



서 론
생물다양성 위기가 심각해지며 생물다양성 보전과 산림 복원이 주요 화두가 되고, 자생수종을 이용한 조림과 산림 복원이 시도되고 있다(Millet et al., 2013; Svejcar et al., 2017). 훼손이 심하여 자생수종의 즉각적인 도입이 어려운 지역에서는 외래수종으로 조림지를 조성하여 토양을 안정시켜 자생식물이 이입되기 좋은 환경을 유도하기도 한다(Lugo, 1997). 시간이 지남에 따라 훼손 조림지의 환경이 개선되면 주변의 천연림에서 수종이 침입하여 주변 천연임분과 종 조성 및 종 다양성이 비슷해진다는 결과가 여러 지역에서 보고되었다(Sonali, 2001; Howard and Lee, 2003; Nagaike et al., 2003; Godefroid et al., 2005; Nagaike et al., 2006). 조림지는 자생종 복원의 촉매 역할을 할 수 있어 조림지 내 자생수종의 천연갱신을 통해 임분을 복원하고 생물다양성을 보전하고자 하는 연구가 다양한 지역에서 이루어지고 있다(Parrota, 1995; Geldenhuys, 1997; Keenan et al., 1997; Kanowski et al., 2005; Gomez-Aparicio et al., 2009; Kim et al., 2013).
우리나라는 국토 녹화의 목적으로 1960-1980년대 대규모의 조림을 실시하였으나, 당시 척박한 토양이나 묘목 수급 등의 이유로 조림 수종의 대다수는 리기다소나무(Pinus rigida Mill.), 아까시나무(Robinia pseudoacacia L.), 일본잎갈나무(Larix kaempferi (Lamb.) Carrière) 등 외래수종이었다(Kim and Zsuffa, 1994). 토양이 개선되며 우리나라의 산림도 황폐지 복구에서 생물다양성을 고려한 산림 복원을 시도할 수 있는 환경을 갖추게 되었다. 외래 수종 조림지에서 자생수종 침입을 이용한 임분 복원 과정은 천연갱신에 의한 우점종의 점차적인 변화라는 점에서 천이 과정과 비슷하며, 식재나 직파에 의해 단기간에 이루어지는 인공갱신과 차별화된다(Donato et al., 2012). 이러한 방식의 복원을 시도하기 위해서는 하층에서 버틸 수 있는 내음성 수종이 있어야 하고, 내음성 수종의 종자를 공급해주는 모수가 주변에 존재해야 한다(Oliver and Larson, 1996; Jang and Park 2010).
전나무속(Abies)은 대부분 내음성 수종으로 천이 후기 단계 또는 노령림의 주요 수종으로 알려져 있다(Zobel and Antos, 1991; Ishikawa et al., 1999; Kim et al., 2005; Taylor et al., 2006). 전나무(A. holophylla Maxim.)는 우리나라 자생수종 중 대표적인 내음성 수종이며 비교적 천연갱신이 잘 되는 수종으로 알려져 있다(Lee, 1999). 전나무는 극동러시아, 중국 북부와 북동부, 한반도에 분포한다(Boldeskul et al., 2015). 한반도 내 전나무는 남쪽으로는 제주도부터 북쪽의 백두산까지 전국에 생육하며 우리나라 온대북부림과 아한대림을 연결하는 수종이다. 한반도 내에 분포하는 전나무속의 다른 수종과 비교했을 때 전나무는 너덜 지역을 비롯한 계곡부 등 습윤한 장소에 다양하게 정착하여 자라며 분비나무(A. nephrolepis (Trautv.) Maxim.)보다 낮은 고도에 분포한다(Lim, 1985; Korea National Arboretum, 2015). 국내에서 전나무림은 광릉부터 대관령, 오대산, 내설악까지 주로 중부 이북 지역에 분포하고 있으며, 남부지역에서는 변산반도 내소사 지역과 수도암 지역의 전나무림이 알려져 있다. 천연 전나무림의 분포는 오대산 월정사 지역에서 600-1200 m (Nam et al., 2000; Kim et al., 2015; Han et al. 2015), 내설악산 길골 지역 600-900 m (Yoon, 2009), 수도암 지역 800-1000 m (Choi and Lee, 2014) 등 해발 500 m 이상 지역에서 주로 보고되어 있으나, 내소사 지역처럼 해발 150 m 미만의 낮은 지역에서 나타나기도 한다.
전나무속은 생장이 느리고 내구성이 약하여 목재 생산 등 상업적으로 선호되는 수종은 아니었다(Quiring et al., 2008; Boucher et al., 2009). 전나무속에 대한 연구는 조림지에서의 생산보다는 대부분 내음성이나 천연림에서의 생태적 역할을 중심으로 이루어졌다(Diaci, 2002; Nakagawa et al., 2003). 전나무에 대한 국내 연구도 천연림의 주요 구성 수종으로서 임분 내 전나무의 분포, 전나무림의 군집 구조나 천이 경향에 대한 연구가 주를 이룬다. 천연 전나무림은 역J자형 직경분포를 보이며 하층의 전나무는 내음성이 있어 전생치수로 생장에 유리한 환경을 기다리는 갱신 전략을 유지한다(Chung, 1996). 이러한 갱신 전략 덕에 전나무림은 자체 갱신에 의하여 군집이 유지되고(Park and Jeon, 2010), 다양한 세대로 이루어진 개체군을 유지하며 넓은 수령분포를 가진다(Choi and Lee, 2014). 전나무는 강원도 지역 침활혼효림에서 신갈나무(Quercus mongolica Fisch. ex Ledeb.), 피나무(Tilia amurensis Rupr.), 고로쇠나무(Acer pictum var. mono Maxim. ex Franch.), 잣나무(Pinus koraiensis Siebold & Zucc.) 등과 함께 분포하며(Kim et al., 2015; Han et al. 2015), 극동러시아 지역 침활혼효림에서는 가문비나무(Picea jezoensis (Siebold & Zucc.) Carrière), 잣나무, 분비나무, 피나무, 고로쇠나무 등과 함께 나타난다(Ishikawa et al., 1999).
경기도는 잣나무, 리기다소나무, 일본잎갈나무 조림지가 전체 산림의 약 30%를 차지하고 있으며, 현재 가장 많이 조림되고 있는 수종은 일본잎갈나무와 잣나무이다(Korea Forest Service, 2020). 경기도 가평군은 특히 잣나무 조림지의 비율이 높은 지역으로 총 산림 면적 약 6만 9천 ha 중 2만 ha가 잣나무 조림지이다(Gapyeong-gun Office, 2013). 한편, 경기도 가평 지역의 자생수종인 전나무는 임분이 산재하고 있으나, 구체적인 현황은 알려진 바가 드물다. 가평군 이화리와 산유리에서 발견된 전나무 임분은 과거 조림지 조성 시 남겨놓은 수림대로 추정되며 기존 천연임분에 대한 정보를 제공하였다. 또한 전나무 임분 주변에는 일본잎갈나무, 잣나무, 밤나무(Castanea crenata Siebold & Zucc.) 조림지가 분포하고 있어, 조림지와 주변 전나무 천연임분 간의 상호작용을 보여줄 수 있을 것으로 예상하였다.
