



서 론
지리지역이나 특정 지형에 있어서 기후가 식물 성립과 성장의 제한요인이 될 때 해발고가 특히 중요하다(Barnes et al., 1998). 특정 입지의 해발고는 그 지역에 기후요인의 정보가 없는 경우 식물 분포 및 생육효과를 판정하는 중요한 지표이며, 산지의 유사 지형에 대한 식생을 비교하거나 고위도와 저위도의 식생을 비교하고자 할 때 중요한 결정요인이다(Son et al., 2016). 따라서 고도구배는 기후변화와 식생분포간의 상호관계, 환경변화에 따른 생물다양성의 분포 패턴 연구를 수행하기 위한 가장 중요한 물리적 인자로 인식되고 있다(An et al., 2019). 해발고에 따른 기후인자는 위도, 방위, 경사도 등에 따라 달라지며, 이는 동일한 산지 내에서도 사면방향 등에 따라 다르다(Daubenmire, 1966; Park et al., 2002). 따라서 동일 사면에서도 해발고에 따라 또는 동일 해발고에서도 사면방향에 따라 군집구조가 달라질 수 있다(Webster, 1961; Katagiri and Tsutsumi, 1978; Oh and Jee, 2000).
제주도는 사면과 해발고도에 따라 강수량과 기온이 매우 다양한 분포를 나타내는 지역으로(Choi, 2013), 북사면과 남사면은 경사가 급하게 나타나고, 동사면과 서사면은 상대적으로 경사가 완만하다(Choi, 2011). 또한 반시계 방향으로 상승하는 저기압의 흐름과 관련하여 남서 또는 북서사면 보다는 남동 또는 북동사면에 강수량이 많으며(Park et al., 2014), 해발고에 따라 상록활엽수, 온대낙엽활엽수림대 및 한대침엽수림대까지 여러 기후대의 식물들이 분포하고 있다(Lee et al., 2010).
제주도 상록활엽수림대의 대부분은 계곡을 중심으로 발달하고 있으며(Jeon et al., 2016) 기후변화에 따라 한반도 상록활엽수림의 분포가 확대되며, 졸참나무(Quercus serrata), 신갈나무(Quercus mongolica)림 등의 낙엽활엽수와의 경쟁이 예상되고 있다(Yun et al., 2011). 낙엽활엽수림은 상록활엽수림과 침엽수림대의 사이이며 제일 저지대에 졸참나무와 개서어나무(Carpinus tschonoskii) 군락이 우점하고 있다(Jeju Natural Environment Ecology Information System, 2001). 한라산의 침엽수림은 대면적으로 집단화되어 있으면서 감소율이 가장 큰 지역이며, 제주 지역은 타 지역보다 온도상승이 가장 가파르게 진행되고 있어 한라산이 기후변화로 인한 산림생태계 취약성이 가장 높을 것으로 추정되고 있다(Kim et al., 2019).
한라산의 해발고와 사면에 관한 식생 연구로는 동사면의 해발고도별 식생변화(Lee et al., 2010), 돈내코계곡의 해발고별 식물군집분포(Oh et al., 2007), 남사면 계곡 모새나무 자생지의 식생구조(Song et al., 2013), 동,서사면 좀꽝꽝나무의 생태적 특성(Lee et al., 2011)에 관한 연구 등이 있다. 한라산은 해발고에 따른 식생대의 수직적 발달이 뚜렷하여 생물지리학적⋅생태학적으로 매우 중요한 지역이며, 남동사면은 강수량이 많기 때문에 생물다양성이 높을 것으로 판단된다. 따라서 본 연구는 한라산 남동사면의 해발고별 식생군집구조와 층위구조를 파악하고 해발고별 종다양도 분석을 통해 해발에 따라 변화하는 산림식생구조 특성을 파악하고자 수행하였다.
