



서 론
군집의 종조성 및 생물다양성 양상은 생육하는 종들의 의 공동 분포(co-distribution)에 의해 형성되며, 지위 및 환경 여과 또는 확률성(stochastic power)에 따르는 것으로 알려져 있다. 잠재 식생으로서 매토종자의 조성과 다양성은 군집 조합 특성을 이해하는 좋은 요소이다(Forey et al., 2011). 매토종자는 식물 군집의 구성을 유지하고 교란 후 종 공급 및 보충 원으로서 역할하며, 개체군에 새로운 유전자형을 도입시킬 수 있다(Cavers, 1995; Leck et al., 2008). 매토종자는 교란 후 초기 재생 과정 및 과거 식생을 유추할 수 있는 중요한 정보원으로서(Hopfensperger, 2007), 토지이용 역사를 반영하고 현재 식물 개체군의 다양성과 동태에 기여하고 있다(Lang and Halpern, 2007). 많은 종들은 수명이 긴 종자를 가지고 있으며(Thompson et al., 1997), 식물군집에서 매토종자는 과거, 그리고 현재 이입하는 시간적으로 혼합된 군집으로서 역사성을 내포하고 있다(Thompson, 2000).
매토종자는 주로 초지 및 교란 후 천이 초기 서식지에서 많은 연구들은 이루어졌다(Thompson and Grime, 1979; Thompson et al., 1997; Thompson, 2000; Yu et al., 2007; Basto et al., 2018). 농업 이용 후 식생 재생 과정(초지에서 성숙림)에서 매토종자 군집의 발달 과정 역시 연구된 바 있고(Cho et al., 2018), 온대 성숙림 지역의 매토종자 특성 역시 연구된 바 있다(Leckie et al., 2000). 매토종자의 생태적 기능을 이해하기 위해서는 가능한 인간에 의한 변형이 거의 없어 가장자리 효과가 적고 외래종의 구성비율이 낮은 숲 생태계에 많은 관심이 필요하다(Pickett and McDonnell, 1989; Matlack and Good, 1990). 특히, 온대림에서 시간과 공간 진행이 복합적으로 포함되는 토양 깊이 증가에 따른 매토종자의 변화 양상에 관한 연구는 찾아보기 어렵다.
생태 군집으로서 매토종자 조합 및 그것의 변화는 결정론, 그리고 확률성 과정들의 중요성 변화를 따르며, 특히 상층(지표 식생에서 표토, 그리고 표토에서 심토)으로부터 지속적 종자 공급과 소실 같은 복잡한 생태 과정의 결과이다(Royo and Ristau, 2013). 지상에서 토양으로 유입된 종자의 크기 및 수명은 변이(종 간 및 종 내)를 동반하기 때문에 매토종자는 지상 식생과는 이질적이다(Simpson et al., 1989; Cho et al., 2018). 매토종자의 풍부도와 밀도는 토양의 깊이에 따라 다르게 나타나고(McGraw, 1987), 천이 단계의 진행에 따라 감소하는 것으로 알려져 있다(Thompson, 1978; Pickett and McDonnell, 1989; Cho et al., 2018). 예를 들면, 종자 크기는 생존력, 치수의 정착 성공 및 생장, 경쟁 능력 그리고 지속성에 큰 영향을 미친다(Westoby et al., 1992; Jakobsson and Eriksson, 2000). 토양 깊이 증가에 따른 매토종자 군집의 변화에서, 환경 변화(토양 물리 및 화학 속성), 그리고 종자 수명 및 수직 이동에 유리한 작거나 가벼운 종자 크기 같은 종 형질(species traits)의 영향일 수 있다(Chambers and MacMahon, 1994). 이러한 점에서, 매토종자 내 다양성을 포함한 군집 구성에 대한 결정론적 그리고 우연(예, 종자의 소멸과 이입으로서 수직 이동 사건)의 두가지 중요 생태 과정의 영향이 존재할 것이다. 먼저, 종자 수명 같은 결정론적 과정이 지배적이라면 매토종자 군집은 종 풍부도(richness)와 수도(abundance) 모두 감소하게 될 것이며(α-diversity 감소), 그에 따라, 매토종자 구성 및 관련 다양성은 수명이 길거나 미세한 종자 크기의 종 조합으로 수렴하여 점차 균질화 될 것이다(ß-diversity의 감소)(Dupuy and Chazdon, 1998; Van Calster et al., 2008; Plue et al., 2010).