본 연구는 경기도 가평 지역에서 전나무가 분포하는 임분을 대상으로 전나무 임분이 잣나무나 일본잎갈나무와 같은 조림 수종을 식재한 조림지와 인접하거나 임분 내 전나무 모수가 분포할 때 임분의 천이 과정을 이해하고자 수행하였다. 또한 이를 바탕으로 조림지에서 지역 자생수종인 전나무로 이루어진 천연림으로의 복원 가능성을 파악하고자 하였다.
재료 및 방법
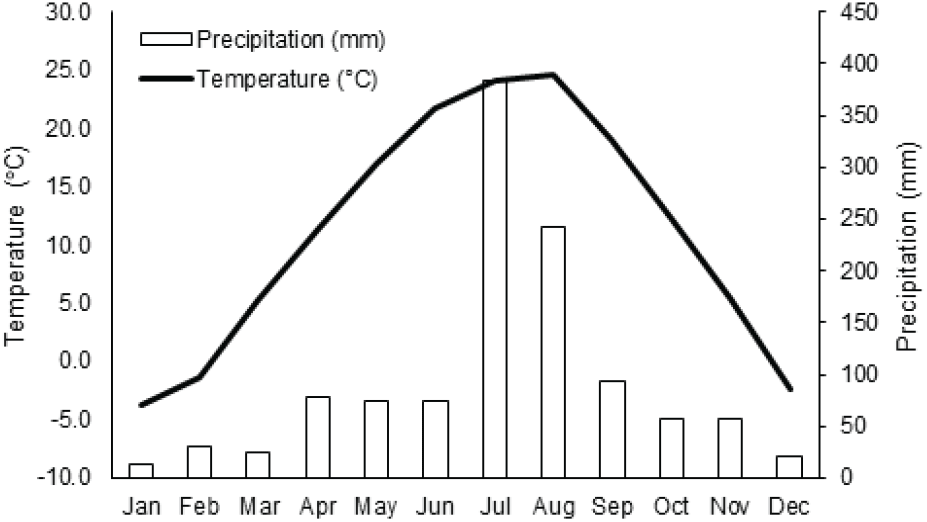
본 연구는 경기도 가평군 가평읍 이화리(37°47’28"-30"N, 127°30’45"-53"E)와 가평읍 산유리(37°45’6"-12"N, 127°30’ 22"-26"E) 지역의 전나무가 분포하는 임분에서 이루어졌다(Figure 1). 조사가 이루어진 가평 지역은 온대 중부 지역의 기후를 가진다. 가장 인접한 기상관측 지점인 남이섬 지점(37°47’23.2"N, 127°31’39.7"E)에서 관측한 자료에 의하면 관측을 시작한 2011년 6월 이후부터 2020년 10월까지 지역의 연평균 기온은 11.09℃, 연평균 강수량은 1150 mm, 평균 풍속은 0.9 m/s를 기록했다. 월별 평균 기온이 가장 높은 달은 8월로 평균 24.6℃를 기록했으며 월별 평균 기온이 가장 낮은 달은 1월로 평균 영하 3.8℃인 것으로 나타났다(Figure 2). 관측이 시작된 이후로 가장 낮은 일평균기온은 -15.8℃, 가장 높은 일평균기온은 31.1℃였으며, 최저 기온은 -22.6℃, 최고 기온은 39.3℃였다. 또한 최고 일강수량은 353.5 mm로 기록되었다(Korea Meteorological Administration, 2020).
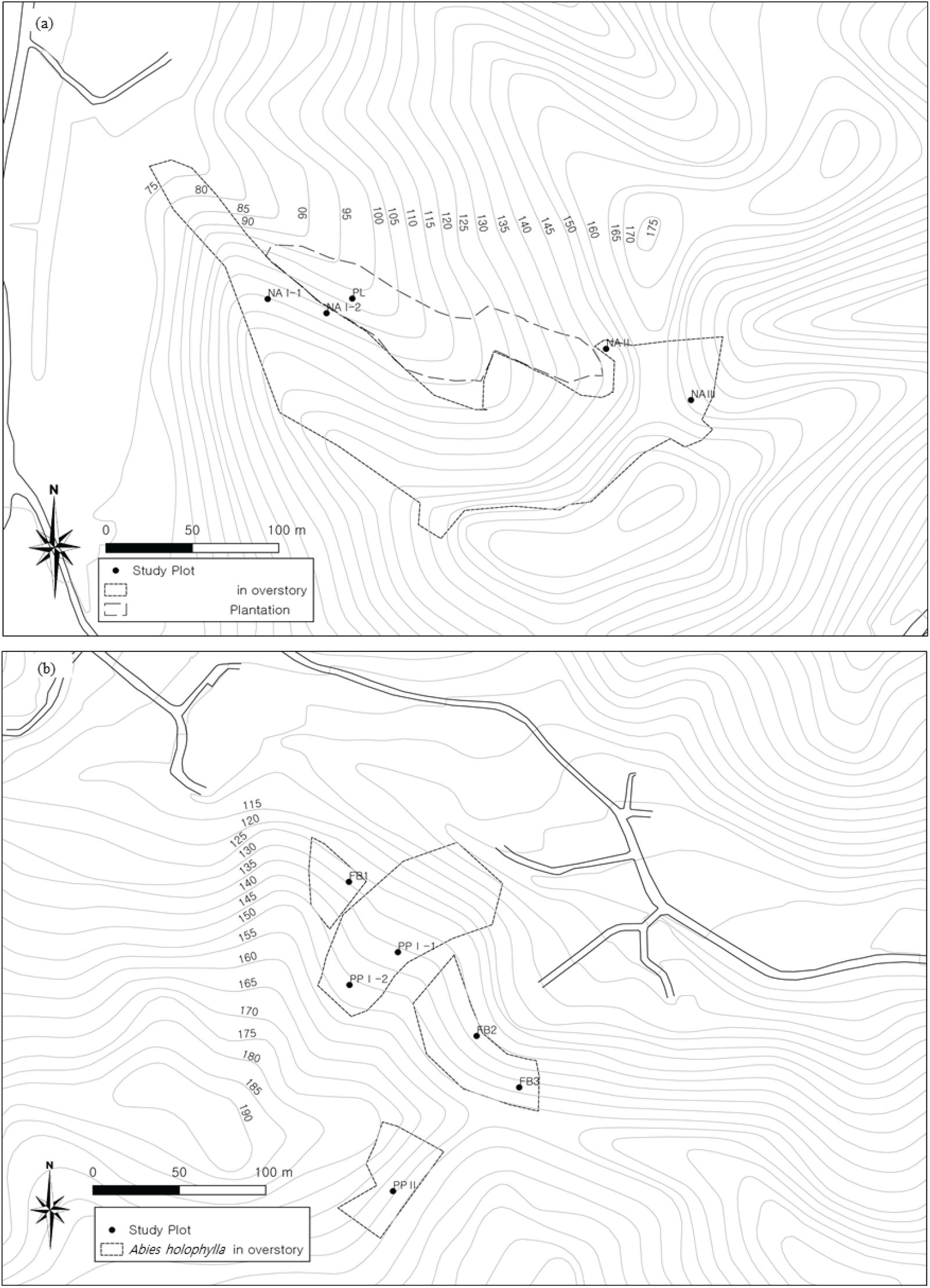
조사지는 해발고도 80-180 m에 위치하고 있으며, 향은 주로 북사면이고, 사면 경사는 11-35%였다. 이화리에 위치한 조사지의 토심은 40-92 cm, 산유리 조사지는 30-90 cm의 범위였다. 이화리 지역의 토양은 오산통(토양형 coarsy loamy, mixed, mesic family of typical Dystrudepts)으로, 표토의 토성은 양토이며 심토의 토성은 사양질이다. 산유리 조사지는 이화리 지역과 동일한 오산통 토양과 청산통 토양(토양형 loamy skeletal, mixed, mesic family of Typic Dystrudepts)이 나타나고 있었다. 청산통은 표토는 양토이며 심토의 토성은 식양질이다(National Institute of Agricultural Sciences, 2020).
