과 가지 바이오매스의 경우에는 흉고직경 6 cm에서 두 회귀선 간 뚜렷한 차이를 보였다[Figure 4(B), (C)]. 이러한 결과는 동일한 흉고직경일지라도 성숙림과 초기 조림지에서 생육하는 개체 간 탄소 할당 특성이 상이할 수 있기 때문에(Nogueira et al., 2014), 초기 조림목 생육 특성에 기반한 바이오매스 상대생장식 개발의 필요성을 시사한다.
Figure 4.
Comparison of biomass estimation regression lines based on diameter at breast height (DBH) between the previous study (Noh et al., 2013) (blue dashed lines) and this study (red soild lines) for P. densiflora. Red dots indicate observed biomass values from harvested saplings in this study.
Download Original Figure
2. 부위별 바이오매스 구성비 및 확장계수
초기 조림목의 각 부위별 바이오매스가 차지하는 비율을 살펴보면, 잎이 36.37%로 가장 높은 비중을 차지하였으며, 그 뒤로 가지(28.52%), 뿌리(17.57%), 줄기(17.54%) 순으로 분포하였다(Table 2). 본 연구에서 나타난 잎의 바이오매스 비율은 자연 갱신된 10년생 소나무에서 보고된 잎의 비율(13-15%)보다 약 2.6배 높게 나타났다(Noh et al., 2010). 바이오매스 확장계수는 4.85로 나타났으며, 이는 전국 단위로 개발된 중부지방 소나무의 확장계수(1.41)뿐만 아니라(KFRI, 2014), 충북지역 15-16년생 소나무에서 보고된 수치(1.78-2.70)와 비교했을 때도 높은 수준 으로 나타났다(Park et al., 2005). 지상부 바이오매스 대비 뿌리 바이오매스의 비(Root-to-shoot biomass ratio, R:S)는 0.22로, 10년생 소나무 임분의 R:S(0.21)와 유사했지만, 71년생 소나무 임분의 R:S(0.26)에 비해 낮은 값을 보였다(Noh et al., 2010) (Table 2).
Table 2.
Mean biomass (kg tree−1) and biomass distribution ratio (%) of each tree component in P. densiflora saplings, based on harvested samplings. Also presented are the root-to-shoot (R:S) and root-to-total (R:T) biomass ratios, stem density (g cm−3), and biomass expansion factor (BEF).
|
Component |
Biomass (kg tree−1) |
Distribution (%) |
|
Stem |
0.92±0.19 |
17.54±1.11 |
|
Branch |
1.53±0.31 |
28.52±1.14 |
|
Foliage |
1.78±0.28 |
36.37±1.19 |
|
Shoot (S) |
4.23±0.78 |
82.43±1.11 |
|
Root (R) |
0.87±0.19 |
17.57±1.11 |
|
Total (T) |
5.33±1.22 |
100 |
|
R:S ratio |
0.22±0.02 |
- |
|
R:T ratio |
0.18±0.01 |
- |
|
Stem density (g cm−3) |
0.36±0.01 |
- |
|
BEF |
4.85±0.25 |
- |
Download Excel Table
이러한 결과는 줄기나 뿌리에 비해 수관(잎, 가지) 부위의 바이오매스 할당이 높은 초기 조림목의 일반적인 생육 특성을 뒷받침한다(Mokany et al., 2006; Nogueira et al., 2014; Menéndez-Miguélez et al., 2022). 그러나 동일 수종 내에서도 생태형 및 토양 양분 가용성 등 다양한 입지 조건에 따라 지하부와 지상부 바이오매스 간 비율은 달라질 수 있으므로, 입지환경 차이에 따른 공간적 변이를 고려한 추가 연구가 필요하다(Ericsson, 1995). 한편, 초기 조림목의 줄기밀도는 0.36±0.01 g cm−3로, 성숙목의 줄기밀도(0.39-0.43)보다 낮게 나타났다(Park et al., 2005). 이는 수목의 연령 증가에 따라 세포벽 두께, 리그닌 함량 및 심재 비율 증가 등 줄기의 해부학적 특성에 기인한다(Sellin, 1994; Nogueira et al., 2014; Shen et al., 2024).