결정론과 대비되는 확률론적 과정(stochastic processes)으로서, 매토종자의 수직 변화에서 무작위 및 우연적인 종자 소실/이입 사건이 중요하게 관여할 수 있다. 확률적 사건이나 지역 생물 및 비생물적 요인(예, 빛 가용성, 토양수분), 토양 내 종자의 수직 이동에 따른 이입 및 쇠락(고갈 또는 모두)이 지배적이라면, 종 풍부도(richness)와 개체수(abundance) 변화는 무시할 수 있거나 예측할 수 없으며 매토종자 군집은 시간이 지남에 따라 더 큰 독립성과 임의성을 나타낼 것이다(Marquis, 1975; Granstrom, 1987; Kjellsson, 1992; Jankowska-Błaszczuk et al., 1998; Leckie et al., 2000; Bossuyt and Hermy, 2001).
우리는 한국의 대표 저지대 성숙림인 광릉숲에서 토양 깊이 증가(상토 및 하토 군집)를 종 조합 해체 과정으로 설정하였다. 그리고 해체 과정에 대한 우점 과정을 이해하기 위해 구성 종의 형질(종자 크기 및 무게), 그리고 토양 층위 간 종 조성을 포함한 α-, ß-, 그리고 ζ-diversity(이후 제타다양성)의 변화 양상을 이용하였다.
재료 및 방법
본 연구는 한국의 유네스코 생물권보전 지역인 광릉숲(2,400 ha)에서 수행되었다. 광릉숲은 수도 서울과 인접하며 수리봉, 죽엽산, 물푸레봉, 그리고 천겸산 등으로 구성되어 있다(N 37° 42’~37° 45’, E 126° 00’~126° 02’). 광릉숲은 1468년 조선 제7대 왕 세조의 능 부속림으로 지정된 이후, 엄격한 출입 및 이용 관리가 이루어진 역사적인 보호구역이며, 약 550여년 동안 화재 같은 대규모 교란의 기록이 존재하지 않는다(Cho et al., 2007). 이러한 문화 및 경관생태적 배경을 갖는 광릉숲은 한국을 비롯한 동북아시아에서 찾아보기 어려운 저지대 성숙림이 잔존하고 있는 중요 지역이다. 광릉숲 활엽수림은 주로 졸참나무가 우점하며, 사면 배수가 좋은 일부 장소는 서어나무, 그리고 건조하거나 기반암이 노출된 장소는 굴참나무, 그리고 하변림은 갈참나무와 귀룽나무가 우점한다(Cho et al., 2007). 침엽수림은 소나무림, 그리고 조림된 잣나무, 전나무, 그리고 리기다소나무가 우점하는 군반들이 존재한다. 광릉숲은 수도권에 위치하여 지속적으로 도시화 및 산업화 영향을 직접, 간접적으로 받고 있다. 최근 20년 동안 광릉숲 주변 지역의 논과 밭은 대규모 아파트 단지와 산업 시설로 거의 모두 전환되었다(Cho et al., 2021). 광릉숲의 연평균기온은 11.8°C, 평균 연강수량은 1,387.3 mm로서 대륙성 기후이며, 온대북부와 접하는 온대중부 식생기후에 포함된다(Cho et al., 2021). 4개 지역(한랭, 상량, 온난 및 해양성 온난 온대)으로 구분되는 한국의 식물상 지역 중, 광릉숲은 온난온대 식물상 지역에 포함된다(Jung and Cho, 2020).
2011년 전체 광릉숲에서 국립수목원 관리 공간(약 1,100 ha) 중 자연 식생에 대하여, ArcGIS 10.1을 활용하여 격자망(300 m×300 m)을 생성하였다. 생성된 격자망에서 조사 활동이 거의 불가능한 급경사 지형, 그리고 천이 초기 단계 같은 이질적 식생을 제외하여 106개 격자를 선택하였고, 각 격자 중심에 각 모서리가 동서남북을 향하는 0.12 ha(35 m×35 m) 방형구를 설치하였다. 전체 방형구 면적의 합은 12.7 ha이다.