조사 지역은 면적은 넓지 않으나 상층을 구성하는 수종이 확연히 다른 임분들이 인접하여 위치하고 있었다. 이화리의 조사 지역은 조림 작업 시 남겨놓은 수림대로 추정되는 전나무가 상층을 우점하는 전나무 임분(NA) 약 2.1 ha와 일본잎갈나무 조림지(PL) 약 0.6 ha로 이루어져 있었다. 산유리의 연구지는 숲가꾸기의 작업 강도, 사면향 등이 차이가 나는 잣나무 조림지 PP I (0.6 ha), PP II (0.2 ha)와 숲가꾸기 작업의 흔적이 있는 수림대(FB, 약 0.4 ha)로 이루어져 있었다(Table 1). 산유리의 잣나무 조림지는 전나무가 상층의 일부를 차지하고 있었다.
NA: Abies holophylla natural stand, PL: Larix kaempferi plantation, FB: Forest belt, and PP: Pinus koraiensis plantation with thinning.
가평군청에 확인한 결과 이화리 조사 지역 일부에서는 1960년 밤나무와 일본잎갈나무를, 1987년 잣나무를 식재하였으며(가평군청 정보공개청구, 2021. 4. 6), 산유리 조사 지역에서는 1974년 잣나무 묘목을 식재한 것으로 확인되었다(가평군청 전화 문의, 2020. 11. 17). 반면 가평읍 이화리와 산유리 지역 두 곳 모두 1958년 이후 전나무 식재 기록이 없었다. 이화리 조사 지역에서는 숲가꾸기 작업 기록이 없어, 일본잎갈나무 조림지에는 식재 이후 특별한 관리는 없었던 것으로 보인다. 산유리 조사 지역에서는 2004년 덩굴제거 작업을 실시한 기록이 있었으며(정보공개청구, 2021. 4. 6), 간벌을 실시한 흔적을 확인하였다.
가평군 가평읍 이화리, 산유리의 전나무가 분포하는 임분에 10 m×10 m 또는 20 m×20 m 크기의 방형구를 설치하고 매목조사를 실시하였다. 조사를 위해 이화리 전나무 임분은 농가와의 거리, 사면향 등을 고려하여 NA I, NA II, NA Ⅲ로 나누고 각각 방형구를 2, 1, 1개 설치하였고, 일본잎갈나무 조림지(PL)에도 방형구 1개를 설치하였다. 산유리 지역에서는 잣나무 조림지 PP Ⅰ에 2개, PP Ⅱ에 1개, 수림대인 FB에 3개의 방형구를 설치하였다. 일부 임분은 면적이 넓지 않아 방형구를 1개만 설치하였다.
방형구 내에 있는 직경 2 cm 이상의 모든 임목의 흉고직경(지상부 1.3 m 높이)을 측정하였다. 전나무, 잣나무, 밤나무, 참나무류를 제외한 수종은 기타 수종으로 기록하였다. 직경급 2 cm 미만 또는 수고 1.3 m 미만의 전나무는 하층의 전나무로 취급하여 각 임분의 방형구 내의 하층 전나무 밀도를 기록하였다. 수종 구성을 알아보기 위 하여 이화리 전나무 임분(NA I), 산유리의 수림대(FB)와 잣나무 조림지(PP I)에 1개의 방형구를 선정하여 방형구 내 흉고직경 2 cm 이상의 수종을 모두 기록하였다.
이화리와 산유리 조사지역의 임분 하층에서 28본의 전나무를 임의로 선정하고, 마디 생장 횟수로 하층 전나무의 수령을 추정하였다. 수령은 전나무가 고정생장을 하는 점을 고려하여 마디 하나를 1년 생장으로 간주하였다. 전나무 성목의 수령을 파악하기 위하여 생장추를 이용하여 이화리의 전나무 6본의 목편 시료를 1.2 m 높이에서 채취하고, 채취한 목편의 나이테를 세어 수령을 추정하였다.
각 방형구에 대해 해발 고도, 방위 및 지리정보위치(GPS 좌표)를 측정하였고, QGIS에서 수치표고모델(DEM)을 이용해 경사도를 측정하였다. 조사 지역의 식재 기록을 확인하기 위해 가평군청 산림조성과에 전화(2020. 11. 27)와 인터넷 정보공개청구(2021. 4. 6.)로 문의하여 1958년 이후의 식재 기록과 숲가꾸기 내역을 수집하였다.
수종 구성과 전나무의 우점도를 알아보기 위하여 임분 내 수종의 상대밀도(RD), 상대피도(RC)를 구하고 상대우점치(IP)를 산출하였다(Brower and Zar, 1977).
상대밀도는 직경 2 cm 이상의 임목을 대상으로 하였으며 피도는 흉고단면적을 이용하여 계산하였다. 모든 조사지를 대상으로 흉고직경을 5 cm 단위로 구분하여 각 직경급에 속하는 ha당 임목 밀도를 계산하였다.
전나무 성목의 밀도와 치수 발생의 관계를 알아보기 위해 임분의 흉고단면적, 전나무의 흉고단면적, 흉고직경 20 cm 이상의 전나무 성목의 흉고단면적과 하층 전나무 밀도와의 상관관계를 스피어만 순위상관계수를 이용하여 분석하였다. 전나무의 임분 내 분포 특성을 알아보기 위해 비계량 다차원척도법(Non-metric multidimensional scaling, NMDS)과 군집분석을 실시하였다. NMDS는 각 직경급의 ha당 밀도를 이용하였으며, NMDS를 통해 나타난 Bray-Curtis distance를 군집 간의 거리값으로 두고 평균기준법을 이용해 군집분석을 실시하였다. NMDS와 군집분석에는 R Studio 3.6.2를 이용하였으며 NMDS는 vegan 패키지의 metaMDS 함수를 이용하였다. 일반 통계처리는 Microsoft Excel 2016(Microsoft, 2016)을 이용하였다.