재료 및 방법
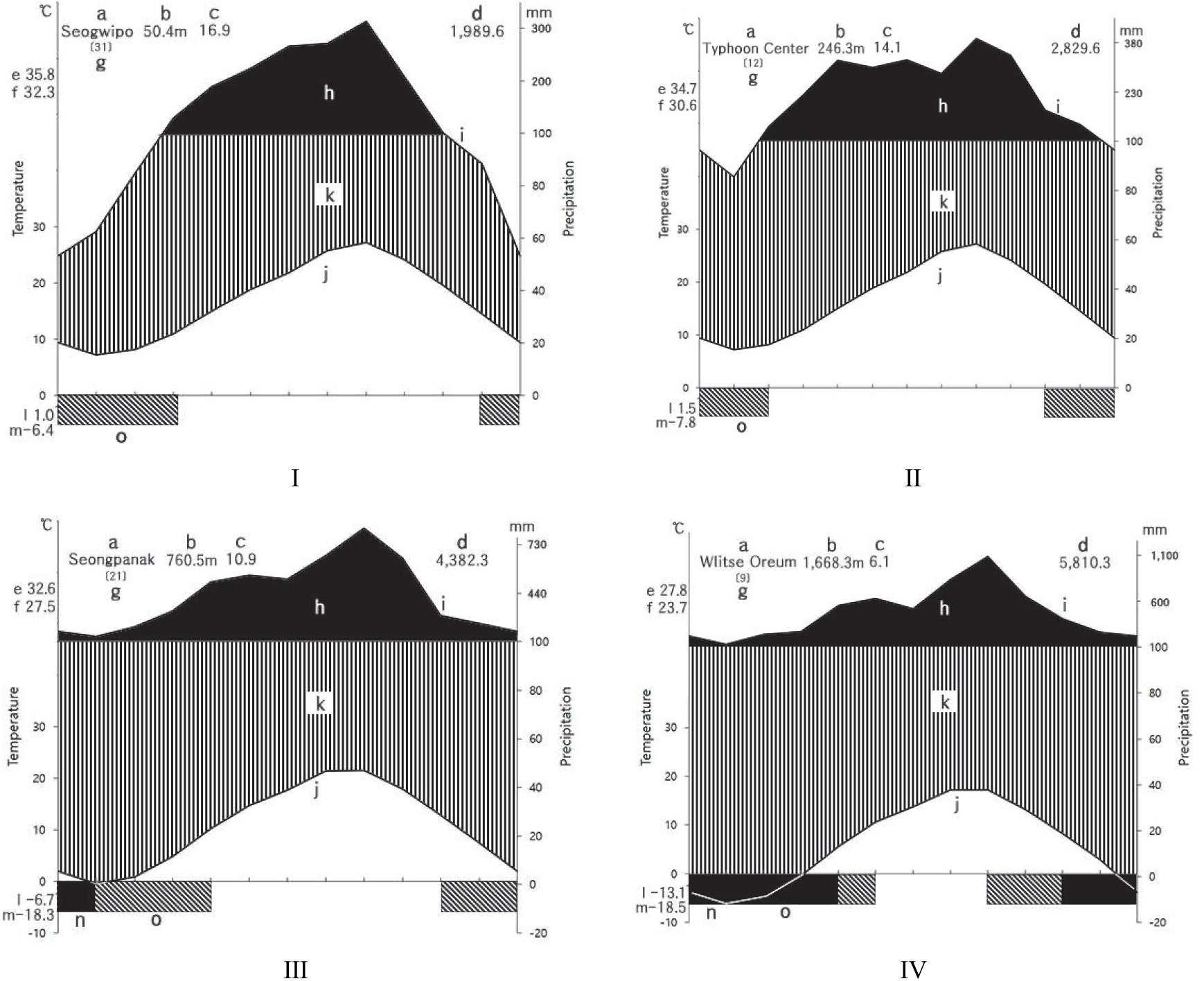
한라산은 우리나라 3대 영산중의 하나로 한반도의 최남단에 위치하고 있으며 해발은 남한에서 가장 높은 1,950 m이며, 백록담을 중심으로 153.332 km2에 달하며, 91.654 km2가 천연보고구역으로 지정되어 있다(Hallasan National Park, 2019). 한라산의 지질은 하위로부터 상위로 탐라층 이전의 전기분석구, 탐라층, 현무암질, 암맥복합체, 탐라층 이후의 후기분석구, 한라산현무암군 및 백록담조면암군으로 구분할 수 있다(Yoon et al., 2005). 한라산은 기후는 해발고도에 따라 강수량과 기온이 매우 다양한 분포를 보이고 있으며(Choi, 2013, Park et al., 2014), 한라산의 지역과 사면, 해발을 기준으로 Choi(2011)가 선정한 기상관측지점을 참고하여 해안지역-서귀포(남사면), 저산간 지역-태풍센터(남동사면), 중산간 지역-성판악(동사면), 고산지역-윗세오름(서,북,남사면)에 대한 기상자료(Korea Meteorological Administration, 2019)를 수집하였다. 기후도(climate diagram)를 작성한 결과(Walter et al., 1975) 대부분 8월에 강수량이 가장 높았으며, 해발이 높아질수록 연평균강수량이 많아졌다. 해발 760m 지점에 위치한 성판악관측소에서 1월, 윗세오름 관측소에서는 1월, 2월, 3월, 12월에 평균기온이 영하로 나타났다(Figure 1).
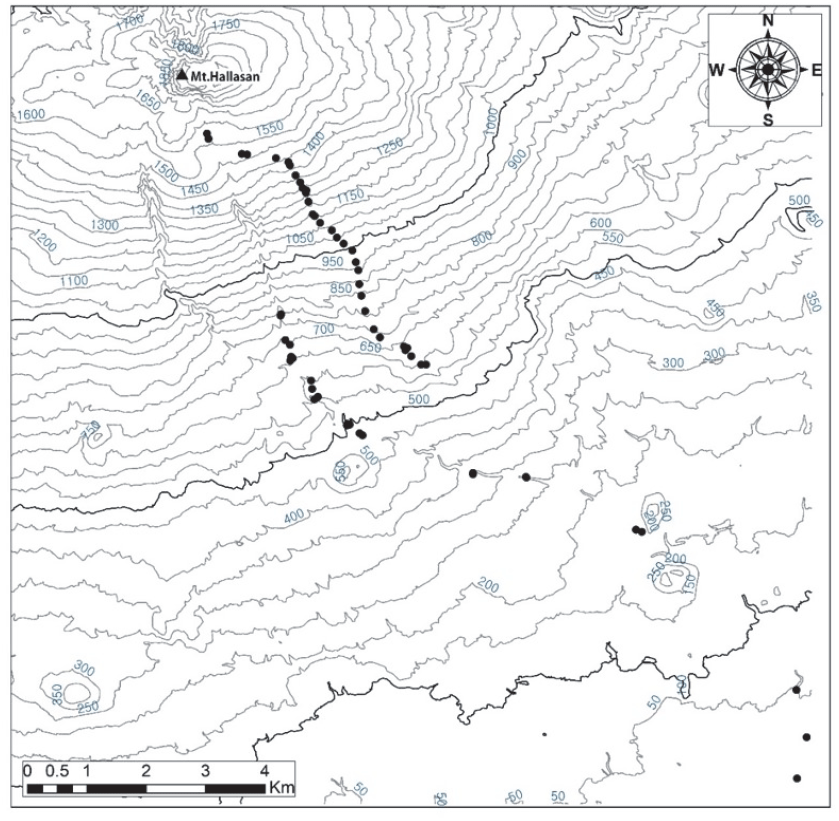
본 조사는 한라산 남동사면의 해발고별 식생구조 및 변화를 알아보기 위하여 2020년 6월에 수행하였으며, 해발 16 m부터 해발 1,565 m까지를 대상으로 해발 100 m 간격마다 2개의 조사구를 20×20 크기로, 총 59개의 조사구에서 식생을 조사하였다. 해발 1,565 m 이상은 출입이 통제되어 있어 조사를 하지 못하였다.
식생조사는 군락의 특징적인 종의 조성을 중시하는 Z-M학파의 식물사회학적 방법(Ellenberg, 1956; Braun-Blanquet, 1964)에 따라 출현하는 구성종의 우점도와 군도 등을 측정하였다. 층위는 교목층, 아교목층, 관목층, 초본층으로 구분하였고, 각 조사지의 해발고도, 방위, 경사를 조사하였다. 종 동정은 원색식물도감(Lee, 2003)과 나무생태도감(Yun, 2016)을 기준으로 하였으며 명명은 국가표준식물목록(Korea Forest Service, 2010a)과 국가생물종지식정보시스템(Korea Forest Service, 2010b)을 기준으로 작성하였다.