3. 입목 탄소저장량 추정
조림지 내 개체목들의 근원경과 개발된 바이오매스 상대생장식을 활용하여 임분 수준의 탄소저장량을 추정한 결과, 약 5.25±0.42 Mg C ha−1로 나타났다. 이는 흉고직경과 기존 바이오매스 상대생장식을 이용하여 추정한 탄소저장량(1.30±0.09 Mg C ha−1)과 큰 차이를 보였다. 이러한 결과는 초기 조림목의 생장 특성이 반영되지 않은 기존 바이오매스 추정식을 적용할 경우, 잎과 가지 등 수관 부위에 의한 탄소저장 및 흡수량이 과소평가 될 수 있음을 보여준다. 소나무 초기 조림목의 목질부 탄소저장량은 3.23±0.29 Mg C ha−1로, 연구대상지와 인접한 입지에서 생장한 동일 임령 일본잎갈나무의 목질부 탄소저장량(1.58±0.12 Mg C ha−1)에 비해 약 2배 이상 높은 수치를 보였다(Song et al., 2025). 이는 탄소저장 측면에서 소나무 초기 조림지가 일본잎갈나무에 비해 높은 잠재력을 가질 수 있음을 시사한다. 그러나 향후 지속적인 모니터링을 통해 식생에 의한 탄소흡수량과 토양 미생물호흡에 의한 지하부 탄소배출량을 종합적으로 분석하여 생태계 수준의 순탄소흡수능력을 정량적으로 평가할 필요가 있다.
결 론
본 연구에서는 소나무 초기 조림목의 바이오매스 상대생장식을 개발하고 부위별 바이오매스 구성비를 분석한 뒤, 기존 바이오매스 추정 회귀선과 비교함으로써 조림목의 초기 탄소 분배 특성을 확인하였다. 근원경을 이용한 소나무 초기 조림목 바이오매스 상대생장식 개발은 탄소저장량 추정의 정확성을 크게 향상시켰다. 또한 수관폭을 이용한 바이오매스 추정은 비교적 측정이 용이한 변수를 활용하여 초기 조림목의 바이오매스를 높은 정확도로 예측할 수 있음을 보여주며, 이를 통해 초기 조림지의 탄소저장량을 지속적으로 모니터링하는데 유용하게 활용될 수 있을 것으로 보인다. 본 연구에서 개발된 조림목 대상 바이오매스 상대생장식과 성숙림 내 개체들을 대상으로 개발된 바이오매스 상대생장식과의 추정 회귀선 및 탄소저장량의 비교 분석은 초기 조림목의 잎과 가지에 의한 탄소저장 및 탄소흡수 잠재력이 재평가될 수 있음을 시사한다. 그러나 개발된 초기 조림목 바이오매스 상대생장식은 포천 지역의 7년생 소나무 초기 조림목에 국한되었기 때문에 보다 다양한 입지환경 및 넓은 범위의 임령을 포함하는 바이오매스 상대생장식 개발이 필요하다. 특히, 토양 양분 조건은 지상부와 지하부 탄소 분배에 영향을 미치기 때문에, 향후 토양 입지 환경에 따른 조림목의 형태적 특성 변이에 대한 추가적인 연구가 필요하다. 본 연구에서 개발된 소나무 초기 조림목 상대생장식은 소나무 재조림지 및 복구지 내 초기 탄소저장량 및 흡수량 추정의 정확성을 향상시키고 입지 생산성을 정량평가 및 모니터링하기 위한 유용한 기초자료를 제공한다.
감사의 글
본 연구는 정인욱학술장학재단 ‘산림 Pioneer 육성 장학사업’의 지원으로 수행되었음.
References
Bilgili, E. and Kucuk, O., 2009. Estimating above-ground fuel biomass in young Calabrian pine (
Pinus brutia Ten.). Energy & Fuels 23(4): 1797-1800.


