서 론
북방계 식물은 빙하기 동안 광범위한 분포를 보였으나, 홀로세의 기온 상승 이후 고산지대와 고위도 지역으로 후퇴하여 제한적으로 분포하게 되었다(Gantsetseg et al., 2020). 물앵도나무는 이러한 북방계 식물 중 하나로, 한반도에서는 북한 평안남도에 분포의 남방한계선이 위치한다. 분포의 경계 지역에 존재하는 개체군은 기후변화로 인한 생육 환경 변동에 취약하므로 향후 지속 가능한 보전을 위해서는 체계적인 현지외(ex situ) 보전 전략이 필수적이다(Kong, 1998; Kim et al., 2011).
일반적으로 아한대 및 온대북부 산림에 서식하는 북방계 식물은 뿌리 및 지상부 생장 속도가 느리고 온난화 상황에 따른 생장 저하 및 낮은 생리적 적응력을 보인다(Dillaway and Kruger, 2011; Pedlar et al., 2024; Goto et al., 2025). 이러한 생리적 제약은 이식 초기 활착 실패로 이어질 가능성이 높으며, 특히 묘목 단계에서의 생장 및 생리적 안정성 확보는 보전 및 재도입의 성공을 결정하는 데 중요하다.
시비는 묘목의 광합성, 간장과 근원경 발달을 촉진하여 초기 활착 및 기존 식생과의 경쟁(Poorter et al., 1995; Quoreshi and Timmer, 2000; Cho et al., 2012)을 향상시킨다. 시비를 통해 공급되는 양분 중 질소는 엽색소 단백질, 전자전달계, RuBisCO 단백질 형성을, 인은 막 용해도, ATP 및 NADPH 생산을 증가시키며, 이로 인해 엽록소 함량 및 최대 전자전달 속도(Jmax; Maximum electron transport rate)와 최대 카르복실화 속도(Vcmax; Maximum carboxylation rate)가 증가하여 광합성의 생화학적 제한이 완화된다(Poorter et al., 1995; Evans, 1989). 이때 시비를 통해 공급된 양분이 전자전달계 단백질보다 카르복실화 효소에 많이 투자되면 Jmax와 Vcmax 간의 불균형이 초래되어 전자전달계의 막힘 현상으로 인한 활성산소종(ROS; Reactive oxygen species)이 발생한다. ROS에 의한 광저해를 회피하기 위해 식물은 Jmax와 Vcmax 의 비율을 유지하는 보수적 조정 전략을 취해 광 흡수량과 이용량 간의 균형을 조절하며 이로 인해 최대양자수율(Fv/Fm; Maximum quantum yield)은 증가하고 비광화학적 형광소멸(NPQ; Non-Photochemical Quenching)은 감소한다(Walker et al., 2014). 시비를 통한 생화학적 과정의 개선은 탄소동화량을 증가와 물질생산량의 증가로 이어지고, 특히 지하부(뿌리)보다 광합성 기관(잎과 줄기)에 더 많은 자원을 배분하여 탄소 획득 능력을 극대화하는 반응이 나타난다(Poorter and Nagel, 2000). 하지만 적절하지 못한 수준의 시비는 양분 이용 효율을 저하시켜 불안정한 H/D율 및 Jmax와 Vcmax 간의 불균형을 초래할 수 있으므로 양묘 과정에서는 수종별 특성을 고려하여 최적의 시비 수준을 구명하고 제시하는 것이 필수적이다(Dumroese et al., 2013; Kwon et al., 2009).
Lonicera 속은 시비 조건에 따라 생장과 생리 반응이 달라지는 것으로 보고되었다. Hayes and Peterson(2020)은 댕댕이나무(L. caerulea)와 L. villosa는 시비 농도 증가에 따른 생장 반응이 크지 않았으나, 분홍괴불나무(L. tatarica)는 지상부 및 지하부 건중량이 유의하게 증가한다고 보고하였다. 또한 L. caerulea의 경우 vermicompost 처리가 엽록소 함량 증가와 광합성 활성의 향상, 생산성 증대되었다(Enescu et al., 2021). 하지만 북방계 관목류에 대한 생리·광합성 수준의 시비 반응 연구는 매우 제한적이며, 특히 물앵도나무은 시비 반응 특성이 전혀 보고되지 않아 보전 및 조림을 위한 적정 시비 수준의 과학적 근거가 부재한 상황이다.