군집분류는 클러스터 분석을 하였으며, 적절한 군집 수를 결정하기 위해 Dufrêne and Legendre(1997)의 지표종 분석(Indicator species analysis, ISA)을 실시하였다. 분석 프로그램은 Pc-ord v7.02를 이용하였고, 지표종 분석의 통계적 유의성 평가는 Monte Carlo method를 사용하였다(McCune and Grace, 2002).
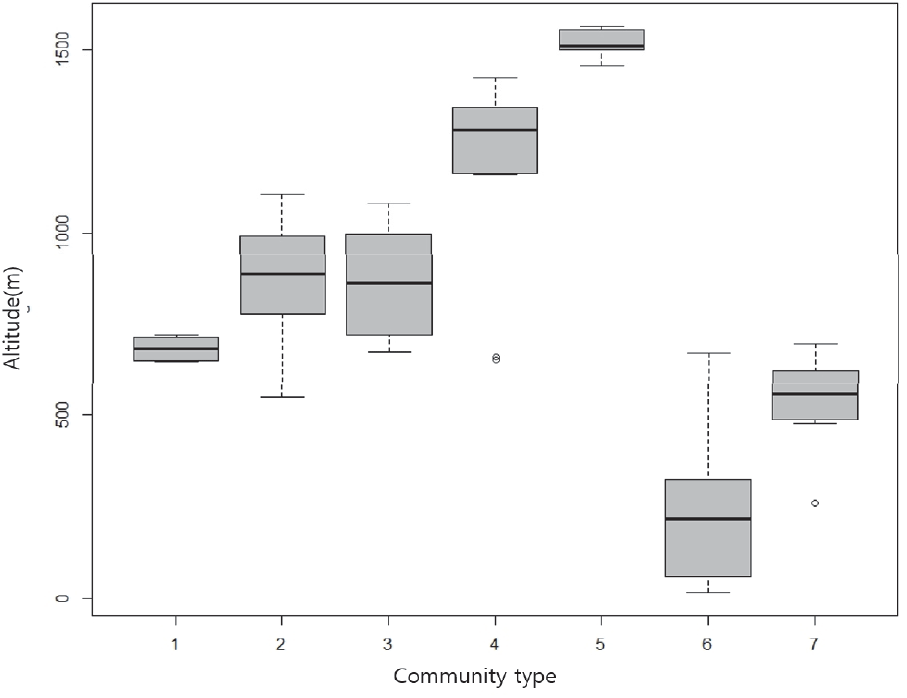
박스플롯(box plot)은 그림으로서 동일항목의 공간적 비교 또는 다양한 실험결과들을 비교할 때 쉽게 활용할 수 있는 방법으로(Park et al., 2014) 각 군집유형별 해발고의 분포를 한 눈에 알아보고자, R v3.5.1을 이용하여 분석하였다.
군집유형과 층위별 주요 구성종의 점유정도를 파악하기 위해 모든 수종을 대상으로 개체수와 피도급의 조합인 우점도 계급을 백분율로 환산한 후 Curtis and McIntosh (1951)의 중요치(Importance Value)를 통합하여 백분율로 나타낸 상대우점치(Importance Percentage)를 층위별로 분석하였다(Brower and Zar, 1977). 상대우점치는 (상대밀도+상대피도+상대빈도)/3로 계산하였으며, 각 층위에 생육하고 있는 수종들의 개체크기를 고하여 (교목층 I.P.×3+아교목층 I.P.×2+관목층 I.P.×1+초본층 I.P.×0.5)/6.5으로 평균상대우점치(Mean Importance Percentage; M.I.P)를 분석하였다. 또한 식생유형별로 평균상대우점치가 높은 수종인을 대상으로 교목층과 아교목층의 해발고별 중요치를 산출하였다.
해발별로 다양성, 우점도, 경쟁 등을 분석하기 위하여 Shannon-Weaver의 종다양도(H’),를 구하고 그에 따른 최대 종다양도(H'max), 종균재도(J'), 그리고 종우점도(1-J′)를 산출하였다(Shannon and Weaver, 1949).
결과 및 고찰
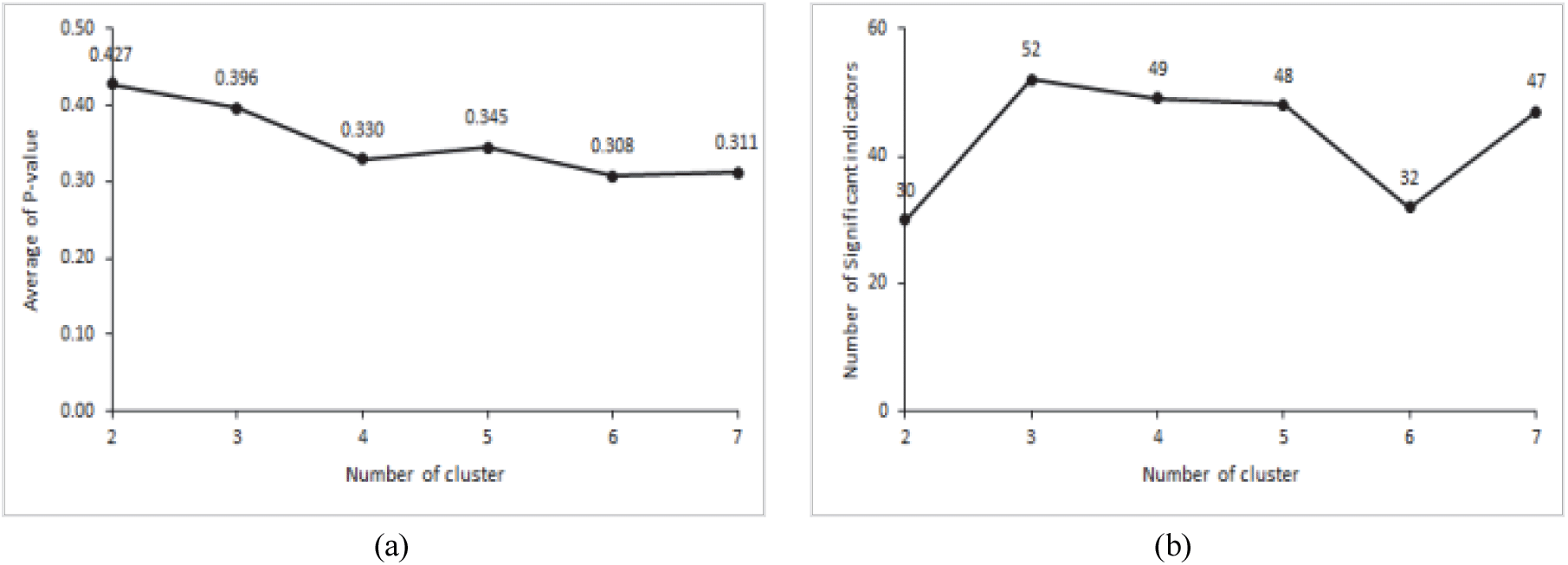
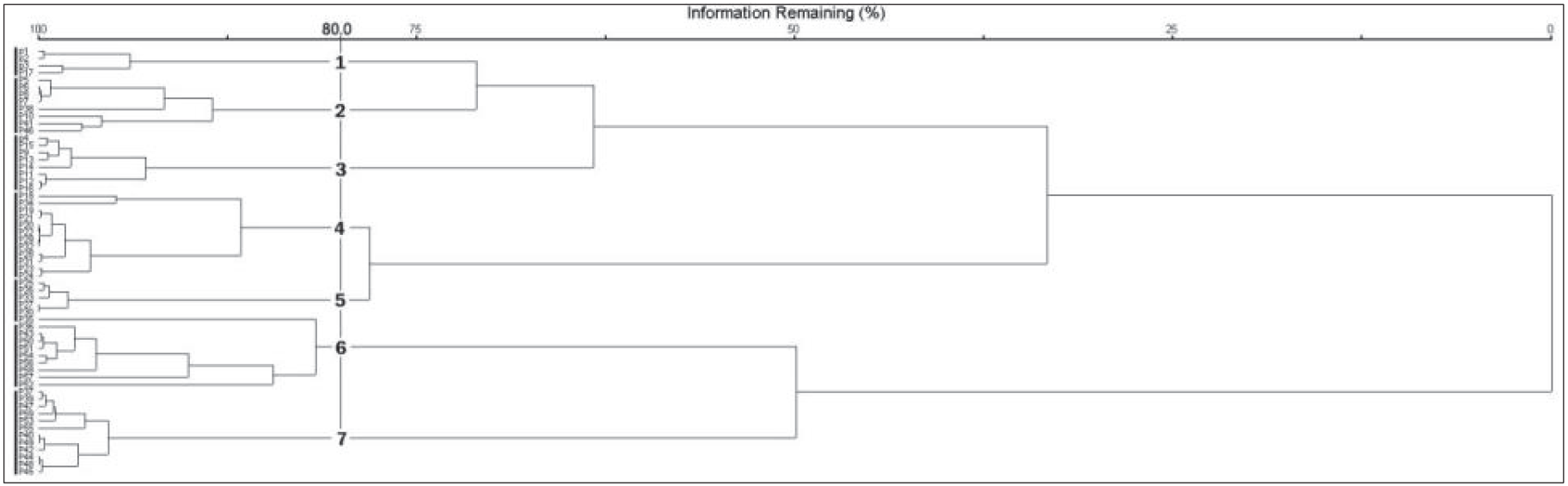
최적의 군집 수를 결정하는 근거자료는 Indicator 값의 평균 P 값이 가장 낮을 때 혹은 유의수준을 만족시키는 수종의 수가 가장 많을 때나, 위의 두 개의 조건을 모두 만족시킬 때이다(Chung and Kim, 2013). 본 연구에서는 군집의 수가 6개 일 때 평균 P 값이 0.308로 가장 낮게 나타났으나, P < 0.01을 만족시키는 수종의 수가 32종으로 낮게 나타났다. 군집의 수가 7개 일 때는 P 값이 0.311로 나타났으며, P < 0.01을 만족시키는 수종의 수가 47종으로 높게 나타나 제주도 남동사면 산림을 7개의 군집으로 분류하였다(Figure 3, Figure 4).