Bjarnadóttir, B., Inghammar, A.C., Brinker, M.-M. and Sigurdsson, B.D., 2007. Single tree biomass and volume functions for young Siberian larch trees (
Larix sibirica) in eastern Iceland. Icelandic Agricultural Sciences 20: 125-135.
Blujdea, V., Pilli, R., Dutca, I., Ciuvat, L. and Abrudan, I., 2012. Allometric biomass equations for young broadleaved trees in plantations in Romania. Forest Ecology and Management 264: 172-184.
Chan, F.C. et al., 2018. Carbon, water and energy exchange dynamics of a young pine plantation forest during the initial fourteen years of growth. Forest Ecology and Management 410: 12-26.
Chave, J., Andalo, C., Brown, S., Cairns, M.A., Chambers, J.Q., Eamus, D., Fölster, H., Fromard, F., Higuchi, N. and Kira, T., 2005. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 145: 87-99.

Claesson, S., Sahlén, K. and Lundmark, T., 2001. Functions for biomass estimation of young
Pinus sylvestris,
Picea abies and
Betula spp. from stands in northern Sweden with high stand densities. Scandinavian Journal of Forest Research 16(2): 138-146.
Cutini, A., Chianucci, F. and Manetti, M.C., 2013. Allometric relationships for volume and biomass for stone pine (
Pinus pinea L.) in Italian coastal stands. iForest-Biogeosciences and Forestry 6(6): 331.
Ericsson, T., 1995. Growth and shoot: root ratio of seedlings in relation to nutrient availability. Plant and Soil 168: 205-214.
Fradette, O., Marty, C., Tremblay, P., Lord, D. and Boucher, J.-F., 2021. Allometric equations for estimating biomass and carbon stocks in afforested open woodlands with black spruce and jack pine, in the eastern Canadian boreal forest. Forests 12(1): 59.
Goodman, R.C., Phillips, O.L. and Baker, T.R., 2014. The importance of crown dimensions to improve tropical tree biomass estimates. Ecological Applications 24(4): 680-698.
Han, S.H. and Park, B.B., 2020. Comparison of allometric equation and destructive measurement of carbon storage of naturally regenerated understory in a
Pinus rigida plantation in South Korea. Forests 11(4): 425.
Juan-Ovejero, R., Elghouat, A., Navarro, C.J., Reyes-Martín, M.P., Jiménez, M.N., Navarro, F.B., Alcaraz-Segura, D. and Castro, J. 2023. Estimation of aboveground biomass and carbon stocks of
Quercus ilex L. saplings using UAV-derived RGB imagery. Annals of Forest Science 80(1): 44.
KFRI (Korea Forest Research Institute), 2014. Carbon emission factors and biomass allometric equations by species in Korea. Seoul, Korea: KFRI.
KFRI (Korea Forest Research Institute), 2016. Strategy for activating the forest carbon offset program in Korea. Seoul, Korea: KFRI. pp. 9-31, 83.
KFS (Korea Forest Service), 2024. Statistical Yearbook of Forestry, Korea Forest Service, Korea: KFS. pp. 204-206.
Kim, S.R., Lee, W.K., Kwak, H.B. and Choi, S.H., 2009. Estimating carbon sequestration in forest using KOMPSAT-2 imagery. Journal of Korean Society of Forest Science 98(3): 324-330.
Madalcho, A.B., Szwagrzyk, J., Maciejewski, Z., Gazda, A. and Bodziarczyk, J., 2023. Allometric scaling models for tree saplings: Assessing potential fodder for ungulate herbivores. Trees, Forests and People 13: 100424.
Mclntire, C.D., Cunliffe, A.M., Boschetti, F. and Litvak, M.E., 2022. Allometric relationships for predicting aboveground biomass, sapwood, and leaf area of two-needle Piñon pine (
Pinus edulis) amid open-grown conditions in central New Mexico. Forest Science 68(2), 152-161.