물앵도나무(Lonicera ruprechtiana Regel)는 인동과(Caprifoliaceae) 인동속(Lonicera)에 속하는 낙엽성 관목으로, 중국 동북부와 러시아 동부(아무르, 우수리), 한반도 북부(평안남도, 평안북도, 함경남도, 함경북도)에 분포하며 남한에는 자생하지 않는다. 수고는 약 2 m까지 자라며, 5-6월에 백색 또는 연한 황색의 꽃을 피우고, 7-8월에는 적색의 장과 열매를 맺는다. 북한 두만강 유역의 낮은 산지와 들판에서 비교적 흔히 발견되며, 내한성과 내건성이 뛰어나 중국 북부에서 녹화용 조림수종으로도 널리 활용되고 있다(Gu et al., 2022). 또한 항균 활성을 비롯해 다양한 생리·약리적 효용이 보고되어 활용 잠재력이 크다(Wang et al., 2009).
따라서 본 연구는 북방계 식물인 물앵도나무 묘목의 생장과 생리적 품질 향상을 위한 적정 시비 수준을 규명하고자 하였다. 본 연구에서는 다음 두 가지 가설을 검증하였다. 첫째, 시비 수준 증가는 엽록소 함량과 광합성 생화학적 용량을 향상시켜 광합성률이 특정 시비 범위에서 최대가 될 것이다. 둘째, 충분한 시비 수준은 자원 배분을 지상부 중심으로 전환시켜 T/R율을 증가시킬 것이며 특정 시비 범위에서 생장이 최대가 될 것이다. 이를 구명하기 위해 물앵도나무 묘목을 대상으로 시비 수준에 따른 생장(근원경, 간장의 상대생장량, 물질생산량) 및 생리적 반응(엽록소 함량, 엽록소 형광 반응, 광합성 특성)을 평가하였다.
재료 및 방법
실험에 사용된 식물체는 2023년 국립DMZ자생식물원 온실에서 발아시킨 후 육묘장으로 이식하여 1년간 노지 생육시킨 실생개체로, 2017년 몽골에서 도입하여 국립DMZ자생식물원에 식재한 물앵도나무 개체에서 2022년 채종한 종자를 사용하였다. 형질이 고르고 우수한 실생묘를 직경 143 mm, 높이 160 mm 정사각 플라스틱 용기(Square Large Pot; Hawbunplaza Co., Ltd., Hanam, Korea)에 이식하였다. 실험에 사용한 실생묘의 평균 근원경은 2.64±0.70 mm, 간장은 20.44±4.80 cm이었으며 처리구 별 유의한 차이는 없었다. 토양은 원예상토(Hanareum No.2; Shinsung mineral Co., Ltd., Goesan, Korea)와 마사(2:1, v/v)를 혼합하여 사용하였다.
본 실험은 국립DMZ자생식물원 내에 설치된 W2,000×D14,000×H2,000 규모의 플롯에 50% 차광율의 이중직 비닐 차광막(UV Large Shade Net; Hansol Co., Ltd., Boseong, Korea)을 적용하여 수행하였다. 본 실험에 앞서 차광 수준(0%, 50%, 75%, 95%)에 따른 적정 재배 조건을 검토하기 위한 예비 실험을 동일 장소에서 실시하였으며, 50% 차광 조건이 물앵도나무의 생육 및 생리 반응이 가장 우수하게 나타남에 따라 본 실험의 차광 수준으로 선정하였다(unpublished data).
