



서 론
우리나라에서 시설양묘에 의한 용기묘 생산 비율은 2013년 35%에서 2020년 현재 72% 수준까지 2배 이상 증가하였으며(KFS, 2021), 이는 우수한 묘목 생장, 양묘기간의 단축, 자동화 및 노동력 절감, 기후변화에 따른 묘목피해 저감 등 묘목 생산과정의 이점뿐만 아니라 조림 후 높은 생장과 우수한 조림 성과를 가져올 수 있기 때문이다(Haase et al., 2006; Salifu and Jacobs, 2006).
용기묘와 노지묘 생산에 의해 우리나라는 최근 약 2만 ha/년 이상 조림사업을 실시하고 있으며, 활엽수 조림은 최근 5년간(2016~2020) 약 38%로 이전 5년(2011~2015년) 대비 약 13%가 감소하였다(KFS, 2020a). 그러나 고부가가치 활엽수 용재의 수요는 반대로 증가하고 있다. 활엽수는 수종이 다양하지만, 용재 생산을 위한 대표 대경재 수종이 많지 않다. 또한, 채종원산 종자 공급량이 적고, 단위면적 당 묘목 생산량이 낮기 때문에 양묘현장에서 생산이 감소하고 있으며, 조림 후 생존율이 낮고, 풀베기 등 사후관리의 어려움이 있어 묘목 생산량과 조림 면적이 감소하는 실정이다(Cho et al., 2017).
졸참나무, 물푸레나무, 느티나무는 우리나라 온대 주요 낙엽 활엽수종으로 목재적 가치가 높은 고급 용재수를 생산할 수 있는 잠재력을 가지고 있다(Bae et al., 2006; Yang et al. 2016). 또한, 노지 양묘기술이 확보되어 있기 때문에 수종별 적정 용기 규격 구명을 통해 원활한 조림목 수급이 가능하며(Cho et al. 2018), 양묘 및 조림 현장에서도 활엽수 대경재 생산을 위한 용기묘 시업기술 개발 요구가 증가하고 있는 실정이다(Cho et al. 2012; Noh et al., 2020). 따라서 활엽수 묘목품질과 생산성 향상뿐만 아니라 조림 확대 및 용재 생산기반을 구축하기 위해서는 시설양묘에 의한 활엽수 용기묘 생산이 필요하며, 이를 위해서는 다양한 시설양묘 기초 연구가 요구된다.
시설양묘과정에서 광, 수분, 온도 등의 생육환경 조절뿐만 아니라 용기의 종류 및 배양토 등의 재료와 시비 및 관수체계 등의 기술도 용기묘 품질에 중요한 영향을 미친다. 특히, 용기의 생육밀도, 구 용적 등에 따른 규격은 묘목의 생육환경에 영향을 미쳐 무기환경과 양분 조건이 변화하여 수종별 생리 특성 및 생육 반응이 달라지며, 묘목품질 차이가 나타나는 것이다(Romero et al., 1986). 또한, 용기 규격은 묘목이 생장하는 동안 생육환경 변화뿐만 아니라 관수 및 시비 효율, 뿌리 생육공간, 양분저장능력 등에 영향을 주며, 이에 묘목품질의 변화가 나타나는 것이다. 우량 묘목을 생산하기 위해서는 용기 규격의 여러 요인에 대한 상호작용 영향 분석이 필요하며, 이러한 요인들을 동시에 만족시킬 수 있는 최적 용기 규격 구명 연구가 이루어져야 한다.
시비는 제한된 생육상토에서 자라는 용기묘 생산과정에서 무기 양분 공급의 중요성은 아무리 강조해도 지나치지 않다. 용기 규격은 시비에 의한 양분 공급 차이를 가져오며, 용기묘의 생장 속도 및 방식이 달라질 수 있다(Haase and Rose, 1995). 이에 따라 묘목품질 또한 변화되는 것이다(Landis et al., 1989). 시비 및 관수에 의한 원활한 양분 공급을 위해서는 일반적으로 생육밀도가 낮으면서, 용적과 구 직경이 큰 용기가 선호된다(Apholo and Rikala, 2003). 생육공간이 넓을수록 관수 및 시비 효과가 증가되며, 양분을 묘목으로 공급하는 생육상토의 양과 표면적이 넓을수록 양분의 저장이 유리하기 때문이다. 그러나 시비 효과 향상을 위해 용기의 생육밀도를 낮추고 구 직경을 증가시키면, 단위면적 당 생산량이 감소하기 때문에 묘목품질, 시비 효과, 묘목 생산량이 동시에 고려된 적정 용기 규격의 구명이 필요한 것이다. 용기에 따른 양분저장능력(nutrient loading)은 시비와 관수를 중심으로 양묘기술에 의해 결정되며(Park et al., 2012), 그 중 용기 규격은 직·간접적인 영향을 미친다. 이러한 양분저장능력에 따라 양묘과정 이후 용기묘의 활착, 생장, 하층식생과의 경쟁력 및 내건성, 내한성 등 환경 스트레스 저항성 향상뿐만 아니라 식재 스트레스 저감 효과가 조림지에서 발현되기 때문에 적정 양분 공급에 의한 고품질 용기묘 생산과 함께 조림 성과 향상을 기대할 수 있다(Luis et al., 2009; Salifu and Timmer, 2003b). 양분과 관련된 연구는 양묘와 조림이 연계되어 과거부터 꾸준히 진행되고 있다(Epstein, 1972; Timmer et al., 1991). 일반적으로 수종별 임계농도 이하에서는 묘목 내 질소 농도와 함량이 높을수록 조림 후 생장이 우수하였으며(Switzer and Nelson, 1963; Van den Driessche, 1984), 묘목의 질소 농도와 함량은 조림 후 생장을 예측하는 지표로 활용되고 있다.