군집명은 각 군집유형에서 P<0.01을 만족시키며, 지표값이 50.0 이상인 수종으로는 교목성 수종을 기준으로 유형1 졸참나무군집, 유형2 개서어나무군집, 유형3 서어나무군집, 유형4 소나무군집, 유형5 구상나무군집, 유형6 구실잣밤나무군집, 유형7 붉가시나무군집으로 명명하였다(Table 1). 지표종은 다양한 유형의 서식처와 환경에 살아가는 종의 개체생태학적 상세한 내용을 이해할 수 있으며, 지표종은 생태계에서 핵심종의 역할을 하는 종이다(Dufrene and Legendre, 1997; Rubio et al., 2011; Cheon et al., 2014). 각 군집에 나타난 지표종으로는 졸참나무군집에서 졸참나무, 참식나무(Neolitseasericea), 홍지네고사리(Dryopteris erythrosora), 때죽나무(Styrax japonicus), 층층나무(Cornus controversa)가 나타났으며, 개서어나무군집에서 개서어나무, 등수국(Hydrangea petiolaris), 나도밤나무(Meliosma myriantha), 서어나무군집에서 서어나무(Carpinus laxiflora), 굴거리나무(Daphniphyllum macropodum), 금창초(Ajuga decumbens)가 나타났다. 소나무군집에서는 소나무(Pinus densiflora), 마가목(Sorbus commixta), 윤노리나무(Pourthiaea villosa), 팥배나무(Sorbus alnifolia), 정금나무(Vaccinium oldhamii), 구상나무군집에서 김의털(Festuca ovina), 털진달래(Rhododendron mucronulatum var. ciliatum), 구상나무(Abies koreana)가 나타났다. 구실잣밤나무군집에서 백량금(Ardisia crenata), 구실잣밤나무(Castanopsis sieboldii), 호자나무(Damnacanthus indicus), 자금우(Ardisia japonica), 후박나무(Machilus thunbergii), 붉가시나무군집에서 붉가시나무(Quercus acuta), 비쭈기나무(Cleyera japonica), 조록나무가(Distylium racemosum) 지표종으로 나타났다. 구실잣밤나무군집은 한국에 분포하는 난온대 상록활엽수림 중 후박나무우점림과 함께 넓은 분포를 나타내는 식물군락이며(Choi, 2013), 구실잣밤나무가 우점하는 군락 내에서 표징종, 구분 종에 따라 식생단위가 분류되고 있으며, 제주도 구실잣밤나무림의 주요 군락으로는 구실잣밤나무-자금우군락(Yun et al., 2011), 구실잣밤나무-후박나무군락, 구실잣밤나무-종가시나무 등으로(Jeon et al., 2016) 지표종인 자금우, 후박나무와 일치하였다.