시비는 수용성 비료인 MultiFeed 20(N:P:K=20:20:20, Haifa chemicals, Israel)을 이용하여 2024년 6월 19일부터 9월 6일까지 총 12주 동안, 주 1회, 4가지 수준(무처리, 500 mg·L−1, 1000 mg·L−1, 2000 mg·L−1)으로 20 L·m−2를 실시하였다. 한여름 고온으로 인해 건조가 심한 시기에는 주 2-3회 20 L·m−2의 관수를 추가로 실시하였다. 실험 개체는 각 처리구당 6반복, 총 24개체를 배치하였다.
고온과 이상기온 현상에 의한 피해를 줄이기 위해 8~9시 또는 17~18시 사이에 관수와 시비를 원칙으로 실험을 수행하였으며, 처리구 내 환기를 위해 차광막 하부를 지면에서부터 20 cm 띄워 공간을 주었다. 실험 기간 중 실험구 내 환경 조건을 기록하기 위하여 온습도 데이터 로거(HOBO U23 Pro v2 U23-001; Onset; Boume, MA, USA)를 사용하였으며, 평균기온은 22.53°C, 최고 기온은 34.20°C, 최저 기온은 13.43°C, 낮 평균 기온은 24.47°C, 밤 평균 기온은 20.68°C, 평균 상대습도는 89.45%였다.
엽록소 함량을 조사하기 위하여 처리구별 세 개체에서 각각 한 장의 잎을 채취한 후 잎 당 0.72 cm2 의 엽편을 5 ml의 DMSO (Dimethyl sulfoxide) 용액이 들어있는 Vial에 담아 65°C의 항온기에서 6시간 동안 처리하여 색소를 추출하였다(Hiscox and Israelstam, 1979). 추출한 용액은 분광광도계(MRX A2000, KLAB Co., Ltd., Republic of Korea)를 사용하여 663 nm, 645 nm의 파장에서 용액의 흡광도를 측정하였고, 이를 통해 엽록소 a, b, a+b 함량 및 a/b를 산출하였다(Arnon, 1949).
시비 처리 수준에 따른 최대양자수율(Fv/Fm)과 비광화학적 형광소멸(NPQ)의 차이는 엽록소 형광반응 측정기(PAM-2100, Heinz Walz GmbH, Germany)를 이용하여 측정하였다. 엽록소 형광 반응은 2024년 9월 말 최상단 신초에서 2~3번째 위치한 건실한 잎을 대상으로 측정하였다. 측정 전 sample clip으로 약 20분간 광을 차단하여 암적응을 유도하고, 광합성계 열방출경로를 불활성화한 뒤 처리구별 6반복으로 실험을 진행하였다. 형광 강도는 Quenching analysis 기법을 활용하여 약 200초 동안 정광과 섬광을 교차 조사하면서 기록하였으며, 이때 Fo는 0.1 μmol·m−2·s−1 이하의 약광을, Fm은 0.8초 동안 8,000 μmol·m−2·s−1의 포화광을 조사하여 측정하였다. 이를 바탕으로 최대양자수율(Fv/Fm)과 비광화학적 형광소멸(NPQ)을 산출하였다(Lichtenthaler et al., 2005).
시비처리에 의한 식물의 광합성 특성 차이는 엽록소 형광 반응을 측정한 잎과 동일한 잎을 대상으로 측정하였다. 챔버 내부 PPFD (Photosynthetic Photon Flux Density)는 1,200 μmol·m−2·s−1, CO2, 농도는 400±2 µmol·mol−1, 공기 유량은 500 μmol·m−2·s−1, 온도는 23±1°C로 유지하여 광포화 상태에서의 순광합성속도(A, net photosynthetic rate), 기공전도도(gs, stomatal conductance), 기공증산속도(E, stomatal transpiration rate)을 측정하였고, 수분이용효율(WUE)은 아래 식을 통해 산출하였다(Kim et al., 2002).
시비처리에 의한 식물의 광합성 특성 차이는 CO2 농도(0-1,400 µmol·mol−1,)에 의한 엽육세포내 CO2-광합성 반응 곡선(A-Ci Curve)으로 비교하였다. 챔버로의 유입 공기유량은 500 μmol·s−1, 온도는 23±1°C로 유지하여 광합성과 엽육 내 CO2 농도(Ci)를 측정하였으며 이를 통해 최대카르복실화속도(Vcmax) 및 최대전자전달속도(Jmax)를 산출하였다(Sharkey et al., 2007).