각 방형구는 교목 집단의 변화 관찰에 사용되었다. 2011년 9월~10월, 각 방형구에서 흉고직경 5 cm 이상의 교목 개체에 대하여 종을 기록하고 흉고(높이 1.5 cm) 직경을 측정하였다. 수관 환경으로서 수관 열림도 canopy openness(%)는 어안렌즈를 사용하여 조사구의 우측 모서리, 좌측 모서리 및 중앙 지점의 세 개 지점에서 촬영 이미지를 얻었다. 촬영된 수관 이미지는 Gap Light Analyzer 2.0을 사용하여 분석하였다(Frazer et al., 1999).
106개 방형구에서 얻어진 교목의 줄기 밀도는 서어나무(164.8±131.3 stems ha−1),졸참나무(79.2±73.1 stems ha−1)및 소나무(41.4±62.5 stems ha−1) 순으로, 그리고 흉고단면적은 소나무(6.9±10.2 m2 ha−1), 졸참나무(6.6±7.8 m2 ha−1) 및 서어나무(4.6±3.7 m2 ha−1) 순으로 높았다. 수관열림도 canopy openness는 소나무림(23.3±8.9%), 졸참나무림(19.5±12.4%) 및 서어나무림(11.5±6.3%) 순으로 높았고, 전체 조사구의 평균 수관열림도는 18.1±9.2%였다(Appendix 1).
초본식생의 변화를 관찰하기 위해 각 방형구 중심에서 세 방향(0°, 120°, 240°)으로 2 m 떨어진 지점에 1 m2(미세방형구)로 세분된 4 m2 소방형구를 설치하였다. 각 미세방형구에서 식물종을 기록하며, 또한 각 종의 피도(%)를 최소단위 0.1로 기록하였다. 각 미세방형구에 출현한 식물 별 피도를 평균하여 하나의 소방형구 값으로 하였고, 세 소방형구에 대한 평균 값을 방형구의 초본식생 속성으로 하였다.
전체 106개 방형구에서 졸참나무, 서어나무 및 소나무가 우점하는 27개 방형구를 매토종자 실험 장소로 선택하였다. 각 방형구에는 하층식생을 위한 세 개의 소방형구가 설치되어 있고, 각 소방형구를 구성하는 네 개 미세방형구에서 하나를 선택하여 매토종자 실험을 위한 토양 시료를 채취하였다. 개별 미세방형구에서, 토양 시료는 직경 5 cm, 높이 5 cm의 토양 채집 캔(부피 98.2 cm3 및 면적 19.6 cm3)으로 낙엽층과 부식층을 제거한 후, 상토(upper soil 0~5 cm), 그리고 10 cm 간격을 두고 하토((lower soil 15~20 cm)의 두 층위에서 채집하였다. 토양 시료는 각 미세방형구에서 무작위로 세 개 지점을 선택한 후, 2개 토양층에서 3회 토양을 채취하였다. 채취된 토양 시료는 층 별로 섞은 후 발아 실험에 사용하였다.
본 연구는 멧돼지의 먹이 활동이 관찰되지 않은 장소에서 채집된 토양을 활용하였다. 멧돼지의 먹이 활동은 토양을 교란시키며, 매토종자에 영향을 줄 수 있다. 멧돼지는 참나무과 수종이 우점하는 환경을 선호하는데(Kim et al., 2019), 광릉숲의 사면은 졸참나무, 그리고 산록완사면 및 범람원은 갈참나무가 우점하고 있다. 또한 연구 지역 인근의 개발은 지역 멧돼지 개체군이 비교적 인간 활동이 적은 광릉숲으로 이동하는 배경이 되고 있다. 그러나 본 연구의 결과는 이러한 멧돼지의 토양 교란 영향을 제한하여 실행되었다.