결과 및 고찰
이화리 조사지는 교목성 수종으로 전나무, 일본잎갈나무, 잣나무, 졸참나무(Quercus serrata Murray), 밤나무 등이 분포했고, 관목성 수종으로는 생강나무(Lindera obtusiloba Blume), 갈매나무(Rhamnus davurica Pall.), 노린재나무(Symplocos sawafutagi Nagam.) 등이 분포하고 있었다. 산유리 조사지에서는 교목성 수종으로 전나무, 잣나무, 갈참나무(Q. aliena Blume), 신갈나무, 밤나무 등이 분포하였고, 하층에는 산뽕나무(Morus australis Poir.), 노린재나무, 생강나무, 붉나무(Rhus chinensis Mill.) 등이 자라고 있었다(Table 2).
전나무의 우점치는 36.1-79.1%의 범위를 보였다(Table 3). 이화리 지역의 전나무 임분에서 전나무의 우점치는 40.4%-79.1%였으며, 일본잎갈나무 조림지(PL)에선 일본잎갈나무의 우점치가 43.0%로 전나무의 우점치(46.2%)보다 다소 낮게 나타났다. 산유리 지역의 수림대 FB에서 전나무 우점치는 58.2%, 잣나무 조림지 PP Ⅰ에서 전나무의 우점치는 65.6%였다. 산유리의 잣나무 조림지 PP Ⅱ에서 전나무의 우점치는 36.1%로 조사지역의 임분 중 전나무의 우점치가 가장 낮았다.
NA: Abies holophylla natural stand, PL: Larix kaempferi plantation, FB: Forest belt, and PP: Pinus koraiensis plantation.
오대산 지역의 전나무 천연림을 대상으로 한 선행연구에서 전나무의 중요도는 30-58%로 나타났으며, 전나무와 함께 출현하는 교목성 수종으로는 피나무, 고로쇠나무, 당단풍나무(Acer pseudosieboldianum (Pax) Kom.) 등의 중요도가 높았다(Lee et al., 2008; Han et al., 2015; Kim et al., 2015). 본 연구 지역의 수종 구성은 자연적인 분포라기보다는 조림지의 영향으로 조림 수종인 잣나무, 일본잎갈나무, 밤나무의 우점도가 높았고, 조림 수종 외에는 참나무류가 상층에 분포하였다. 강원도 오대산 지역의 천연림과 연구지인 가평은 기상, 지형 등 환경의 차이가 있으며, 인간의 간섭 정도도 다르다. 오대산 지역 전나무림은 해발고도 600-1,500 m에 위치하며, 연평균기온 6.6℃로 가평에 위치한 연구 지역보다 온도가 낮다(Han et al., 2015). 또한 오대산은 1975년 국립공원으로 지정되어 천연림이 보전되어 왔다. 반면, 연구 지역은 해발고도 200 m 이하의 낮은 구릉지로 인가와 농지가 맞닿아 있어 조림 이후에도 지역 주민의 활동이 지속적으로 이루어졌을 것이다. 이러한 환경과 인간 간섭이 수종 구성과 임분 구조의 차이로 나타난 것으로 판단된다.
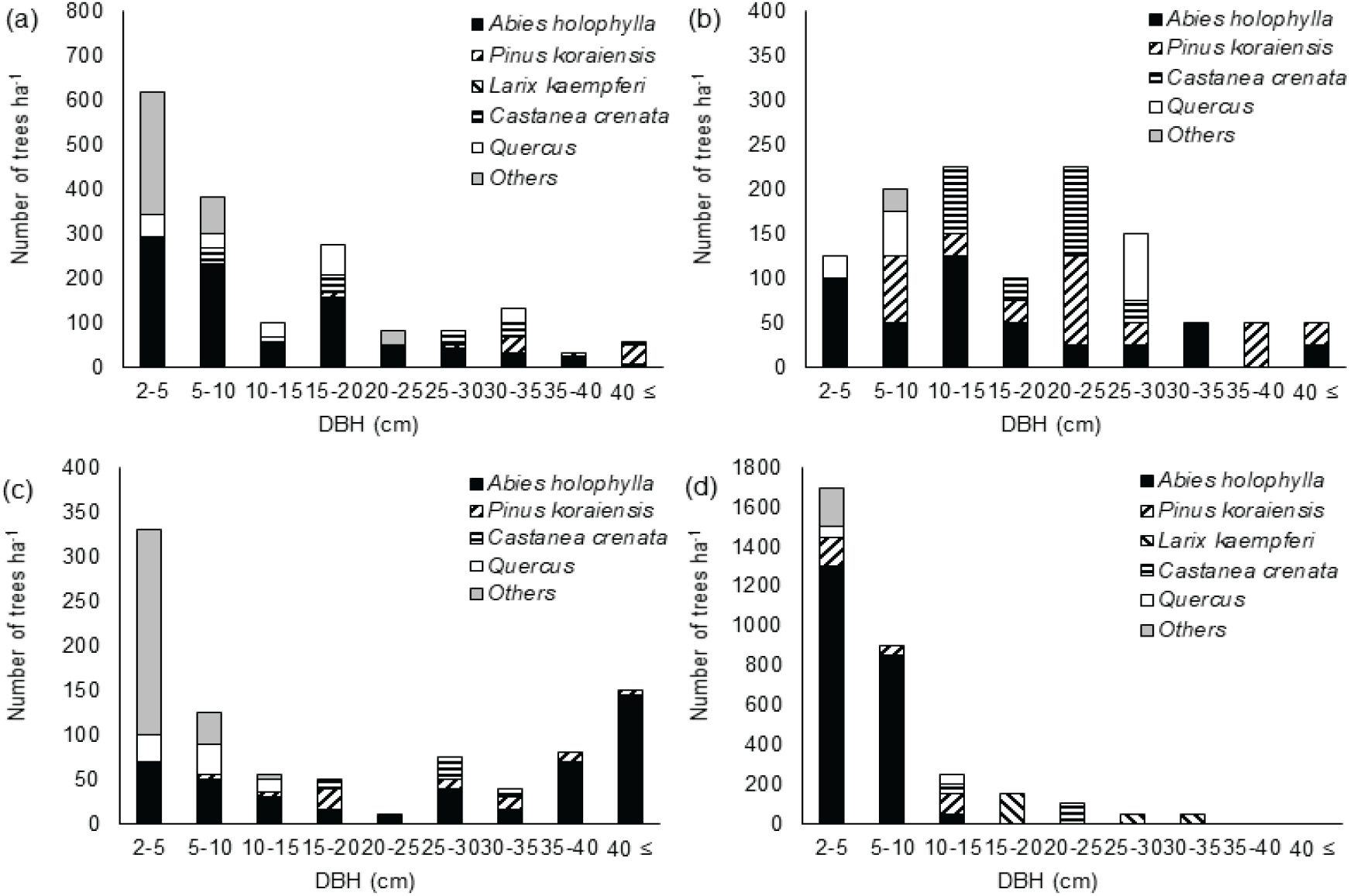
이화리 전나무 임분은 전체적으로 전나무가 40 cm 이상 직경급까지 분포하고, 20-30 cm 직경급에는 밤나무 등의 활엽수도 분포하고 있었다[Figure 3(a) and (b)]. 전나무는 전나무 임분 NA Ⅰ에서 2-10 cm 직경급의 밀도가 높아 역 J형에 가까운 직경분포를 보였으며, 전나무 임분 NA Ⅱ에서는 대부분의 직경급에서 ha당 100본 이하의 상대적으로 낮은 밀도를 보였다.