구실잣밤나무군집의 평균해발은 253.2 m이며, 16-672 m로 가장 낮은 곳에 나타났다. 붉가시나무군집의 평균해발은 541.0 m이며, 258-695 m에 나타났으며, 한라산 돈내코계곡에서는 해발 350-750 m에 나타나고 있었다(Oh et al., 2007). 졸참나무군집의 평균해발은 682.0 m이며, 분포범위는 647-720 m로 가장 좁게 나타났다(Figure 5). 졸참나무의 분포범위는 중부지방에서 600 m, 남부지방은 800 m 이하로 나타났으며 북부로 갈수록 해발고도가 낮아진다(Kim and Kim, 2017). 서어나무군집의 평균해발은 862.6 m이며, 분포범위는 673-1,079 m로 가장 넓게 나타났다. 제주도의 자연환경생태정보시스템(Jeju Natural Environment Ecology Information System, 2001)에 의하면 한라산에 분포하는 낙엽활엽수림의 사면별 해발고도 분포 범위는 동사면 해발 600-900 m과 북사면 해발 600-1,200 m에서는 서어나무, 졸참나무, 개서어나무 등이, 서사면(어리목 및 영실등산로 주변)의 해발 약 900-1,400 m 일대에는 서어나무, 개서어나무, 졸참나무, 물참나무군락 등이, 남사면 해발 약 1,000-1,200 m에서는 서어나무, 개서어나무, 물참나무군락 등으로 구성되어있다. 개서어나무군집의 평균해발은 870.4 m이며, 분포범위는 549-1,105 m에 나타났다. 소나무군집의 평균해발은 1,186.6 m이며, 분포범위는 652- 1,423 m에 나타났다. 한라산에 분포하는 소나무의 사면별로 해발고도 분포범위는 동사면 1,050-1,150 m, 남사면 650-1,400 m, 북사면 750-1,350 m(Sin, 1981; Kim et al., 2019)로 본 조사지의 소나무군집의 분포범위는 남사면과 일치하였다. 구상나무군집의 평균해발은 1,517.6 m이며, 1,456-1,565 m로 가장 높은 곳으로 나타났다. 한라산의 온대림은 해발 600m-1,500 m 범위로(Yim, 1985), 온대림 수종인 졸참나무, 서어나무, 개서어나무, 소나무가 비슷한 고도에서 나타난 것으로 판단되었다. 주요 국립공원 아고산대에 자생 하고 있는 구상나무는 해발고도의 범위가 넓으며, 주로 1,100 m-1,500 m 구간에 주로 분포하고 있었으며(Park et al., 2019), 한라산 동사면의 경우 1,350 m 이상에서 출현하고 있었다(Lee et al., 2010).
클러스터에 의해 분류된 7개의 군집유형을 대상으로 중요치(Importance value)를 산출한 결과(Table 2) 군집유형 1의 경우, 교목층은 졸참나무 35.3, 층층나무 9.6, 섬벚나무(Prunus takesimensis) 8.3 등으로, 아교목층은 동백나무(Camellia japonica) 25.3, 사스레피나무(Eurya japonica) 11.9, 개서어나무 8.9 등으로, 관목층 참식나무 18.2, 굴거리나무 16.4, 사스래피나무 15.4 등으로, 초본층 마삭줄(Trachelospermum asiaticum) 9.3, 졸참나무 4.2, 참식나무 3.0 등으로 나타났다. 제주도 중산간 지역의 졸참나무는 넓은 지역에 자라고 있었지만 표고 재배를 위해 지나치게 벌채되어 개체수가 많이 감소하였다(Korea Forest Research Institute, 2009). 본 연구결과에서는 졸참나무군락으로 유지될 것으로 판단되었으나, 제주도 어리목과 성판악지구에서는 졸참나무의 세력이 약해지면서 서어나무류의 천이가 진행될 것으로 판단되었으므로(Song, 2014), 지속적인 연구가 필요할 것으로 판단되었다.
군집유형 2의 경우, 교목층은 개서어나무 53.5, 섬벚나무, 솔비나무(Maackia faurie), 소나무 7.5 등으로, 아교목층 단풍나무(Acer palmatum) 12.8, 개서어나무 10.1, 당단풍나무(Acer pseudosieboldianum) 9.3 등으로, 관목층 굴거리나무 16.6, 사스레피나무 13.5, 주목(Taxus cuspidata) 12.9 등으로, 초본층 굴거리나무 3.0, 주목 2.2, 개서어나무 1.4 등으로 나타났다. 본 유형은 개서어나무군락으로 유지될 것으로 판단되었으며 영실, 성판악 지역에서 한동안 개서어나무의 세력이 유지 될 것(Lee et al., 2014)이라는 연구와 일치하였다.
군집유형 3의 경우, 교목층 서어나무 53.8, 개어서나무 14.8, 소나무 10.9 등으로, 아교목층 서어나무 12.9, 단풍나무 12.4, 굴거리나무 12.2 등으로, 관목층 굴거리나무 21.9, 사스레피나무 14.5, 새덕이(Neolitsea aciculata) 8.5 등으로, 초본층 제주조릿대(Sasa palmata) 26.6, 굴거리나무 4.5, 사스레피나무 3.9 등으로 나타났다. 개서어나무 군락과 서어나무군락에서 초본층의 제주조릿대의 중요치가 높았으며 이는 서어나무류의 우점군락은 냉온대 남부저산지대의 토지적 잠재자연식생을 구성하는 단위식생으로 알려져 있으며(Choi et al., 2015), 서어나무와 개서어나무가 졸참나무 보다 우세하면서 하층에는 제주조릿대가 우점한다는 견해와 일치하였다(Kim and Lee, 2006; Kim, 2006).