매토종자에 대한 발아실험은 국립수목원의 온실에서 실시하였다. 채취한 토양 시료는 실내 조건에서 일주일 건조 후 체를 이용하여 뿌리와 근경 같은 번식체를 제거하였다. 걸러진 토양 시료는 2 cm× 36 cm×9 cm 크기의 삽목상자에 배양토를 7 cm두께로 깔고 그 위에 3 cm 깊이로 고르게 포설하였다. 기온이 높지 않은 4~7월에는 2일 1회, 기온이 높은 8월~9월에는 1일 1회 물을 주었다. 지면의 종자가 유입되는 것을 막기 위하여 1 m 높이의 선반 위에서 실험을 진행하였다. 버드나무와 같이 바람에 의한 종자 유입을 막기 위해 0.1 mm×0.1 mm mesh의 천을 덮고 관수 시에만 제거하였다. 6월, 7월 및 9월 총 3회에 걸쳐 발아된 식물의 종명과 개체수를 기록하였다.
동정된 개체는 항상 제거하였고, 묻혀 있는 종자들이 발아할 수 있도록 원예용 포크를 사용하여 토양을 꾸준히 섞어주었다. 동정이 불가능한 개체는 포트로 옮겨 동정이 가능해질 때까지 성장시켰으며, 불가능 한 개체는 속 수준으로 동정하였다(예, 사초류). 출현식물 및 발아식물의 식별은 이동혁(Lee, 2014), 김태영 및 김진석(Kim and Kim, 2011) 및 이창복(Lee, 1989), 그리고 학명은 산림청의 국가표준식물목록(Korea National Arboretum, 2017a)을 따랐다.
우리는 식생 층위(지표, 표토 및 심토)에 따른 종 조합 변화 및 관련 다양성 양상을 분석하였다. 각 방형구에 대하여, 토양 채집 캔의 면적(19.6 cm)을 cm2로 변환하여 종의 발아 밀도를 계산하였다. 또한 전체 방형구에 대한 개별 종의 출현 빈도(%)를 계산하였다. 식피율로 기록된 미세방형구의 초본 피도 및 출현 빈도를 계산하여 식생 층위 별 구성 비교에 활용하였다. 토심에 따른 매토종자의 형질 변화 분석을 위해 한국 야생식물 종자도감 Seed Atlas of Korea(Korea National Arboretum, 2017b)을 참고하여 발아된 종 별 종자 크기(mm2, length×width) 및 천립중(1000 seeds weight)(g)을 구하였다. 토양 층위에 따른 α-다양성, 서식지 선호성 habitat affinity, 기원 species origin, 그리고 생육형 growth form을 포함한 종풍부도(species richness) 및 밀도(germinant density), 그리고 종자 형질 seed trait(무게 및 크기) 비교는 모두 paired t-test를 사용하였다.
서식지 선호성은 저자들의 전문 시각을 따랐으며, ⅰ) 숲 속에 출현하는 종(Forest species; 수관이 울폐된 곳에서 잘 살아가는 광 요구성이 높지 않은 종), 숲의 가장자리 또는 수관이 열린 지역에서 출현하는 종(Edge species; 숲에서 출현하는 종 중 광요구성이 높은 종), 길가 및 빈터에서 출현하는 종(Ruderal species; 주된 서식지가 숲이 아닌 광 요구성이 높은 천이 초기 환경의 주요 종)의 세 유형으로 구분하였다. 지리적 기원은 자생 또는 외래 식물로 구분하였다. 마지막으로 생육 형태는 교목(tree), 관목(shrub), 덩굴성 목본(vines) 및 광엽초본(forbs) 및 협엽초본(graminoids), 그리고 양치식물(ferns)의 여섯 개 유형으로 구분하였다.
전통적인 베타다양성(ß-diversity)를 대신하여, Non parametric Multidimensional scaling(이후 NMDS) 서열 분석을 통한 군집 조성 내의 이질성, 그리고 개별 방형구 군집 간 공통 출현 종수로서 제타다양성(ζ-diversity, ζi)의 두 가지 도구를 활용하였다. NMDS를 통해, 토양 층위 내 방형구 군집 자료의 퍼진 정도는 매토종자 종조성의 단순화 또는 이질화를 나타낸다. 서열 분석에 적용된 변수는 개별 식물 종의 피도(하층식생) 및 밀도(매토종자) 값이다.
일반적으로 베타다양성(예, Jaccard, Sørensen 또는 Simpson의 이질성 값을 사용)은 두짝 비교(pair-wise comparison)로서 n개 조사구 내 종 구성 변환(species turnover)의 단일 값으로 표현하는데(Jost et al., 2011), 제타다양성은 다양성 변화의 구조화 방식에 대한 이해도를 향상시킨 측정 도구이다(McGeoch et al., 2019).