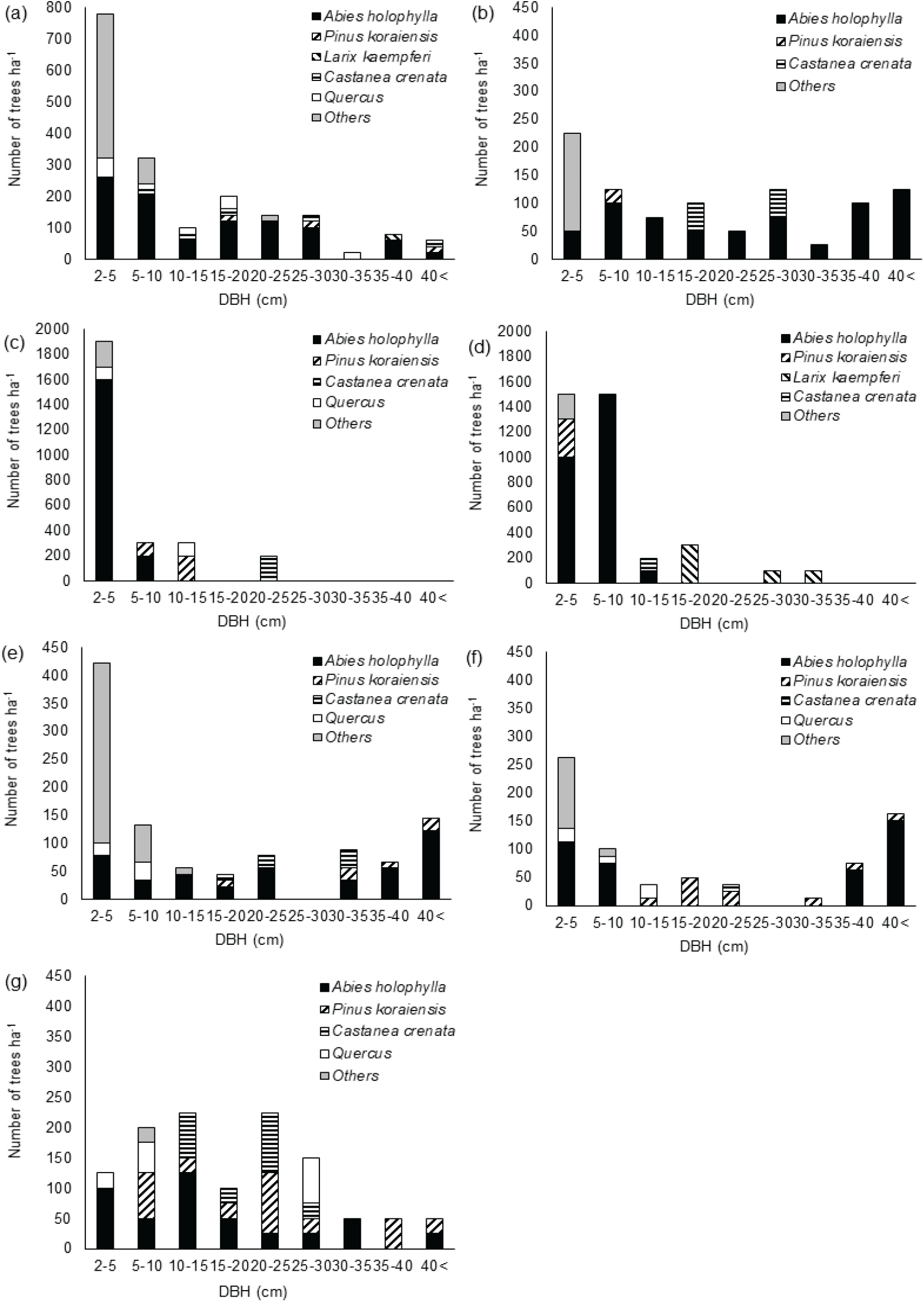
이화리의 전나무 임분 NA III와 일본잎갈나무 조림지(PL)에서 상층부는 각각 밤나무와 일본잎갈나무가 ha당 200본 내외의 밀도로 분포하고, 10 cm 이하 직경급에서는 전나무의 밀도가 ha당 1,000본 이상으로 전나무가 하층을 우점하고 있었다[Figure 3(c) and (d)]. 일본잎갈나무 조림지에서 일본잎갈나무는 15 cm 이상의 직경급에만 분포하고 있었으며, NA III에서도 밤나무는 20 cm 이상 직경급에만 분포했다. 이화리는 1960년 밤나무와 일본잎갈나무 식재 기록이 있다. NA II 임분과 NA III 임분은 능선으로 나뉘지만 인접하게 위치하고 밤나무의 직경급이 비슷한 것으로 볼 때, 이들 임분의 밤나무는 1960년 인근 지역에 밤나무 조림 시 조사지 일부에 밤나무를 식재했거나, 인근 지역에서 밤나무 종자가 비슷한 시기에 들어온 것으로 보인다.
산유리의 수림대 FB에서 전나무는 40 cm 이상과 25-30 cm의 직경급을 제외한 직경급에서 40본/ha 내외의 비슷한 밀도를 보였고 하층에서 활엽수종이 높은 밀도로 자라고 있었다[Figure 3(e)]. 잣나무 조림지 PP Ⅰ에서 전나무는 낮은 직경급과 높은 직경급에 나뉘어 분포하였다[Figure 3(f)]. 10-35 cm의 중간 직경급에는 전나무가 분포하지 않고 잣나무가 분포하고 있었으며 40 cm 이상 직경급까지 잣나무가 자라고 있었다. FB 임분과 PP Ⅰ 임분 모두 10 cm 미만 직경급에서 잣나무는 분포하지 않았고, 전나무와 참나무류, 관목류가 분포하고 있었다. 산유리의 잣나무 조림지는 2004년 숲가꾸기 작업을 실시하였다. 덩굴제거 작업을 한 것으로 기록되어 있으나 당시 하층의 솎아베기가 이루어져 중간 직경급 임목의 밀도가 낮은 것으로 보인다. 10-35 cm 직경급에 해당하는 전나무의 부재도 당시 솎아베기의 영향으로 보이며, 직경 10 cm 미만의 개체는 2004년 이후 갱신된 개체로 판단된다. FB 임분도 중간 직경급의 밀도가 낮아 숲가꾸기 작업이 있었던 것으로 보이나 PP Ⅰ에서 실시한 작업보다는 낮은 강도의 작업이었을 것으로 추정된다. 임분 내에 10 cm 미만의 직경급 임목의 벌채 흔적이 남아 있었다.