본 연구에서는 졸참나무, 물푸레나무, 느티나무를 대상으로 생육밀도와 용적이 다른 16 종류의 용기에서 생산된 묘목의 질소 양분 특성을 조사·분석하여, 용기의 생육밀도와 용적에 따른 질소 양분 특성 변화 및 두 요인간의 상호작용을 구명하고, 다중회귀분석기법을 적용하여 생육밀도와 용적 기준의 적정 용기 규격을 제시하고자 수행하였다.
재료 및 방법
시험지는 국립산림과학원 산림기술경영연구소 시설온실(N 37º 45′, E 127º 10′)이다. 공시 수종은 졸참나무(Quercus serrata), 물푸레나무(Fraxinus rhynchophylla) 및 느티나무(Zelkova serrata)이다. 졸참나무는 충북 충주시, 물푸레나무는 충남 공주시, 느티나무는 세종특별자치시에서 2012년 가을 채종한 종자이며, 세 수종 모두 종묘사업실시요령(KFS, 2020b) 종자품질기준 합격 종자를 이용하였다.
용기의 생육밀도와 용적 처리를 위해 요인설계(factorial design) 방법을 이용하였다. 용기는 100, 144, 196, 256본/m2의 4가지 생육밀도(growing density)와 220, 300, 380, 460 cm3/구의 4가지 용적(cavity volume)을 달리하여 제작한 총 16종류 규격이다. 용기의 생육밀도와 용적 처리는 노지묘 생육밀도 기준(KFS, 2020b)과 우리나라 활엽수 6수종 용기 구명 선행 연구 결과(Cho et al., 2012)를 참고하였다. 활엽수 노지묘 1년생은 대부분 m2 당 100본 이하의 낮은 생육밀도이지만, 용기묘는 상대적으로 높은 생육밀도에서도 노지묘보다 품질이 우수한 묘목을 생산할 수 있으므로 약 2배인 m2 당 256본까지 처리하였다. 용적은 수종별 뿌리 형태 및 부피량에 따라 다르지만 약 100 cm3 전후 차이에서 묘목품질의 변화가 관찰된다는 연구 결과(Landis et al., 1990)를 참고하여 용적 처리를 실시하였다. 용기의 생육밀도와 용적 처리를 위해 받침[Figure 1(A)]과 원기둥형 플라스틱 재료[Figure 1(B)]를 이용하였으며, 플라스틱 용기 구(cavity)의 용적별 특성은 Table 1과 같다.

| Volume (cm3) | Diameter (cm) | Depth (cm) | Depth/Diameter ratio |
|---|---|---|---|
| 220 | 4.80 | 12.2 | 2.54 |
| 300 | 5.34 | 13.4 | 2.51 |
| 380 | 5.86 | 14.1 | 2.41 |
| 460 | 6.27 | 14.9 | 2.38 |
양묘 연구에 이용된 용기묘는 생육밀도를 조절할 수 있는 받침(tray, 50 cm × 50 cm)에 25본씩 양묘하였다. 16가지 용기 규격별 각각 12 tray씩, 총 192 tray에 수종별 총 4,800본을 이용하였다. 연구대상 수종의 종자를 2일간 침수 후 파종하였으며, 시험 묘목은 지면에서 70 cm 높이 용기 받침 위에 배치하였다. 피트모스, 펄라이트, 버미큘라이트를 1 : 1 : 1(v/v)로 혼합한 생육상토(Table 2)를 이용하였다. 4월부터 10월까지 매일 스프링클러를 이용하여 m2 당 20 L를 관수하였으며, 또한, 6월부터 3달 동안 수용성 비료인 MultiFeed 19 (19N : 19P2O5 : 19K2O, Haifa Chemicals, Israel)를 1 g L-1 (1,000 ppm) 배액으로 희석하여 주 1회 관수와 병행하여 시비하였다.
| Bulk density (g m-3) | pH | EC (ds m-1) | O.M. (%) | Total N (%) | P2O5 (mg kg-1) | Exch. cations (cmolc kg-1) | C.E.C. (cmolc kg-1) | ||
|---|---|---|---|---|---|---|---|---|---|
| K+ | Ca2+ | Mg2+ | |||||||
| 0.37 | 6.1 | 0.06 | 4.0 | 0.08 | 3.0 | 0.3 | 3.3 | 2.5 | 24.3 |
양묘과정에서 용기 규격에 따른 질소 양분 특성을 분석하기 위해 2013년 10월 용기묘 12본 중 무작위로 4본을 선정하여 뿌리, 줄기, 잎으로 구분하여 각각의 건중량 및 질소(N) 농도를 조사하였다. 용기묘의 질소 농도를 측정하기 위해 부위별로 나누어 105℃에서 16시간 이상 건조시켰다. 건조된 시료를 곱게 갈아 CN 분석기(Variomax CN, ELEMENTAR, Germany)를 이용하여 1,200℃ 고온에서 연소 후 측정하였다(Jones Jr, 1999; RDA, 2000).