제타다양성은 i 개 군집(방형구 같은 조사 단위)으로 조사된 개별 종조합에 대한 종 교대(species turnover) 측정을 제공한다. 간단히, 제타다양성은 i개 군집이 공유하는 출현 종 수를 나타내며(말하자면, i개 조사구의 종 교집합), i는 제타 순차(zeta order)로 표현된다. 제타다양성의 측정 단위는 ‘공유된 종 수’이며, 최소 값은 0(둘 또는 그 이상의 조사 장소들은 공유된 종이 없음), 그리고 최대 값은 군집 자료의 총 종풍부도의 영향을 받는다. 제 1 제타 순차의 값은 자료의 평균 종풍부도와 동일하다. 제타 순차의 증가는 필연적으로 공통 출현하는 종의 감소를 가져오므로 i의 증가에 따른 제타다양성은 항상 감소한다.
제타다양성 감소의 보완 도구로서 제타다양성 잔존 비율(zeta-diversity ratio) 분석을 역시 실시하였다. 제타 비율(Zeta ratio)은 어떤 순차에서 공통 출현종이 다음 순차에서 출현할 확률을 나타내며, 초기 순차에서 증가는 조사자료 내 희귀종, 그리고 제타 순차 후기로 갈수록 일반종의 기여가 증가하는 것이 표현된다(McGeoch et al., 2019). 예를 들면, 순차 9에서의 잔존 비율은 조사된 10개 조사구 중 적어도 9개에서 잔존하는 종들의 출현 확률로 해석할 수 있다. 제타 비율의 변화 곡선은 자료의 성질에 따라 증가형 및 감소형, 점근형(asymptotic) 및 봉형(modal)의 모양을 보인다(McGeoch et al., 2019).
토양 깊이 발달에 따른 종 조합 변화 양상에서 우점하는 생태 과정(결정성 또는 확률성)의 평가는 제타다양성 변화 양상의 멱함수(power function) 및 지수 함수(exponential function)에 대한 적합도 검정을 이용하였다. 적합도의 유의성은 Akaike’s Information Criterion(AIC)을 이용하였다(Akaike, 1974). 제타다양성 양상에 대한 지수 및 멱함수 적합은 생태 과정에 관한 서로 다른 가설들이 관계되는데, 종 교대는 확률론적(exponential zeta fitting, 조사구 수와 독립), 또는 니체 분화 과정(power zeta fitting, 조사구 수와 비독립)을 따른다는 가설들이다(Scheiner et al., 2011).
단변량 및 다변량 통계 분석은 R studio 프로그램을 사용하였으며, ζ-diversity 분석은 zetadiv 패키지(Latombe et al., 2018), 그리고 서열 분석은 vegan 패키지의 함수 ‘metaMDS’를 이용하였다(Oksanen et al., 2019).
결 과
하층식생에서 59분류군(36과 50속 52종 4변종 1아종 2품종)의 식물이 출현하였고, 평균 종풍부도 및 식피율(%)은 각각 8종 및 11.5%였다(Table 1 및 Figure 1). 생육형(Growth form)에서, 광엽초본 15종, 교목 18종, 관목 9종, 협엽초본 9종, 만목 6종, 그리고 양치식물 2종의 순으로 풍부도가 높았다. 서식지 선호성(Habitat affinity)은 숲종이 42종(하층식생 전체의 71.2 %) 및 총 식피율 64%로 가장 큰 비중을 차지하였고 가장자리종 14종(33%) 및 빈터종 3종(3%) 순으로 풍부했으며, 외래종은 출현하지 않았다.
Note: Codes for habitat affinity (Hab.) are forest (F), edge (E) and ruderal (R). Codes for origin (Ori) are native (N) and exotic (E). Cover < 0.1% is report as “t” (trace).