물앵도나무의 시비 처리 수준에 따른 생장 반응을 조사하기 위해 처리구당 6개체씩 간장과 근원경을 조사하였으며, 불규칙한 근원의 모양에 의한 오차를 줄이기 위해 근원경의 가로와 세로 값의 평균을 산출하였다. 또한 이를 바탕으로 처리구별 상대생장량과 묘목의 건전도를 판단할 수 있는 H/D율을 계산하였다(Bayala et al., 2009).
결 과
엽록소 b 함량은 2,000 mg·L−1 시비 처리구에서 유의하게 높은 값다. 엽록소 a와 a+b 함량은 시비 수준이 높아짐에 따라 증가하는 경향이 있었지만 처리구 간 유의성은 없었다. 엽록소 a/b비는 반대로 시비 수준이 높아짐에 따라 감소하는 경향이 있었지만 처리구 간 유의성을 보이지 않았다(Table 1). Fv/Fm과 NPQ는 시비 처리에서 더 높았지만 통계적 유의성은 없었다(Table 2).
Differences between groups were tested with Duncan’s multiple range tests or, when variances were unequal, with Dunnnett’s T3 test. Statistical significance was set at p < 0.05 and analyses were performed using SPSS (version 21). Values are means ± SD (n=6).
Differences between groups were tested with Duncan’s multiple range tests or, when variances were unequal, with Dunnnett’s T3 test. Statistical significance was set at p < 0.05 and analyses were performed using SPSS (version 21). Values are means ± SD (n=6).
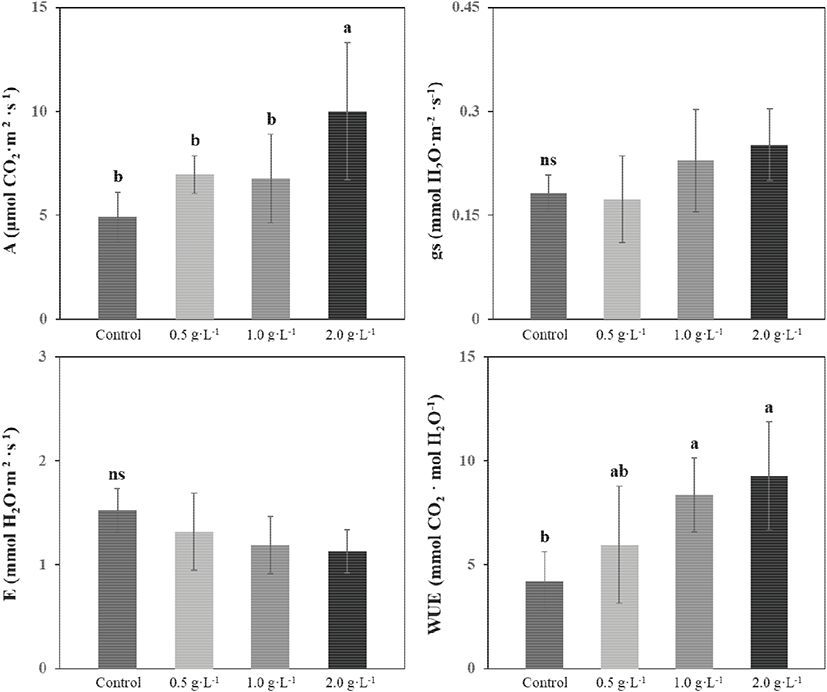
잎을 통한 기체의 이동을 나타내는 gs는 시비 수준이 높아짐에 따라 증가, E는 감소하는 경향을 보였지만 처리구 간 유의미한 차이가 나타나지 않았다(Figure 1). A는 2,000 mg·L−1 시비 처리구에서 10±3.32 μmol CO2·m−2·s−1로 유의하게 높았고 무처리구에서 4.93±1.20 μmol CO2·m−2·s−1로 가장 낮았다. WUE 또한 2,000 mg·L−1 시비 처리구에서 9.26±2.60 mmol CO2·mol H2O−1로 가장 높았고 무처리구에서 4.22±1.41 mmol CO2·mol H2O−1로 가장 낮았다.