남사면에 위치한 산유리의 잣나무 조림지 PP Ⅱ는 전나무와 잣나무가 상층에서 비교적 고르게 분포하며 경쟁하는 양상을 보이고 있었다[Figure 3(g)]. 밤나무는 10-30 cm 직경급에서는 다수 분포하였으나 10 cm 미만 직경급에서는 나타나지 않았다. 이들 밤나무는 잣나무 조림목의 크기가 작을 때 인근에서 유입된 것으로 보이며, 이후 지속적인 갱신은 이루어지지 않았다. 5-10 cm 직경급에서 잣나무와 전나무가 비슷한 밀도로 분포한 반면 5 cm 미만 직경급에서는 잣나무는 나타나지 않았다. 이 임분은 임목의 직경분포와 임분 상태를 볼 때 PP I보다 낮은 강도의 숲가꾸기 작업을 실시한 것으로 보였다. 참나무류는 낮은 직경급과 20-25 cm 직경급에 불규칙하게 분포하고 있었다.
PP I은 잣나무 조림지로 상층에 종자를 공급하는 잣나무가 분포하고 있음에도 PP II에 비해 10 cm 이하 직경급의 잣나무 밀도가 현저하게 낮았다. PP I 임분은 하층에서 전나무가 참나무류와 관목류와 경쟁하는 반면, PP II는 5 cm 이상 직경급에서는 잣나무도 상당수 분포하고 있었다. 잣나무 조림지 PP I에서 솎아베기 작업으로 하층의 잣나무를 제거한 이후 그 자리를 참나무류와 관목류가 대신한 것으로 보인다.
조사 지역의 하층 전나무 밀도는 ha당 50-5,820본으로 다양하게 나타났다(Table 4). 교목층의 흉고단면적, 전나무의 흉고단면적 또는 직경 20 cm 이상의 전나무 성목의 흉고단면적은 하층의 전나무 밀도와 뚜렷한 상관관계를 보이지 않아, 전나무 성목의 밀도 혹은 상층 피도와 하층의 전나무 밀도는 연관이 낮은 것으로 나타났다. 전나무는 광도가 낮은 조건에서도 치수의 발생이 높은 것으로 알려져 있으며, 강원도 평창군의 전나무림에서 상층울폐도와 치수의 발생은 무관한 것으로 나타난 결과와도 일치한다(Chung, 1996).
DBH < 2 cm also includes trees with height < 1.3 m. NA: Abies holophylla natural stand, PL: Larix kaempferi plantation, FB: Forest belt, and PP: Pinus koraiensis plantation.
빛은 내음성 수종의 치수 발생에는 상대적으로 영향이 크지 않으나 치수의 생장과 생존에는 중요한 요소로 작용한다(Sanchez-Gomez, 2006). 이화리 일본잎갈나무 조림지와 NA Ⅲ 임분은 하층의 전나무 밀도가 다른 임분보다 높았다. 이들 임분은 상층부에 수관 틈이 발생하였거나, 상층부의 밀도가 낮기 때문에 전나무 치수가 자라고 생존하는 데에 유리하였을 것으로 판단된다. 다만 현재의 전나무 치수 밀도와 2-5 cm 직경급 임목 밀도를 고려했을 때 향후 전나무 간의 경쟁이 치열해질 가능성이 있다.
가평 지역 숲 하층의 전나무 밀도로 볼 때 이 지역은 전나무의 천연갱신이 이루어질 수 있는 기본적인 환경이 조성되어 있다고 볼 수 있다. 또한 전나무 소경목의 밀도는 이 지역이 전나무 치수가 생존하고 성목으로 자라나는 데에도 적합하다는 것을 보여준다. 한편 조사 지역의 일본잎갈나무 조림지에서 일본잎갈나무 치수가 발견되지 않았고, 잣나무 역시 전나무에 비해 적은 수의 치수가 발생하고 있었다. 일본잎갈나무는 내음성이 약한 양수로, 상층이 울폐된 조림지에서는 치수의 생존과 생장이 어렵기 때문에 일본잎갈나무 조림지 내에서는 일본잎갈나무의 갱신이 어렵고 시간이 지나며 자생수종이 일본잎갈나무를 대체하는 경향을 보인다(Kim et al., 2013). 잣나무 조림지 주변의 천연 활엽수림에서는 잣나무 치수의 갱신이 용이하나 잣나무림 내에서는 상층의 울폐 정도와 임상 환경에 따라 잣나무 갱신이 다르게 나타나며, 낙엽층이 두껍고 임내 광이 부족하면 잣나무 갱신이 어렵다(Hayashida, 1989; Ji et al., 2011). 조사 지역의 상층 밀도는 일본잎갈나무와 잣나무 치수가 갱신하기에는 울폐하여 일본잎갈나무와 잣나무 갱신은 불리한 반면, 내음성이 강한 전나무 치수는 갱신이 가능한 것으로 판단된다.
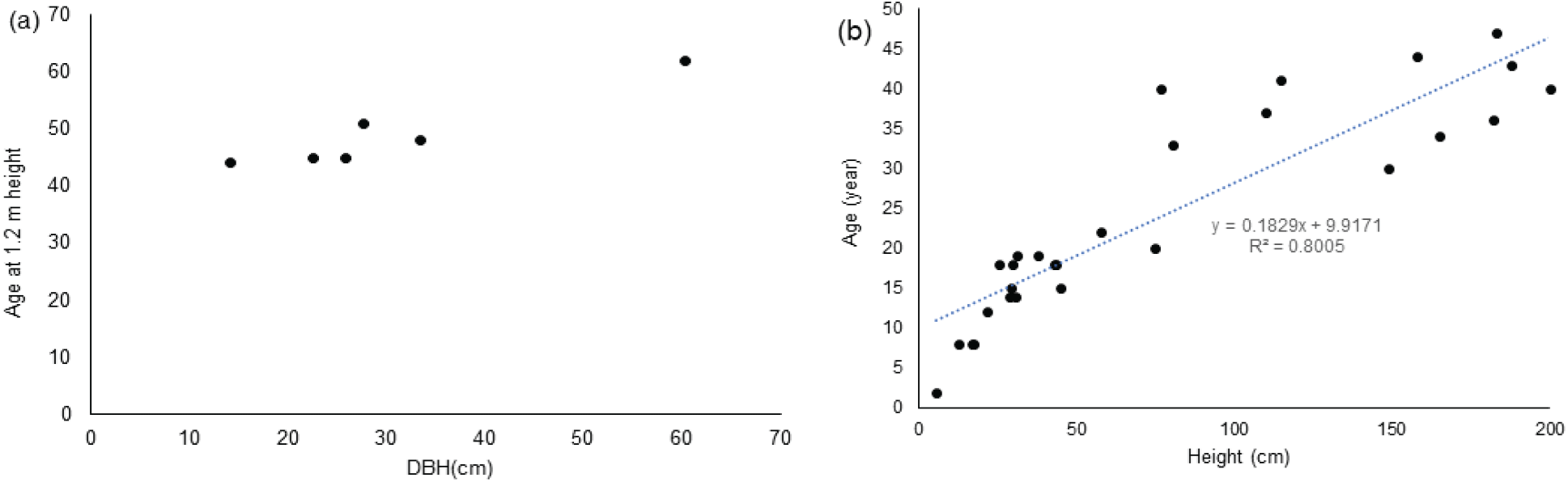
조사지에서 수고 5.5-200 cm 범위의 전나무 28본의 수령은 최소 2년에서 최대 40년으로 추정되었다[Figure 4(b)]. 하층 전나무의 수고와 수령과의 관계는 천연 임분의 하층에서 전나무가 수고 1 m 이상 자라는데 25년 이상이 걸릴 수 있다는 것을 보여준다(Choi and Lee 2014). NA I 임분에서 흉고직경 14.1-33.5 cm의 범위의 전나무는 1.2 m 높이에서 수령이 44-51년이었다[Figure 4(a)]. 조사지 내 흉고직경 60.3 cm의 전나무는 1.2 m 높이에서 수령이 62년이었다. 이를 이용하여 조사지 임분의 상층을 차지하는 직경 30 cm 이상의 전나무의 수령은 70년 이상으로 추정할 수 있다. 이는 조사지의 전나무 임분 NA I, II, III와 FB가 조림지 조성 시 능선 부근에 남겨둔 수림대에 속하며 이 지역의 천연임분이라는 것을 뜻한다. 오대산국립공원 월정사 전나무숲에서 흉고직경 11-82 cm 범위의 전나무 수령이 41-135년생이었으며 특히 흉고직경이 30 cm인 전나무는 모두 수령이 50년 이상이었다(Lee et al., 2008). 이화리의 일본잎갈나무 임분이 1960년에 조성되었고, 산유리의 잣나무 임분이 1970년대 조성되었다면 조림지 내에 분포하는 30 cm 이상의 대형 전나무는 조림지 조성 시 임분 내에 남겨두었던 잔존목으로 판단된다.