군집유형 4의 경우, 교목층 소나무 62.1, 산개벚지나무(Prunus maximowiczii) 7.6, 마가목 6.1 등으로, 아교목층 마가목 9.8, 산개벚지나무 9.1, 팥배나무(Sorbus alnifolia) 8.7 등으로, 관목층 주목 9.1, 굴거리나무 7.8, 참꽃나무(Rhododendron weyrichii) 4.6 등으로, 초본층 제주조릿대 42.5, 굴거리나무와 주목 1.5, 개서어나무 1.0 등으로 나타났다. 한라산 소나무군락은 우리나라 중부지방 소나무림에 비해 서어나무, 꽝꽝나무, 제주조릿대의 피도가 높게 나타나는 것이 특징이다(Song et al., 2012). 또한 서어나무 군락으로 천이가 될 것으로 판단한 연구가 있으나(Kim et al., 1982) 본 연구에서는 소나무의 중요치가 낙엽활엽수종에 비해 높게 나타났으므로 소나무군락 유지될 것으로 판단되었다.
군집유형 5의 경우, 교목층 구상나무 45.5, 소나무 25.6, 사스래나무(Betula ermanii) 19.2 등으로, 아교목층 구상나무 59.7, 산딸나무(Cornus kousa) 20.1, 사스래나무와 노린재나무(Symplocos chinensis f. pilosa) 10.1 으로, 관목층 털진달래(Rhododendron mucronulatum var. ciliatum) 24.1, 산철쭉(Rhododendron yedoense f. poukhanense) 11.5, 구상나무 10.1 등으로, 초본층 제주조릿대 48.0, 호장근(Fallopia japonica) 5.1, 구상나무 2.8 등으로 나타났다. 한라산 진달래밭 지역과 영실지역 두 지역에서 교목층은 구상나무, 관목층은 주목, 초본층은 제주조릿대의 중요도가 가장 높게 나타난 Song et al.(2014)의 연구와 일치하였다. 본 군락에서 출현하는 시로미는 한반도 북부와 한라산에 격리 분포하고, 해발 1,700-1,900 m 종으로 지구온난화가 지속될 경우 난온대성 수종과 경쟁하거나, 생리생태적인 스트레스로 적응하지 못하고 멸종될 위기에 있으므로(Kong, 2002) 지속적인 연구가 필요할 것으로 판단되었다.
군집유형 6의 경우, 교목층 구실잣밤나무 51.6, 붉가시나무 8.7, 조록나무 7.4 등으로, 아교목층 구실잣밤나무 12.2, 동백나무 12.0, 조록나무 9.1 등으로, 관목층 조록나무 9.7, 동백나무와 사스레피나무 5.5, 구실잣밤나무 4.7 등으로, 초본층 사스레피나무 2.5, 동백나무 2.4, 구실잣밤나무 1.6 등으로 나타났다. 가거도(Lee et al., 2015)와 홍도(Kim et al., 2018)의 구실잣밤나무군락의 구성종인 붉가시나무, 동백나무, 후박나무, 참식나무, 황칠나무, 사스레피나무 등과 일치하였다.
군집유형 7의 경우, 교목층 븕가시나무 53.5, 구실잣밤나무 13.1, 참가시나무(Quercus salicina) 6.9 등으로, 아교목층 16.1, 붉가시나무 12.1, 황칠나무(Dendropanax morbiferus) 10.1 등으로, 관목층 동백나무 17.5, 조록나무 12.6, 사스레피나무 11.2 등으로, 초본층 콩짜개덩(Lemmaphyllum microphyllum)굴 9.2, 붉가시나무 5.4, 동백나무 4.9 등으로 나타났다. 보길도(Kim et al., 2000)와 홍도(Lee et al., 2010)의 붉가시나무군락의 구성종인 황칠나무, 동백나무, 콩짜개덩굴, 사스레피나무 등이 일치하였다.