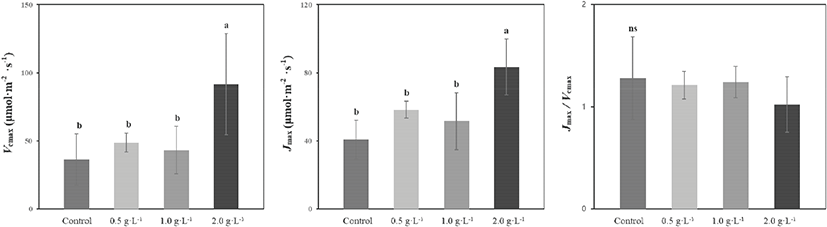
광합성 능력을 제한하는 요소인 Vcmax와 Jmax 모두 2,000 mg·L−1 시비 처리구에서 유의하게 높은 값을 보였다. Jmax/Vcmax는 시비 수준이 증가할수록 감소하는 경향을 보였지만 유의성은 없었다(Figure 2).
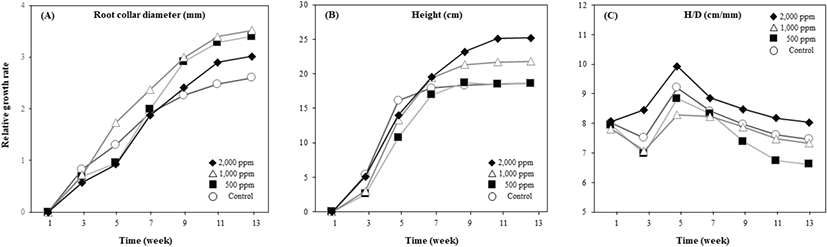
시비 수준에 따른 물앵도나무 근원경의 상대생장량은 1,000 mg·L−1 시비 처리구에서 가장 높았고 대조구에서 가장 낮았지만 처리구별 유의성은 없었다(Figure 3). 시비 수준이 높아질수록 간장의 상대생장량은 증가하는 경향을 보여 2,000 mg·L−1 시비 처리구에서 유의하게 높았다. 시비 수준에 따른 물앵도나무의 H/D율은 2,000 mg·L−1 시비 처리구에서 가장 높고 대조구, 500 mg·L−1, 1,000 mg·L−1 시비 처리구에서 비슷한 값을 보였지만 처리구별 유의성은 보이지 않았다.
지상부와 지하부 모두 2,000 mg·L−1 시비 처리구에서 유의하게 높은 물질생산량을 보였으며 시비 수준이 증가함에 따라 물질생산량도 증가하는 경향을 보였다(Table 3). LWR은 무처리구에 비해 시비 처리구가 약 2.5~2.8배 유의하게 높았다. SWR은 500 mg·L-1 시비 처리구에서 대조구에 비해 낮았지만, 1,000 mg·L−1, 2,000 mg·L−1에서 각각 0.27±0.03, 0.29±0.04 g·g−1 으로 유의하게 높았다. RWR은 대조구가 0.70±0.04 g·g−1 으로 시비 처리구에 비해 유의하게 높은 값을 보였다. T/R율은 대조구에서 유의하게 낮았으며 시비 처리구의 약 0.55~0.70배 수준이었다.
Differences between groups were tested with Duncan’s multiple range tests or, when variances were unequal, with Dunnnett’s T3 test. Statistical significance was set at p < 0.05 and analyses were performed using SPSS (version 21). Values are means ± SD (n=6). zns: non-significance at p < 0.05 among different level of fertilization within the same pigment.