Menéndez-Miguélez, M., Calama, R., Del Río, M., Madrigal, G., López-Senespleda, E., Pardos, M. and Ruiz-Peinado, R., 2022. Species-specific and generalized biomass models for estimating carbon stocks of young reforestations. Biomass and Bioenergy 161: 106453.
Mokany, K., Raison, R.J. and Prokushkin, A.S., 2006. Critical analysis of root: shoot ratios in terrestrial biomes. Global Change Biology 12(1): 84-96.
Nogueira, L.R., Engel, V.L., Parrotta, J.A., Melo, A.C.G.d. and Ré, D.S., 2014. Allometric equations for estimating tree biomass in restored mixed-species Atlantic forest stands. Biota Neotropica 14(2): e20130084.
Noh, N.J. et al., 2010. Carbon and nitrogen storage in an age-sequence of
Pinus densiflora stands in Korea. Science China Life Sciences 53: 822-830.
Noh, N.J., Kim, C., Bae, S.W., Lee, W.K., Yoon, T.K., Muraoka, H. and Son, Y., 2013. Carbon and nitrogen dynamics in a
Pinus densiflora forest with low and high stand densities. Journal of Plant Ecology 6(5): 368-379.
Noh, N.J., Kwon, B., Yang, A.R. and Cho, M.S., 2020. Effect of planting density on early growth performances of
Zelkova serrata trees. Journal of Korean Society of Forest Science 109(3): 281-290.
Pajtík, J., Konôpka, B. and Lukac, M., 2008. Biomass functions and expansion factors in young Norway spruce (
Picea abies [L.] Karst) trees. Forest Ecology and Management 256(5): 1096-1103.
Park, I.H., Park, M.S., Lee, K.H., Son, Y.M., Seo, J.H., Son, Y. and Lee, Y.J. 2005. Biomass expansion factors for
Pinus densiflora in relation to ecotype and stand age. Journal of Korean Society of Forest Science 94(6): 441-445.
Pati, P.K., Kaushik, P., Khan, M.L. and Khare, P.K. 2022. Allometric equations for biomass and carbon stock estimation of small diameter woody species from tropical dry deciduous forests: Support to REDD+. Trees, Forests and People 9: 100289.
Roxburgh, S.H., Paul, K.I., Clifford, D., England, J.R. and Raison, R.J. 2015. Guidelines for constructing allometric models for the prediction of woody biomass: how many individuals to harvest? Ecosphere 6(3): 1-27.
Sellin, A., 1994. Sapwood–heartwood proportion related to tree diameter, age, and growth rate in
Picea abies. Canadian Journal of Forest Research 24(5): 1022-1028.
Shen, W. et al., 2024. Variation pattern in the macromolecular (cellulose, hemicelluloses, lignin) composition of cell walls in
Pinus tabulaeformis tree trunks at different ages as revealed using multiple techniques. International Journal of Biological Macromolecules 268: 131619.
Silva, M.R.F., McHugh, I., Neto, A.P., Pauwels, V.R.N., Cartwright, I. and Daly, E., 2022. Trading a little water for substantial carbon gains during the first years of a
Eucalyptus globulus plantation. Agricultural and Forest Meteorology 318: 108910.
Song, K.H., Park, M.W., Lim, S.W., Lee, S.T., Chung, S.H. and Noh, N.J., 2025. Developing woody biomass allometric equations for estimating carbon stocks in early-stage
Larix kaempferi plantations in Pocheon region. Journal of Forest and Environmental Science 41(1): 1-7.
Thippawan, S., Chowtiwuttakorn, K., Pongpattananurak, N. and Kraichak, E., 2023. Allometric models to estimate the biomass of tree seedlings from dry evergreen forest in Thailand. Forests 14: 725.
Zianis, D., Xanthopoulos, G., Kalabokidis, K., Kazakis, G., Ghosn, D. and Roussou, O., 2011. Allometric equations for aboveground biomass estimation by size class for
Pinus brutia Ten. trees growing in North and South Aegean Islands, Greece. European Journal of Forest Research 130: 145-160.