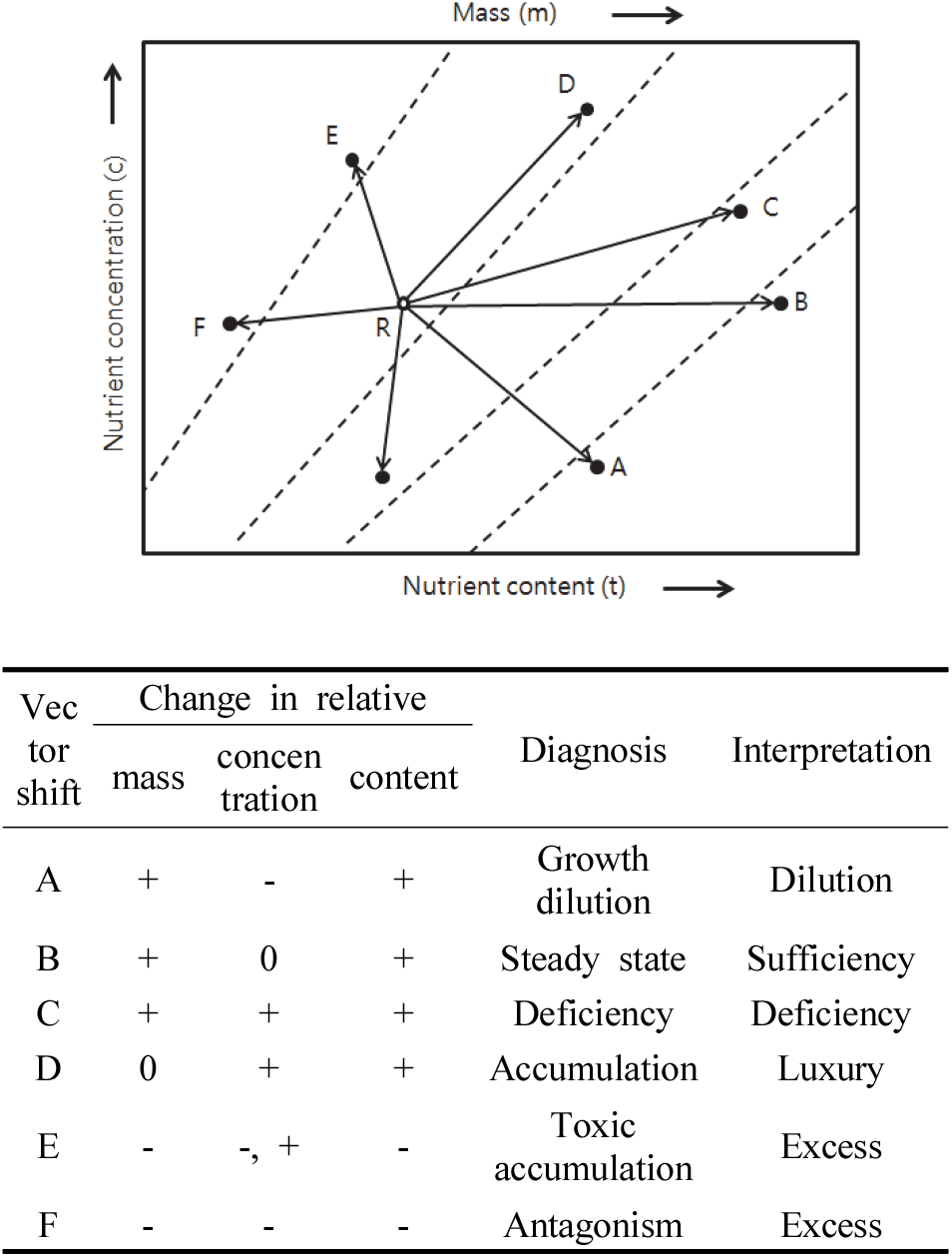
용기 종류에 따른 생장과 질소 양분변화 관계를 구명하기 위해 조사된 묘목의 질소 농도와 함량 및 건중량을 이용하여 벡터분석(Haase and Rose, 1995; Park et al., 2013)을 수행하였다. 벡터분석에서 화살표의 길이는 용기별 질소 양분조건에 따른 묘목의 반응 크기를 나타낸다(Figure 2). 식물체 질소 농도가 식물 생장과 식물체 질소 함량 증가만큼 증가하지 않을 때 “양분결핍”으로 판단하고(벡터 C), 생장과 질소 함량이 증가하는 동안 농도가 일정하게 유지되는 경우 “양분최적”으로 판단하였다(벡터 B). 질소 농도는 증가하지만, 생장 변화가 없는 경우는 질소를 과다하게 소비하는 “양분과다” 상태로 판단하며(벡터 D), 생장이 감소하는 경우는 질소에 의한 “과량집적” 또는 “양분독성” 상태로 판단했다(벡터 E 또는 F). “양분희석”(벡터 A)은 특정 원소의 결핍으로 인해 생장 제한이 다른 원소로 해소될 때 나타나는 것으로 판단했다. 질소 농도 측정에 사용된 용기묘의 뿌리, 줄기, 잎은 2013년 9월 중순 이후 수집된 시료를 이용하였다. 수목은 일반적으로 여름 이후 양분재이동(nutrient retranslocation)이 시작되며, 가을부터는 잎의 양분이 뿌리와 줄기로 활발히 이동하게 된다(Lugo et al., 1990; Salifu and Timmer, 2003a). 이러한 양분재이동에 의한 질소 농도 결과의 오류를 최소화하기 위해 무게가중양분농도(mass-weighted nutrient concentration) 측정 방법을 이용하여 묘목 전체의 질소 농도를 기준으로 분석하였다(Niinemets and Tamm, 2005). 잎에서 재이동된 양분은 줄기와 뿌리에서 측정되기 때문에 묘목 전체의 질소 농도는 양분재이동에 의한 변화를 최소화할 수 있다.
용기 규격이 활엽수 세 수종의 질소 양분 특성에 미치는 영향을 비교․분석하기 위해 SAS System for Windows Version 8.2 (SAS Institute Inc., USA) 프로그램을 이용하여 용기의 생육밀도와 용적에 관한 이원분산분석(Two-way ANOVA)을 실시하였으며, 16가지 용기 종류에 따른 유의성 분석을 위해 Duncan의 다중검정법(Multiple comparison)을 이용하였고, 생육밀도와 용적의 양적 증가 또는 감소에 따른 질소 양분 특성과의 상관관계(Correlation analysis)를 분석하였다. Sigmaplot Version 12.0 (Systat Software Inc., USA) 프로그램을 이용하여 용기의 생육밀도와 용적이 동시에 고려된 용기 규격을 최종적으로 제시하기 위해 질소 양분 함량을 기준으로 다중회귀분석(Multiple regression)을 다음의 식(Gaussian function)으로 실시하였다.
위 식에서 x, y는 각각 용기의 생육밀도와 용적이며, z는 질소 양분 함량을 뜻한다. e는 자연로그상수이며, x0, y0는 z 값이 가장 높을 때의 각각의 생육밀도와 용적이다.