고 찰
본 연구에서 시비 처리는 물앵도나무의 A, WUE, Vcmax, Jmax를 뚜렷하게 증가시켰다. 질소 및 인 시비 처리는 엽 내 질소와 인의 함량을 증가시킨다(Yuan and Chen, 2015). 엽 내 질소 및 인 함량의 증가는 전자전달 단백질 및 카르복실화 효소 생성을 증가시키고 생화학적 제한(Vcmax, Jmax)을 완화하여 광합성 능력을 향상시킨다(Sharkey et al., 2007). Walker et al.(2014)는 메타 분석을 통해 Vcmax와 엽 내 질소 함량 사이의 강한 상관성, 그리고 엽 내 인 함량 증가는 높은 엽 내 질소 함량 조건에서 탄소동화능력을 약 두 배까지 향상시킬 수 있음을 보고하였다. 시비 처리의 영향으로, 물앵도나무는 기공 조절을 통한 생물리학적 과정보다는 생화학 과정의 개선이 뚜렷하게 관찰되었다(Sharkey et al., 2007)
본 연구에서 시비 수준에 따른 Fv/Fm는 처리구 간 차이가 미미하였고 NPQ는 시비 처리구에서 무처리구에 비해 평균값이 낮았지만 개체간 편차가 커 유의성을 보이지 않았다.의성을 보이지 않았다. Fv/Fm의 경우, 0.79에서 0.84가 대부분의 식물의 적정 값이며 이보다 낮을 때는 스트레스 상황을 의미한다(Maxwell and Johnson, 2000; Kitajima and Butler, 1975). 또한 시비 처리로 인한 광화학적 전자전달 및 카르복실화 능력(Vcmax, Jmax)의 향상은 전자전달계 내에 정체되는 과잉 에너지를 감소시켜 NPQ를 감소시킬 수 있다(Walker et al., 2014). 따라서 물앵도나무의 경우, 시비 처리가 광계II의 활성에는 직접적인 영향을 주지 않았지만 전자전달계 내 과잉 에너지로 인한 스트레스를 감소킨 것으로 생각된다.
본 연구에서 물앵도나무는 시비 농도가 높아질수록 Jmax와 Vcmax가 증가했지만 Jmax/Vcmax 은 처리구별로 유의적인 차이를 보이지 않았다. 탄소동화율은 Jmax와 Vcmax 로 정의되는 광합성 기구의 생화학적 용량에 제한을 받는다(Quebbeman and Ramirez, 2016). 이 때 광 수확 및 전자전달과 관련된 Jmax가 카르복실화 효소 단백질인 RuBisCo와 관련된 Vcmax 보다 과도하게 높아지면 카르복실화 제한이 생기며 이로 인해 전자전달계 내에 ROS와 과잉 여기 에너지가 생기게 된다. 따라서, 물앵도나무는 시비 처리 시 Jmax/Vcmax 의 보수적 조정을 통해 광저해 위험을 키우지 않는 범위에서 광합성 능력을 확대하여 광 흡수량과 이용량 간의 균형을 조절한 것으로 보인다(Quebbeman and Ramirez, 2016; Walker et al., 2014).
본 연구 결과, 엽록소 b 함량만이 2,000 mg·L−1 시비 처리구에서 유의하게 높았다. 질소는 엽록소 분자와 광수확 단백질의 주요 구성 물질로 시비 수준이 증가하면 질소 대사의 촉진으로 인해 엽록소의 생합성이 증가될 수 있다(Li et al., 2025). 내음성 식물은 숲의 하층부와 같은 저광도 환경에서 광합성 반응 중심의 유지와 관련된 엽록소 a 생산보다 광 포집과 관련된 엽록소 b 생산에 더 많은 자원을 할당하여 빛 이용효율을 높인다(Pearcy and Sims, 1994). 본 연구에서 물앵도나무는 시비 처리를 통해 양분 흡수가 증가하였을 때 엽록소 b 함량이 유의하게 증가한 것은 50% 차광 조건에 대한 적응 기작이 우선적으로 작동한 것으로 추정되며 엽록소 a/b 또한 유의한 차이는 없지만 고농도 시비 처리 시 평균값이 감소하는 경향을 보였다. 하지만 엽록소 a+b 함량은 시비 처리 수준별 유의성이 없는 것으로 보아 물앵도나무는 시비 처리를 통해 공급된 질소를 엽록소 분자보다 전자전달 단백질 및 카르복실화 효소 생성에 우선 투자하여 광합성 능력을 향상시킴과 동시에 광 흡수량과 이용량 간의 균형을 조절한 것으로 생각된다.