조사 임분은 4개의 유형(AN, AP, AM, P)으로 분류되었다(Figure 5). 비모수 군집분석에서 전나무의 직경분포만을 이용하였으나, 군집분석 결과는 임분 유형과 산림작업의 영향, 전나무의 분포 특성을 보여주었다.
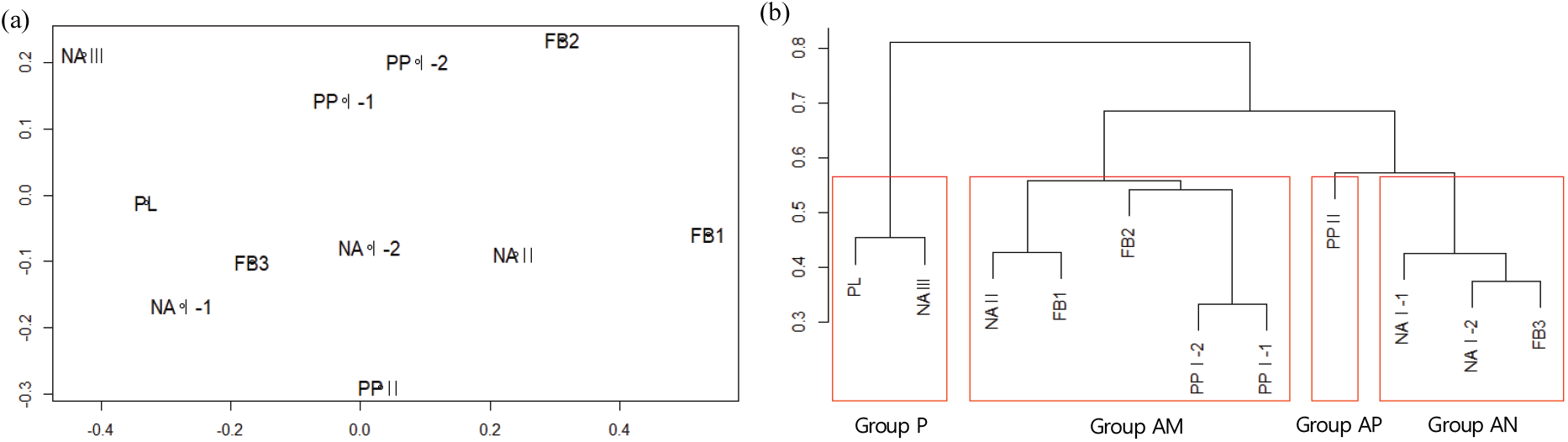
AN 유형은 이화리의 전나무 임분 NA Ⅰ과 산유리 수림대 임분의 일부가 포함되었다. 이 유형은 전나무가 낮은 직경급부터 직경 40 cm 이상의 대경목까지 연속적으로 분포하며 낮은 직경급의 밀도가 높은 역 J형의 직경분포를 보인다[Figure 6(a)]. 이는 전나무 천연림의 직경분포와 비슷한 형태이다(Chung, 1996; Lee et al., 2012). 또한 대부분의 직경급에서 전나무의 밀도가 가장 높다. 이 유형의 임분은 전나무가 우점하고 있으며 앞으로도 전나무가 우점할 것으로 예상된다.
AP 유형은 전나무 모수가 임분에 남아있는 조림지로 산유리의 잣나무 조림지 PP Ⅱ를 포함한다[Figure 6(b)]. 직경 50 cm 이상의 대형 전나무가 분포하는 것으로 보아 잣나무 조림 시 기존의 전나무를 일부 남겨둔 것으로 판단된다. 이 유형은 전나무와 잣나무가 대부분의 직경급에서 경쟁하는 양상을 보이며, 중간 직경급에서는 밤나무, 참나무류 등의 활엽수도 전나무와 경쟁하고 있었다. 30 cm 이상 직경급에서는 전나무와 잣나무가 비슷한 밀도로 분포하였고, 20-30 cm 직경급에서는 밤나무와 참나무류 등 낙엽활엽수의 밀도가 높았다. 직경 20 cm 미만의 소경목 단계에서 전나무의 밀도가 가장 높긴 하지만 잣나무나 참나무류 등 다른 수종의 밀도와 크게 차이가 나지 않았다. 전나무는 활엽수와 혼효된 상태에서는 이들과 경쟁에서 이겨 우세목으로 생장하는데 오랜 시간이 걸리는 것으로 알려져 있다(Lee et al., 1996). PP Ⅱ 임분은 비교적 광조건이 좋은 남동사면에 위치하고 있기 때문에 지속적으로 유지가 가능한 전나무림으로 변화하기 위해서는 교목성 낙엽활엽수에 대한 관리가 필요한 것으로 보인다.
AM 유형은 전나무 모수가 임분 내 분포하고 있으나 산림작업의 영향으로 중간 직경급의 전나무 임목 밀도가 낮아진 유형이다[Figure 6(c)]. 산유리의 잣나무 조림지 PP Ⅰ이 주를 이루고 있으며 산유리의 FB 임분과 이화리의 전나무 임분 NA Ⅱ가 포함되었다. 산유리의 임분은 직경 50 cm 이상 크기의 전나무도 분포하였다. 대부분의 직경급에서 전나무가 ha 당 100본 이하의 밀도를 보이고, 15-25 cm 직경급에서 급격히 낮은 밀도를 보이다 25 cm 직경급부터 다시 증가하여 변동하는 모습의 직경분포를 보였다. 산유리 지역은 약 45년 전 잣나무를 식재한 기록이 있고, 임분의 잣나무 평균 흉고직경은 24.6-34.9 cm였다. 현실림 임분수확표에 따르면 임령 45년의 잣나무 흉고직경은 약 30 cm 내외로(National Institute of Forest Science, 2016), 산유리의 잣나무림은 기록에 남아있는 식재 사업에 의해 조성된 것으로 보인다. 잣나무의 식재 기록과 상층 전나무의 직경은 임분 상층의 전나무가 잣나무 조림 시 남겨둔 잔존목이라는 것을 알려주며, 이들이 전나무 갱신에 모수의 역할을 한 것으로 판단된다.