결과 및 고찰
졸참나무는 뿌리, 줄기, 잎 및 묘목 전체의 무게가중질소농도 모두 196본/m2-460 cm3/구 용기에서 가장 높은 값을 보였다. 또한, 용기의 생육밀도와 용적에 따라 각각 유의적 차이를 보였으며, 두 요인에 대한 상호작용 영향이 나타났다. 묘목의 부위별 질소 농도 및 무게가중질소농도 모두 196본/m2 용기에서 유의적으로 가장 높았으며, 용기 용적이 커질수록 질소 농도가 증가하는 경향을 나타냈다(Table 3). 물푸레나무의 경우 잎의 질소 농도는 용기에 따른 통계적 차이가 없었다. 그러나 뿌리, 줄기 및 묘목 전체의 무게가중질소농도는 모두 256본/m2-300 cm3/구 용기에서 가장 높은 값을 보였으며, 용기의 생육밀도와 용적간의 상호작용이 나타났다. 또한, 생육밀도 기준으로 144본/m2 용기가 가장 높은 뿌리 질소 농도를 보였으며, 196본/m2 > 100본/m2 > 256본/m2 순이었다(Table 4). 느티나무는 뿌리, 줄기, 잎의 질소 농도가 가장 높은 용기가 각각 달랐지만, 생육밀도와 용적간의 상호작용은 존재하였다. 묘목 전체의 무게가중질소농도는 196본/m2-380 cm3/구 용기에서 가장 높았으며, 100본/m2-300 cm3/구 용기에서 가장 낮게 나타났다. 생육밀도 기준의 뿌리 질소 농도는 144본/m2 용기에서 가장 높았으며, 나머지 3가지 생육밀도에서는 통계적 차이가 없었다. 또한, 용적이 작을수록 잎의 질소 농도가 감소하는 경향을 나타냈다(Table 5).
일반적으로 식물이 조직 내 필요로 하는 적절한 잎의 질소 농도 수준은 약 1.5%이다(Epstein, 1999). 또한 수종별 차이는 있지만, 양묘과정에서 시비가 계속 증가하더라도 잎의 질소 농도는 약 3.5% 이상 증가하지 않는다(Ingestad, 1979). 본 연구에서도 세 수종 모두 잎의 질소 농도가 약 1.5~3.0% 범위 수준이었으며, 용기에 따른 생육밀도와 용적 차이로 양묘과정에서 질소 농도의 변화를 확인할 수 있었다. 일반적으로 시비 및 관수 효과, 뿌리 공간 확보에 의한 양분 흡수 능력 향상 등의 이유로 생육밀도가 낮고, 용적이 클수록 용기묘의 양분 농도가 높다고 보고되고 있다(Aghai et al., 2014). 그러나 Van den Driessche(1984)는 전나무를 대상으로 생육밀도가 높을수록 양분 농도가 높아지는 반대의 경향을 보고하였으며, 자작나무 용기묘는 용기 규격에 따른 양분 농도의 차이가 없었다는 연구 결과도 있다(Apholo and Rikala, 2003). 본 연구에서도 용기의 생육밀도와 용적의 증가 또는 감소에 따른 변화보다는 두 요인의 복합적인 상호작용 영향이 더 큰 것으로 판단되며, 느티나무와 졸참나무는 물질생산량이 높았던 용기에서 질소 농도가 높아지는 경향을 보였다. 이는 수종별 적정 생육환경에서 활발한 생리 및 생장 활동에 의해 원활한 양분 흡수가 이루어질 수 있었기 때문이다(Rikala, 1989). 높은 양분 농도를 지닌 묘목은 우수한 물질생산량과 함께 결합하여 조림 후에도 높은 활착과 생장을 보인다(Hsu et al., 1996). 이는 묘목의 양분저장능력(nutrient loading)과 관련되어 있으며(Timmer, 1996), 양분재이동으로 줄기와 뿌리로 축적된 양분이 조림 후 활착과 생장에 도움을 주기 때문이다(Nambiar and Fife, 1991). 줄기 질소 농도가 증가할수록 조림 후 생장이 증가하는 정의 상관을 볼 수 있는 것이다(Puertolas et al., 2003). 그러나 2.4% 이상의 잎 내 질소 농도에서는 생존율이 저하된다는 보고도 있으며(Van den Driessche, 1988), 이는 양묘과정에서 임계농도 이상의 시비에 의한 양분의 과량집적 또는 양분독성의 피해가 조림 후에 나타난 것으로 판단된다(Imo and Timmer, 1999). 즉, 수종별 적정 용기 적용에 의한 시비 및 관수 효과 향상과 함께 적정 시비 기술이 동시에 이루어져야 우량 묘목의 생산과 조림 성과를 기대할 수 있는 것이다.
용기 규격에 따른 졸참나무는 196본/m2-380 cm3/구 용기에서 유의적으로 높은 질소 함량을 보였으며, 100본/m2- 300 cm3/구 용기에서 가장 낮았다. 또한, 용기의 생육밀도와 용적 두 요인간 상호작용이 존재하였다. 졸참나무는 m2 당 196본 용기에서 다른 3가지 생육밀도보다 약 2~3배 높은 질소 함량을 보였으며, 용기 용적이 작을수록 질소 함량이 유의하게 감소하는 경향을 나타냈다[Figure 3(A)]. 물푸레나무는 용기의 생육밀도와 용적 두 요인간 상호작용이 있었으며, 100본/m2-380 cm3/구와 256본/m2-220 cm3/구 용기에서 각각 최대와 최소 질소 함량을 나타냈다. 느티나무와 반대로 100본/m2 용기에서 가장 높은 질소 함량을 보이면서, 생육밀도가 낮을수록 질소 함량이 유의적으로 증가하는 경향을 나타냈다. 또한, 용기 용적 기준으로 220 cm3/구 용기에서 가장 낮은 질소 함량을 보였으며, 나머지 3가지 용적은 통계적 차이가 없었다[Figure 3(B)]. 느티나무의 질소 함량은 196본/m2-380 cm3/구 용기에서 가장 높았으며, 반면 100본/m2-300 cm3/구 용기에서 가장 낮았다. 생육밀도와 용적 각각에 따라 유의적 차이가 나타났으며, 두 요인에 대한 상호작용이 존재하였다. 생육밀도 기준으로 100본/m2 용기에서 가장 낮은 질소 함량을 보였으며, 나머지 3가지 용기에서는 통계적 차이가 없었다. 또한, 380 cm3/구와 460 cm3/구 용기에서 가장 높은 질소 함량을 보이면서 용적이 클수록 질소 함량이 증가하는 경향을 나타냈다[Figure 3(C)].