물앵도나무는 시비 수준이 증가함에 따라 상대생장량 및 물질생산량이 대체적으로 증가하는 경향을 보였다. 이는 시비 수준 증가에 따른 광합성 기구의 생화학적 제한 완화가 탄소 동화 능력을 높여, 지상부와 지하부의 건중량 및 간장 증가로 이어졌다고 해석할 수 있다. 낙엽활엽 관목에서 적정 H/D율은 수종에 따라 상이한 것으로 알려져 있다(KFS, 2025). 단풍나무의 경우 수고 11–20 cm에서는 H/D율이 5.5–10 cm·mm, 23–31 cm에서는 8–10 cm·mm 범위가 적정값으로 보고되었다. 또한 때죽나무 25–37 cm 묘목은 8–12 cm·mm, 족제비싸리 30–52 cm 묘목은 8.5–13 cm·mm 수준을 적정 범위로 제시하고 있다(KFS, 2025). 본 연구에서 물앵도나무의 H/D율은 수고가 약 25 cm까지 생장한 2,000 mg·L−1 처리구에서 8–10 cm·mm, 15–20 cm 수준에 도달한 다른 처리구에서는 7–9 cm·mm로 나타나, 기존에 보고된 다른 낙엽활엽 관목의 적정 H/D율과 유사한 범위를 유지하였다. 이러한 결과는 형태적 지표인 H/D율 측면에서 시비 수준에 따른 구조적 불안정 신호가 감지되지 않았음을 의미하며, 따라서 본 연구에서 적용한 시비 범위는 초장의 과도한 증가로 인한 형태적 불균형 없이 생산성을 향상시킬 수 있는 실용적 수준으로 판단된다(Bayala et al., 2009).
본 연구에서 물앵도나무는 2,000 mg·L−1시비 처리구에서 지상부와 지하부 건중량이 모두 유의하게 증가하였다. 또한 대조구에 비해 시비 처리구에서 LWR과 SWR은 유의적으로 높고, RWR은 낮았으며 T/R율 또한 시비 처리구에서 유의하게 높았는데, 이는 공급된 양분이 지상부 생장에 우선적으로 투자되었음을 알 수 있다. 이러한 패턴은 양분이 풍부한 조건에서 식물이 광합성 기관(잎과 줄기)에 더 많은 자원을 배분하여 탄소 획득 능력을 극대화하려는 전략이다 (Poorter and Nagel, 2000).
결 론
본 연구에서 물앵도나무 묘목은 시비 수준이 높아질수록 광합성의 생화학적 용량(Vcmax, Jmax)과 순광합성속도(A), 수분이용효율(WUE), 그리고 지상부·지하부 건중량이 유의하게 증가하였다. 특히 2,000 mg·L−1 처리에서 생장과 물질생산량의 증대가 가장 뚜렷했으며, Jmax/Vcmax 비율과 Fv/Fm, gs, E는 처리 간 유의한 변화가 없어 고농도 시비가 광저해 또는 기공 수준의 스트레스를 유발하지 않고 생산성 향상에 기여했음을 시사하였다. 또한 시비처리구에서 LWR과 SWR이 증가하고 RWR이 감소하여, 공급된 양분이 탄소 획득을 극대화하기 위해 지상부(잎·줄기)로 우선 배분했음을 확인하였다. 다만 본 연구는 단일 생장기와 제한된 온·습도 범위에서 수행되었으므로, 장기 야외 식재지에서의 성장·생존 반응, 더 높은 농도 구간 및 투여 빈도를 다양화하여 추가 실험이 필요하다.