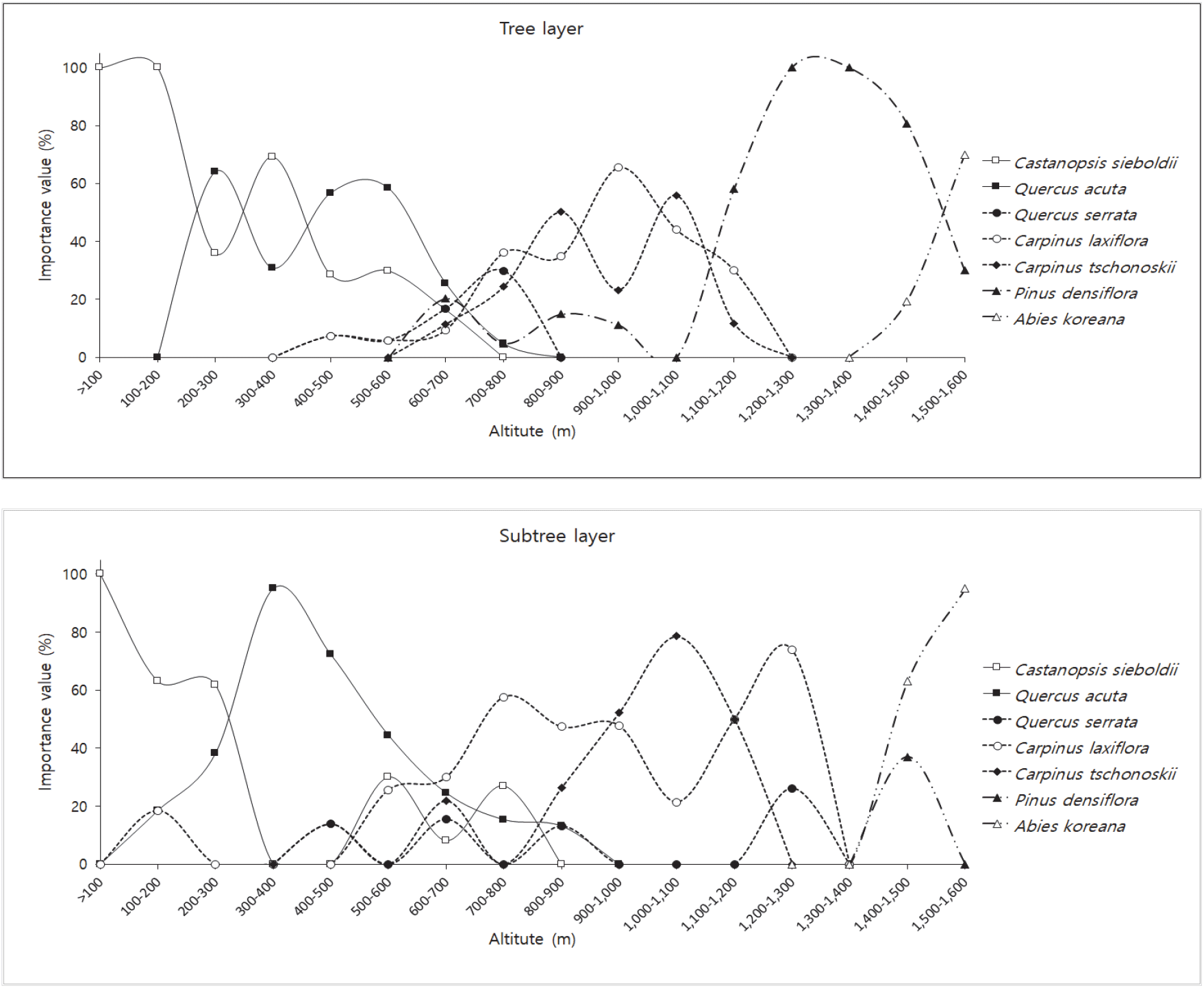
군집유형별로 상대우점치(Mean importance Percentage)가 높은 수종인 구실잣밤나무, 붉가시나무, 졸참나무, 서어나무, 개서어나무, 소나무, 구상나무를 대상으로 교목층과 아교목층의 해발고별 중요치를 산출하였다(Figure 6).
100 m 미만에서부터 600 m까지는 구실잣밤나무와 붉가시나무의가 주를 이루었으며, 400-500 m에서 부터 졸참나무와 서어나무가 출현하였다. 해발 600-700 m 구간에서 교목층의 구실잣밤나무와 붉가시나무의 중요치가 급격하게 낮아지며, 해발 700-800 m부터 교목층의 서어나무, 졸참나무, 개서어나무의 중요치가 높게 나타났으며 아교목층에서 구실잣밤나무와 붉가시나무가 나타나고 있었다. 800-900 m까지 붉가시나무가 출현하였는데 이러한 결과는 붉가시나무의 경우 난대성 상록활엽수종 중에서는 내한성이 비교적 강한수종으로 난대수종의 분포 북한계선에 인접한 내륙이나 난대지역의 고산지대에서 자랄 수 있다(Oh, 1994; Oh and Kim, 1996;, Yeo, 2005, Kim and Park, 2019)는 연구결과와 일치하였다. 1,000-1,100 m 구간 교목층의 개서어나무(55.9)와 서어나무(44.1)의 중요치의 차이가 크게 나타나지 않았는데 한라산 남사면의 서어나무군락과 개서어나무군락은 해발 1,000-1,100 m 부근에서 혼생하여 분포한다(Jeju Natural Environment Ecology Information System, 2001)는 것과 일치하였다. 1,100-1,200 교목층에서 소나무(58.1)의 중요치가 높게 나타났으며, 한라산국립공원 내 해발고도에 따른 소나무림 분포면적은 1,010~1,400 m 구간의 면적이 가장 넓게 나타났으며(Kim et al., 2019), 한라산 소나무림의 분포 중심 해발고도가 1,200 m에 있다는 연구결과와 일치하였다(Kim et al., 1982; Song et al., 2012; Kim et al., 2019). 1,400-1,500 m 구간에서 교목층은 소나무(80.8)가, 아교목층에서는 구상나무(63.0)의 중요치가 높게 나타났다. 1,500-1,600 m 구간에서는 교목층과 아교목층에서 구상나무의 중요치가 높게 나타났다. Song et al.(2010)의 한라산 구상나무림의 분포면적 조사 결과 1,500-1,600 m에서 가장 높은 비율이 나타난다고 하였다.