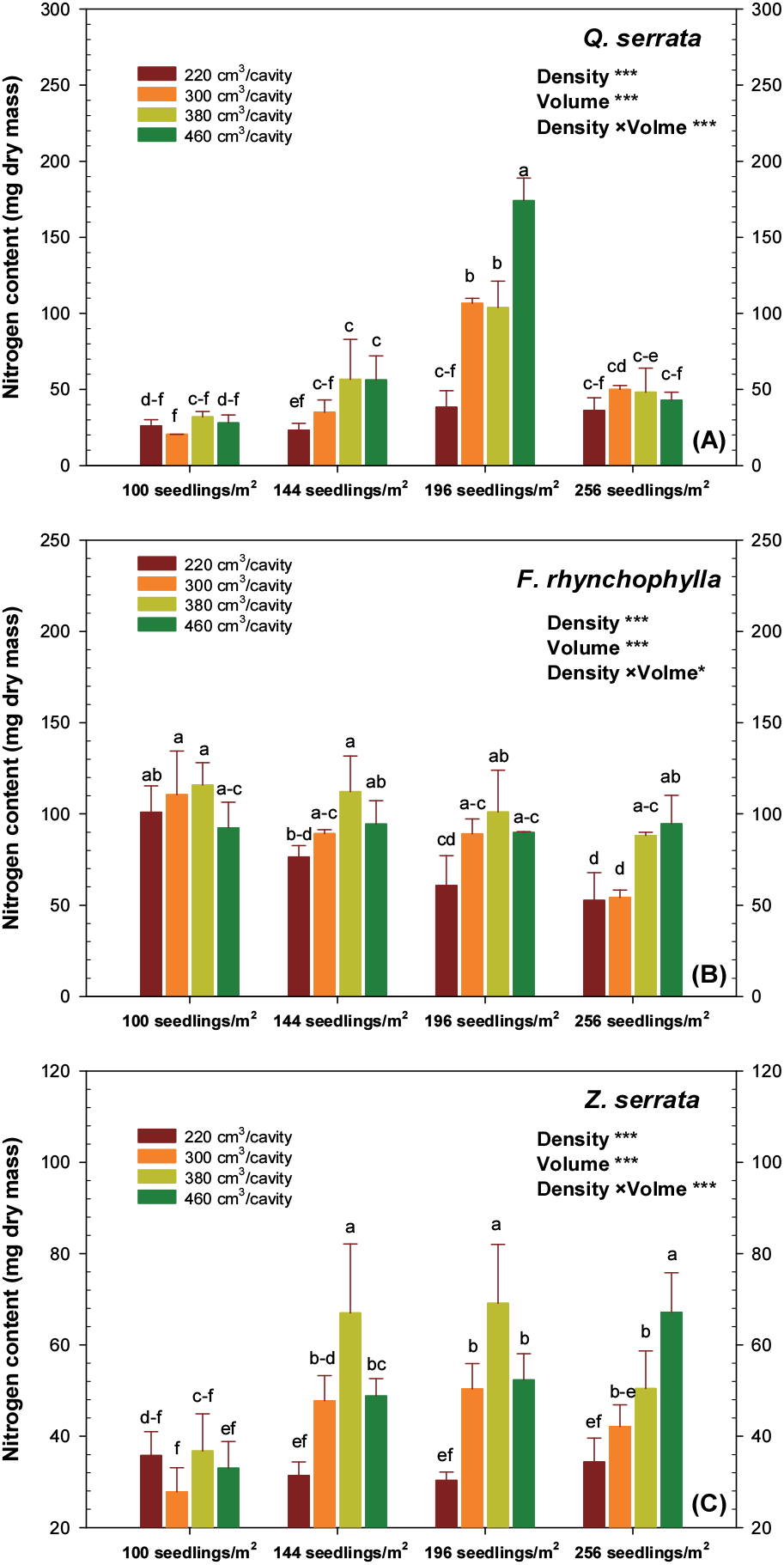
수목의 양분 농도는 생장 및 생리 활성의 잠재력이라 볼 수 있으며(Timmer, 1996), 양분 함량은 이러한 잠재력에 양적인 개념이 더해진 것이라 할 수 있다. 즉, 묘목의 외형적 크기 및 물질생산량과 연계하여 양분 특성의 최종 산물인 양분저장능력으로 표현되는 것이다(Way et al., 2007). 이러한 양분 함량은 농도와 함께 조림 후 생장을 예측하는 지표로 활용되고 있다(Haase et al., 2006). Switzer and Nelson(1963)은 양묘과정에서 Pinus taeda 묘목의 질소 함량과 조림 후 생장은 정의 상관이 84%였다고 보고하였다. 본 연구에서도 질소 농도 연구 결과와 같이 용기 규격에 따른 묘목의 질소 함량 변화를 관찰할 수 있었다. 또한 느티나무와 졸참나무의 경우 용적이 220 cm3인 용기는 생육밀도와 관계없이 다른 3가지 용기 용적보다 낮은 질소 함량을 보였다. 이러한 이유는 좁은 용기 구 직경과 작은 지하부 생육공간에 의한 뿌리의 생육 부진으로 양분이용효율의 저하가 나타나기 때문이다(Tsakaldimi1 et al., 2005).