AM 유형에서 10-25 cm 직경급의 밀도가 낮은 것으로 보아 산유리에서 2004년 실시한 숲가꾸기는 하층간벌이었을 것으로 추정된다. 20 cm 미만 직경급에서는 전나무가 역 J형의 직경분포를 가지고, 수종 구성도 참나무류와 관목류가 다수를 차지하여 NA I 임분과 비슷한 경향을 보인다. 따라서 AM 유형은 향후 전나무림으로 발전할 가능성이 높다.
P 유형은 전나무 모수가 임분 내에 존재하지 않으나 인근에 존재하는 조림지 유형이다[Figure 6(d)]. 2-10 cm 직경급에서 전나무의 밀도가 약 2000본/ha로 매우 높은 반면, 직경 15 cm 이상의 상층을 차지하고 있는 일본잎갈나무, 밤나무 등의 조림 수종은 그 밀도가 낮고 치수 또한 나타나지 않았다. 따라서 조림 수종의 천연갱신은 이루어지지 않고 가까운 거리에 위치한 전나무 임분으로부터 종자가 유입되어 전나무로의 천연갱신이 진행되고 있는 것으로 보인다. 실제로 종자를 생산할 수 있는 전나무가 있는 임분과 P 유형 임분의 거리는 20 m 이내로 주변으로부터 종자 유입이 가능한 거리였다. 전나무속은 종자의 90% 이상이 반경 50 m 이내에 떨어지는 종자 산포 범위를 가지는 것으로 알려져 있다(Franklin and Smith, 1974). 또한 전나무 치수의 유전적 동질성이 나타나는 범위는 20 m 이내인 것으로 나타났다(Hong et al., 2001). 이 유형의 경우 임분 내 전나무 소경목의 높은 밀도로 볼 때 조림 수종은 전나무로 대체되고 AN 유형의 임분과 같은 연속적인 전나무 분포를 보이는 전나무림으로 발달할 것으로 예상된다.
전나무의 분포 범위는 경기도와 강원도를 포함하고 있으며, 전나무는 우리나라의 다른 전나무속 수종보다 낮은 고도에 분포한다(Korea National Arboretum, 2015). 전나무 식재 기록이 없음에도 이화리의 전나무 임분과 같이 활발한 치수 발생과 안정적인 직경 분포를 보이는 전나무 임분이 가평 지역 내에 분포하고 있다. 가평에 인접한 광릉 지역에도 전나무 천연림이 존재한다(Kim and Lee, 2019).
이화리와 산유리의 조사지에서 하층에 발생하는 교목 치수의 대부분이 전나무였고, 전나무는 임분 내에서 유령목과 성숙목으로도 자라고 있었다. 이화리의 조림지는 전나무 종자를 공급할 수 있는 전나무 천연 임분과의 거리가 전나무속의 종자 산포 범위 이내이고, 산유리의 조림지는 종자를 공급할 수 있는 전나무 모수가 조림지 내에 남아있었기 때문에 이러한 결과가 나온 것으로 보인다. 반면, 잣나무와 일본잎갈나무는 상층에 모수가 우점하나, 하층에 치수의 발달이 미약하였다. 전나무의 갱신 현황과 직경분포를 볼 때 전나무는 치수가 지속적으로 갱신되고 있고, 낮은 직경급부터 높은 직경급에 걸쳐 고루 분포하며 다른 수종보다 높은 밀도를 보이고 있었다. 큰 변화가 없다면 조사 지역의 임분은 향후 상층도 전나무가 우점하는 임분으로 발달할 것으로 예상된다.
단일 수종 동령림으로 조성된 조림지에서 자생수종의 숲으로 복원되는 데에는 종자 공급원과의 거리, 지형적 특성, 기후, 임목밀도와 임분 상층 구조 등이 영향을 미친다. 천연갱신을 이용한 수동적 복원(passive restoration)은 종자의 양과 다양성을 증가시키는 천연임분과의 근접성을 우선적으로 고려해야 하는 것으로 알려져 있다(Kremer and Bauhus, 2020).
가평 지역의 일본잎갈나무와 잣나무 조림지에서 종자의 공급이 가능한 전나무 모수가 남아있거나 인접한 곳에 전나무림이 위치한다면 자생수종인 전나무의 천연갱신이 가능하고 이들 조림지가 점차 전나무림으로 천이가 진행되어 수동적 복원이 이루어질 가능성이 있다는 것을 보여준다. 전나무의 입지, 전나무 치수 발생과 생장, 내음성에 관한 추가 연구가 계속된다면 경기 지역 내 생태적 천이를 이용하여 자생수종의 복원을 유도할 수 있는 관리 방법을 개발, 적용할 수 있을 것으로 기대한다.
임분의 상층을 유지한 상태에서 천연갱신을 이용한 임분 복원은 복층림 형성이 단계적으로 수반되기에 하층에서 생장할 수 있는 자생수종의 내음성과 생장에 관한 정보가 필요하다. 향후 자생수종의 내음성과 생장에 관한 연구가 이루어지고, 이를 바탕으로 한 갱신과 관리 기술이 개발된다면, 노령림 복원, 복층림이나 택벌림 경영도 가능할 수 있을 것이다.
결 론
경기도 가평 지역에서 전나무가 분포하는 임분을 대상으로 임분구조와 하층의 전나무 천연갱신, 조림지에서의 천이와 자생수종 임분으로의 복원 가능성을 조사하였다. 전나무는 조림지를 포함한 조사 지역의 모든 임분에서 높은 우점치를 보였다. 또한 타 수종의 조림지에서도 조림지 내에 종자 공급이 가능한 전나무 모수가 남아있거나 전나무 임분이 인접해있으면 전나무의 천연갱신이 조림 수종보다 활발하게 이루어지고 있었다. 직경 분포와 하층 밀도는 조림지와 산림 작업에 따라 차이가 있었으나, 조사지의 모든 임분은 향후 전나무림으로 발달될 가능성을 보였다. 가평 지역에서 전나무의 천연갱신 현황은 가평 지역이 전나무 생육에 적합한 곳임을 나타내며, 전나무 모수가 인접한 조림지는 천연갱신을 이용한 전나무림의 복원이 가능할 것으로 예상된다. 추가적인 연구를 통해 생태적 천이를 이용하여 자생수종이 우점하는 임분의 복원을 유도할 수 있는 관리 방법을 개발, 적용할 수 있을 것으로 기대한다.