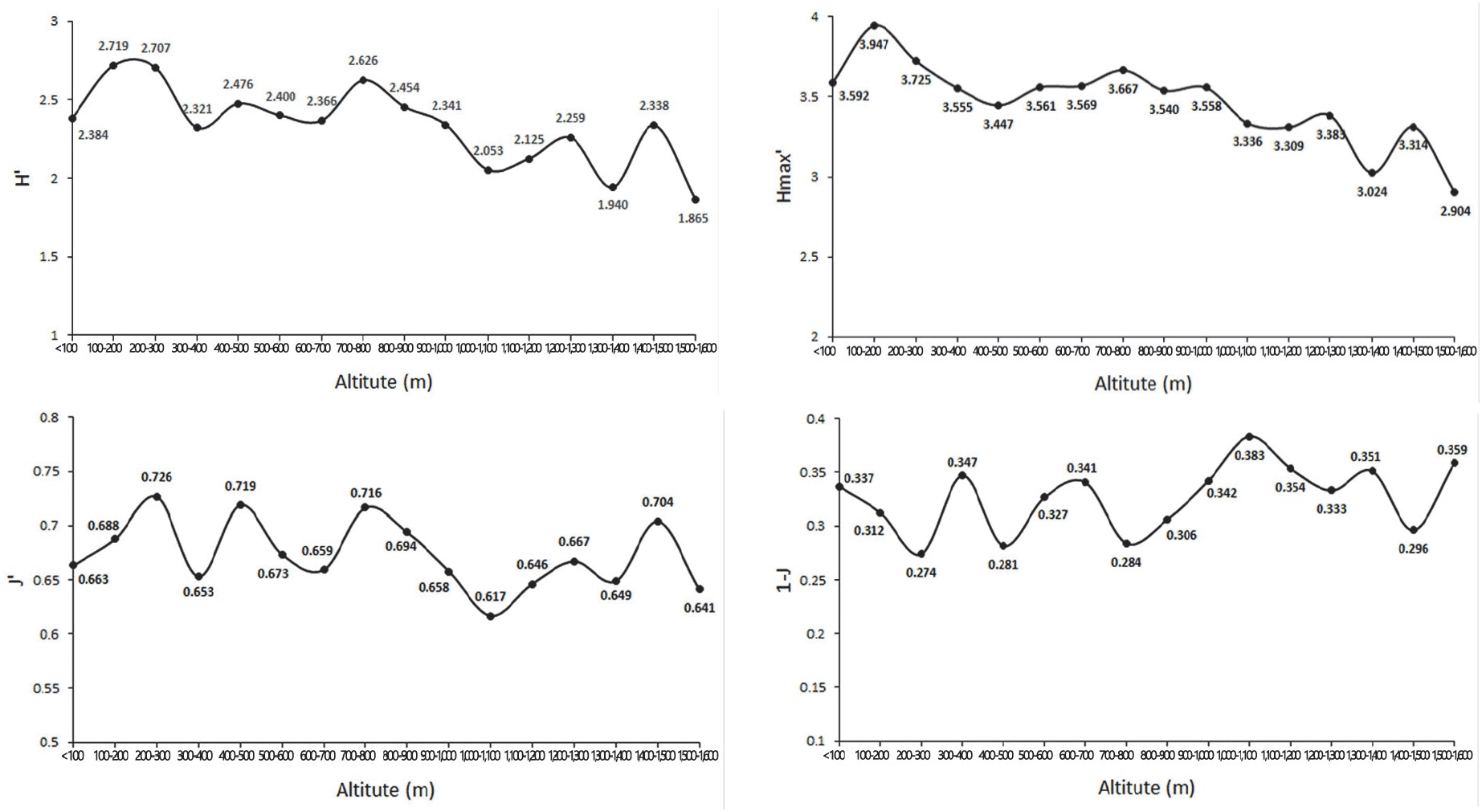
종다양도는 임분의 안정도를 나타내는 지표로서 어느 지역의 종다양도가 높다는 것은 종간 경쟁이 심한 것으로 생태적으로는 건강하다는 의미를 가진다(Song et al., 2019; Moon, 2001). 일반적으로 지구상의 위도와 고도가 낮아질수록 산림군집의 종다양성은 증가하는 경향을 보이는데(Lee et al., 1996), 본 연구에서도 해발이 높아질수록 종다양도가 낮아지는 경향이 나타났다(Figure 7). 이러한 결과는 초본층의 제주조릿대의 중요치가 점차적으로 높게 나타나는데 조릿대의 경우 상록성인 잎이 지피층에 도달하는 빛을 사계절 연속적으로 차단해 지피층에 발생한 치수를 현저하게 피압해 타식물의 생장 및 종다양성에 영향을 주었기 때문인 것(Song et al., 2019; Park et al., 2012)으로 판단되었다.
Odum(1983)은 같은 곳에 형성된 생태계에서도 주변부와 추이대의 경우 종의 수와 밀도가 높아지는 경향이 있다고 하였는데 해발 700-800 m는 붉가시나무군락에서 서어나무, 졸참나무, 개서어나무 혼합림으로, 1,200-1,300 m는 소나무, 서어나무 혼합림에서 소나무군락으로, 1,400-1,500 m는 소나무군락이지만 교목층과 아교목층에서 구상나무가 나타나는 추이대로 종다양도가 높아지는 경향이 나타난 것으로 판단되었다.
결 론
고도구배는 기후변화와 식생분포간의 상호관계, 환경변화에 따른 생물다양성의 분포 패턴 연구를 수행하기 위한 가장 중요한 물리적 인자로 해발고에 따른 기후인자는 위도, 방위, 경사도 등에 따라 달라지며, 이는 동일한 산지 내에서도 사면방향 등에 따라 다르다. 제주도는 사면과 해발고도에 따라 강수량과 기온이 매우 다양한 분포를 나타내는 지역으로(Choi, 2013) 여러 기후대의 식물들이 분포하고 있다. 따라서 본 연구에서는 한라산 남동사면의 해발고별 식생군집구조와 층위구조를 파악하고 해발고별 종다양도 분석을 통해 해발에 따라 변화하는 산림식생구조 특성을 파악하고자 수행하였다.
군집유형으로는 유형1 졸참나무군집, 유형2 개서어나무군집, 유형3 서어나무군집, 유형4 소나무군집, 유형5 구상나무군집, 유형6 구실잣밤나무군집, 유형7 붉가시나무군집으로 구분되었다. 중요치 분석 결과 졸참나무군집은 아교목층의 개서어나무에 의해 천이가 진행될 것으로 판단되므로 지속적인 연구가 필요할 것으로 판단되었다. 구상나무군집의 분포범위는 1,456-1,565 m에 집중적으로 나타났으며, 한라산 동사면의 경우 1,350 m 이상에서 출현하여(Lee et al., 2010) 사면별로 분포범위의 차이가 있는 것으로 판단되었다. 종다양도의 경우, 추이대에서 종다양도가 높아지는 것으로 판단되었으며 추이대는 양쪽 군집에서 관찰되지 않는 특유의 식물 종들이 분포하는 경우도 있기 때문에 지속적인 관찰이 필요할 것으로 판단되었다.
최근 기후환경 변화에 의해 식물의 분포와 생육범위가 변화여 산림 식생대가 변하고 있다. 한라산은 해발고에 따른 식생대의 수직적 발달이 뚜렷하여 생물지리학적⋅생태학적으로 매우 중요한 지역으로 식생대의 변화 연구에 필요한 기초 자료를 지속적으로 구축할 필요가 있다. 따라서 해발고와 여러 사면별로 한라산의 산림식생구조, 개체군 분포 등에 관한 후속 연구가 필요할 것으로 판단되었다.