물푸레나무는 질소 농도와 용기의 규격과는 상관이 없었다. 졸참나무 잎의 질소 농도는 생육밀도와 용적 모두 정의 상관을 보였으며, 줄기와 용적만 정의 상관이 나타났다. 느티나무는 용기의 생육밀도와 잎의 질소 농도만 정의 상관관계를 보였다. 졸참나무의 질소 함량은 생육밀도와 상관이 없었으며, 물푸레나무의 질소 함량은 생육밀도와 부의 상관을 나타냈다. 느티나무의 질소 함량은 생육밀도 와 정의 상관을 보였다. 또한, 묘목의 질소 함량은 세 수종 모두 용적과 정의 상관을 나타냈다(Table 6). 이는 Pinus contorta 묘목의 질소 함량이 용기 용적, 높이 및 직경과는 정의 상관을 보인 기존 연구 결과와 같은 경향을 나타낸 것이다(Van den Driessche, 1984). 양분 특성과 생장은 정의 상관관계를 가진다고 보고되고 있으며(Paterson, 1996), 양묘과정에서의 높은 양분 함량 결과는 조림 후 우수한 묘목품질을 기대할 수 있다.
양분벡터분석은 시비와 관련된 연구에 대부분 적용되고 있으나 본 연구는 용기의 생육밀도와 용적에 따른 시비 및 양분저장능력을 알아보기 위해 실시하였기 때문에 대조구를 16가지 실험대상 용기 중 시비 효과와 양분저장능력이 가장 낮을 것으로 예상되는 256본/m2-220 cm3/구 용기를 기준으로 분석하였다.
졸참나무는 생육밀도가 낮고, 용적이 큰 100본/m2-380 cm3/구, 100본/m2-460 cm3/구 용기에서 ‘양분최적’ 상태를 보였으며, 약 20% 이상 높은 생장을 나타냈다. 생육밀도가 m2 당 196본인 용기들은 용적이 220 cm3인 용기를 제외하고 모두 ‘양분결핍’ 상태를 보였다. Salifu and Jacobs (2006)도 Quercus rubra 노지묘를 대상으로 질소 기준의 양분벡터분석을 통해 무시비 처리구는 ‘양분결핍’ 상태였으나, 시비 처리에 의해 ‘양분최적’ 상태로 변화되었다고 보고하였다. ‘양분결핍’ 상태의 용기는 본 연구에서 조사된 졸참나무 적정 생육밀도와 같은 수준이며, 묘목품질과 양분저장능력의 향상을 위해 시비량 증가가 가능할 것이다. ‘양분과다’ 현상이 196본/m2-220 cm3/구 용기에서 나타났으며, 100본/m2-300 cm3/구와 144본/m2-220 cm3/구 용기에서는 생장이 감소하는 ‘양분독성’ 상태로 판단된다[Figure 4(A)]. 생육밀도가 낮고, 용적이 작은 용기는 시비 효과가 보통 이상이지만, 용적이 작아 축적된 양분을 생장 활동에 이용하지 못해 묘목이 피해를 입을 수 있는 것이다. 노지묘 생산에서도 과량 시비에 의해 토양 산도 증가, 알루미늄 독성 등에 의한 생장 감소가 보고되고 있다(Teng and Timmer, 1995). 물푸레나무는 100본/m2-300 cm3/구 용기에서 약 80% 이상 높은 생장을 보이면서 ‘양분최적’ 상태를 보였다. 그러나 생육밀도가 높고, 용적이 작은 196본/m2-220 cm3/구, 256/m2-300 cm3/구 용기에서는 생장이 감소하는 ‘과량집적’ 상태가 나타났다. 또한 질소 농도는 증가하지만, 생장의 변화가 없는 ‘양분과다’ 상태가 144본/m2-220 cm3/구 용기에서 보였으며, 그 외 용기 규격에서는 대부분 ‘양분결핍’ 상태였다[Figure 4(B)]. 이는 물푸레나무가 느티나무와 졸참나무와 달리 상대적으로 잎 면적이 넓고, 지상부가 펼쳐져 있는 생장 형태이기 때문에 시비 및 관수 차단 효과가 높았기 때문으로 판단된다. 물푸레나무는 시비 회수의 증가 또는 시비 방법의 개선 등을 통해 생장을 증가시킬 수 있으며, Fraxinus rhynchophylla 노지묘를 대상으로 양분벡터분석을 통해 ‘양분결핍’ 상태에 따른 질소 시비량 증가를 제언한 연구(Park et al., 2013)와 같이 해석할 수 있다. 느티나무의 경우 144본/m2-300 cm3/구 용기가 대조구보다 약 30% 이상 높은 건중량을 보였으며, 일정한 질소 농도에서 생장과 질소 함량이 증가하는 ‘양분최적’ 상태였다. 용기별 정도의 차이는 있지만 144본/m2-380 cm3/구, 196본/m2-300 cm3/구, 196본/m2-380 cm3/구, 256본/m2-460 cm3/구 용기는 질소 농도가 묘목의 생장과 질소 함량 증가만큼 증가하지 않은 ‘양분결핍’ 상태이었으며, 대부분 생육밀도가 높은 용기이다. 144본/m2-220 cm3/구 용기는 생장이 감소하는 ‘과량집적’ 상태였으며, 나머지 대부분의 용기는 ‘양분희석’ 상태의 결과를 보였다[Figure 4(C)]. 본 연구결과 느티나무는 m2 당 약 180~220본의 생육밀도가 적정한 것으로 조사되었다. 그러나 수종이 요구하는 적정 생육밀도와 달리 시비효율은 생육밀도가 낮을수록 향상되기 때문에 생장이 우수한 용기에서 ‘양분결핍’ 상태가 나타났으며, 집중적인 시비 관리가 있을 시 묘목품질과 양분저장능력이 더욱 증가할 수 있을 것으로 판단된다.
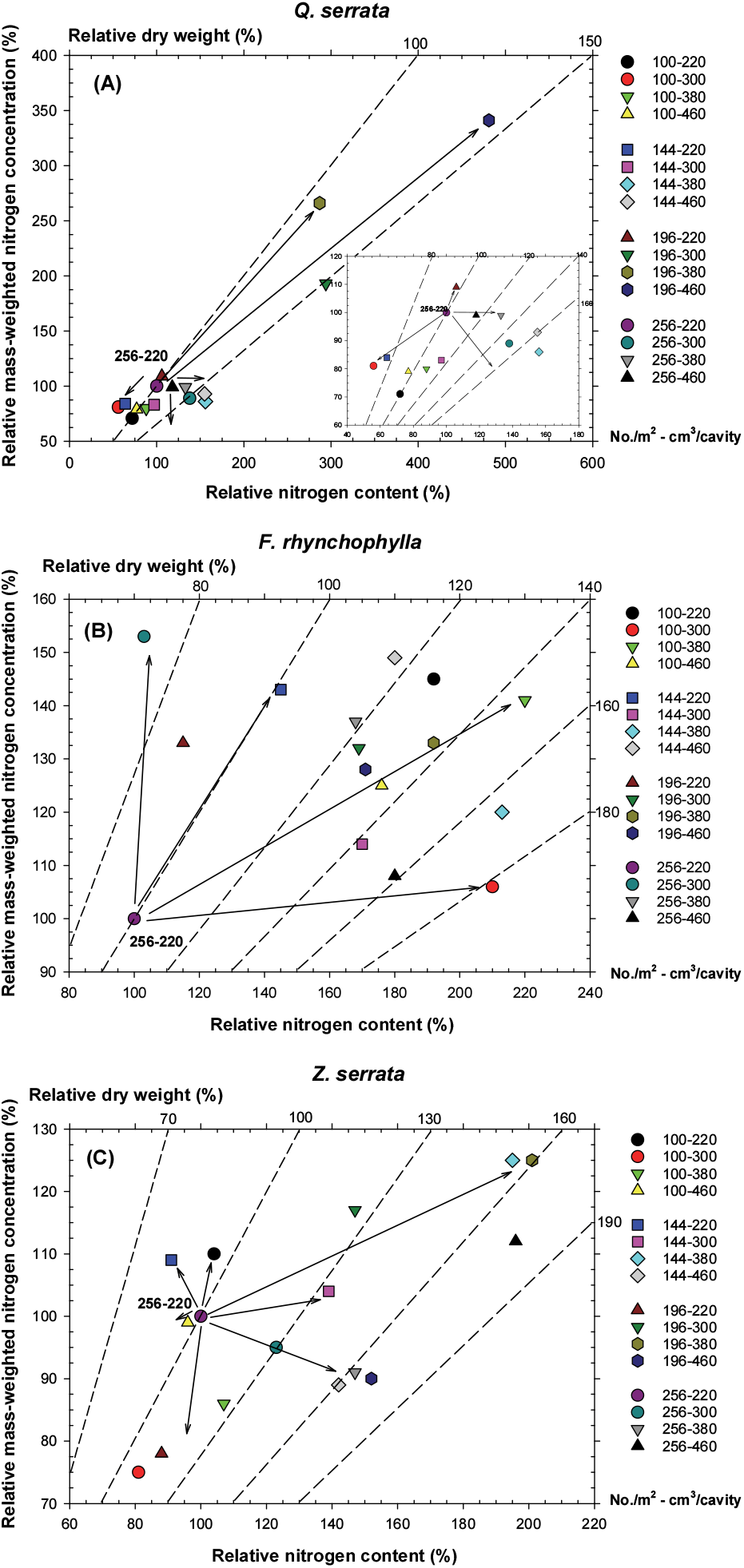
양분벡터분석은 묘목의 생장, 질소 농도의 변화, 묘목이 흡수한 질소 함량의 변화를 동시에 연계하여 분석할 수 있으며, 시비 효과에 따른 생장과 양분 특성 관계를 추적할 수 있다. 본 연구에서도 용기의 생육밀도와 용적에 따른 시비 효과 차이로 수종별 묘목품질과 양분저장능력이 다르게 나타났으며, 수종별 용기 규격의 양분 상태를 판단하여 시비량 증가 또는 감소를 제시할 수 있다. 양분벡터분석을 실시하여 수종별 최적 양분 상태의 용기를 알 수 있었다. 또한 수종별 적정 용기 규격에서 양분 상태와 관련하여 문제가 있을 경우 시비 기술 개선을 통해 최적의 양분저장능력의 묘목을 생산할 수 있을 것으로 판단된다. Salifu et al.(2009)는 Quercus rubra와 Q. alba 노지묘를 대상으로 실시한 연구에서 양분 농도는 조림 후 생장과 정의 상관을 가진다고 보고하였다. 양묘과정에서 묘목의 양분저장능력 향상 또는 양분 함량 증가는 생장에 적은 영향을 미치지만, 조림 후 생장 및 활착에 점진적으로 효과를 주어 조림 성과가 향상된다(Benzian et al., 1974; Margolis and Waring, 1986). 특히, 양분저장능력은 하층식생과의 경쟁이 심한 임분, 건조지, 척악지 등 입지 환경이 불량한 조림지에서 능력을 더욱 발휘할 수 있는 것이다(Malik and Timmer, 1995).
양묘과정에서 용기의 생육밀도와 용적의 상호작용이 확인된 묘목의 질소 함량을 기준으로 수종별 용기의 두 요인이 동시에 고려된 적정 용기 규격을 제시하고자 다중회귀분석을 Figure 5와 같이 실시하였다(Dominguez-Lerena et al., 2006). 다중회귀분석을 통해 도출된 함수식을 이용하여 상위 10%의 용기 규격을 기준으로 생육밀도와 용적의 최소(이상)와 최대(이하) 범위를 제시하였다.
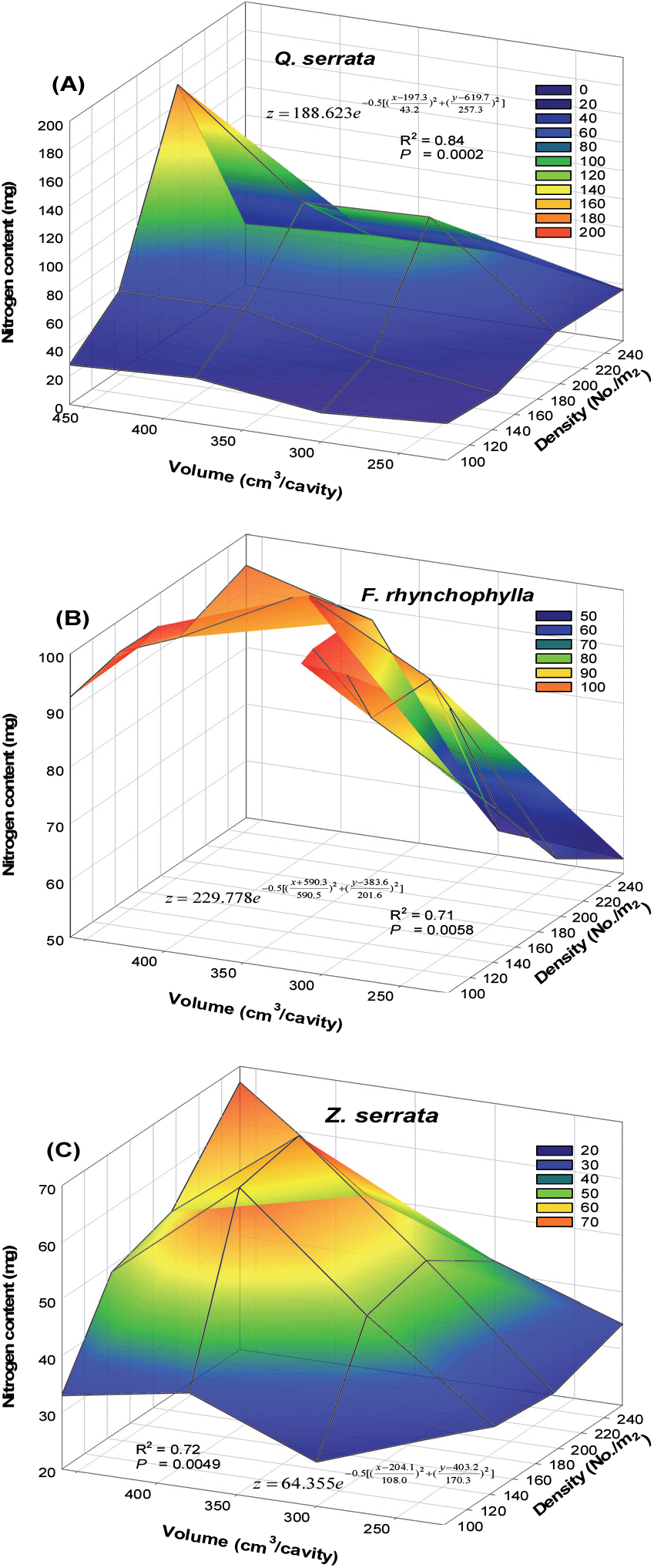
수종별 양분저장능력을 판단할 수 있는 질소 함량이 우수한 용기 규격은 졸참나무, 물푸레나무, 느티나무 각각 180~210본/m2과 410~460 cm3/구, 100~120본/m2과 350~420 cm3/구, 190~220본/m2과 380~430 cm3/구 범위였다(Table 7).
| Species | Density (No./m2) | Volume (cm3/cavity) | ||
|---|---|---|---|---|
| Min. | Max. | Min. | Max. | |
| Q. serrata | 180 | 210 | 410 | 460 |
| F. rhynchophylla | 100 | 120 | 350 | 420 |
| Z. serrata | 190 | 220 | 380 | 430 |
느티나무, 물푸레나무, 졸참나무는 시설양묘과정에서 용기의 생육밀도와 용적 차이 및 두 요인의 상호작용에 따른 무기환경과 양분 조건 등의 변화된 생육환경이 수종별로 각기 다른 질소 양분 특성 차이가 나타났다. 또한, 수종별 양분 요구도와 연계된 생리 및 형태학적 특성에 따라 적정 용기 규격에 차이가 있었다. 추가적으로 양분벡터분석을 실시하여, 용기의 생육밀도와 용적에 따른 시비 효과 차이로 수종별 묘목품질과 양분저장능력이 다르다는 것을 확인할 수 있었다. 이는 수종별 용기 규격에 따른 묘목의 ‘양분결핍’과 ‘과량집적’ 등의 양분 상태를 판단하여 이에 맞는 시비 기술 개선을 통해 최적의 양분저장능력을 가지는 묘목을 생산할 수 있을 것으로 기대된다.
결 론
본 연구는 졸참나무, 물푸레나무, 느티나무를 대상으로 용기 규격이 시설양묘 과정에 질소 양분 특성에 미치는 영향을 구명하기 위해 수행되었다. 16 종류의 용기에서 생산된 묘목의 질소 양분 특성을 조사·분석하였다. 생육밀도와 용적에 따른 용기묘의 질소 양분 농도 및 함량 변화를 알아보기 위해 이원분산분석 및 다중회귀분석을 이용하였다.
세 수종 모두 용기 규격에 따라 묘목의 질소 양분 특성은 유의적 차이를 보였으며, 생육밀도와 용적 두 요인간의 상호작용이 질소 농도 및 함량에서 확인되었다. 세 수종 모두 질소 함량과 용적은 정의 상관관계를 보였지만, 물푸레나무의 질소 함량은 생육밀도와 부의 상관관계를 나타냈다. 추가적으로 양분벡터분석을 실시하여, 용기 규격에 따른 시비 효과 차이로 양분결핍, 과량집적 등의 양분저장능력이 다르다는 것을 확인하였다.
묘목 질소 함량을 기준으로 다중회귀분석기법을 적용한 결과, 졸참나무는 180~210본/m2과 410~460 cm3/구, 물푸레나무는 100~120본/m2과 350~420 cm3/구, 느티나무는 190~220본/m2과 380~430 cm3/구가 최적 용기 규격으로 최종 도출되었다. 수종별 적정 용기 적용으로 시설양묘 과정에서 양분저장능력이 높은 묘목 생산과 함께 조림 후에도 우수한 생장을 기대할 수 있을 것이다.