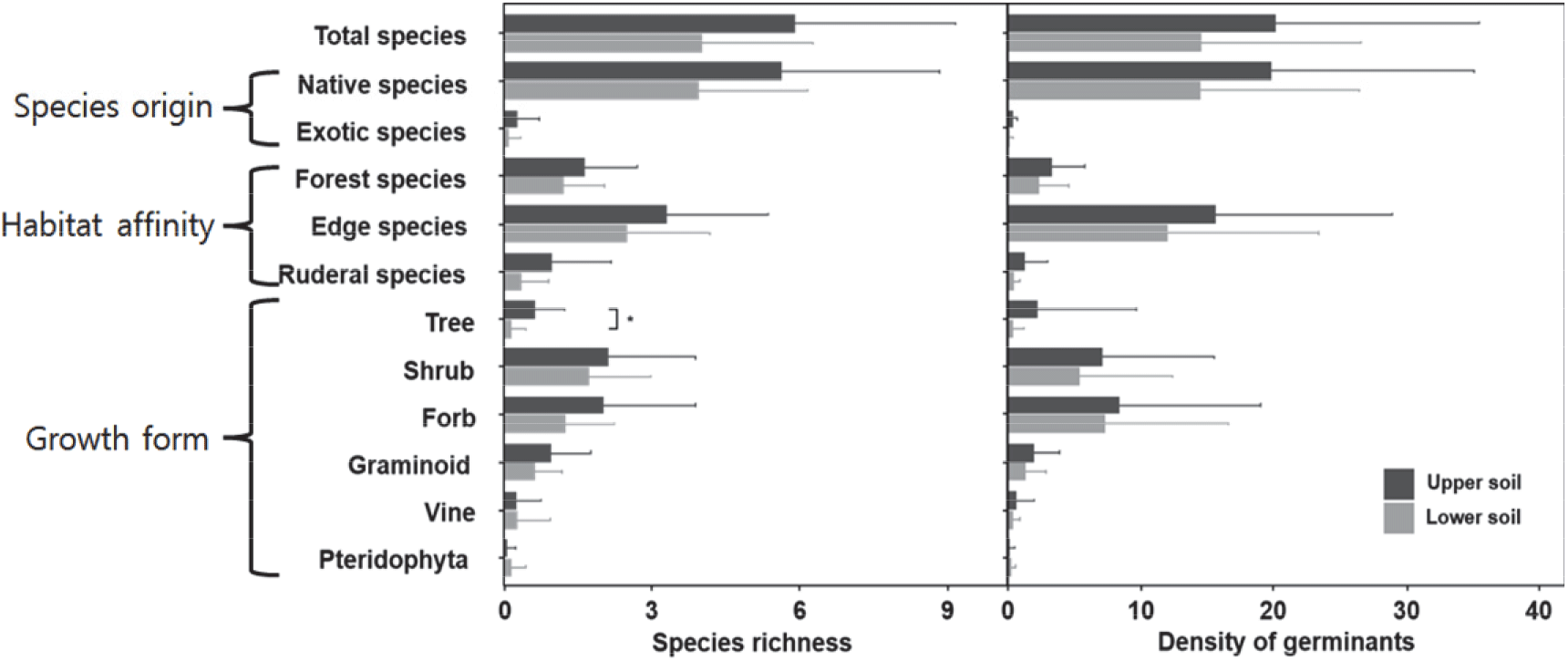
발아 실험을 통해 47 분류군(27과 40속 44종 3변종)을 포함하는 934개체가 발아하였다. 토양 층위에 관계없이, 방형구 별 평균 4.9종 및 17.3개체(단위 면적의 평균은 326.3 germinant m-2)가 발아하였다. 발아된 식물 중, 교목은 5종(10.6 %), 관목은 9종(19.1 %), 만목은 3종(6.3 %), 양치식물 1종(2.1 %), 협엽초본 4종(8.5 %), 광엽초본 25종(53.2 %)이었다(Table 1). 가장자리종은 22종(발아 개체의 77 %)으로 가장 풍부하였고, 이후, 빈터종(13종 및 발아 개체의 5 %), 숲종(12종 및 발아 개체의 18 %) 순이었다. 외래종은 모두 광엽초본 3종(개망초 Erigeron annuus, 망초 Conyza canadensis, 붉은서나물 Erechtites hieracifolia)이었고, 상토에서 3종, 그리고 하토에서 1종이 출현하였으며, 발아 개체 중 1 % 이하로 매우 낮은 구성비를 보였다.
상토(0~5 cm) 및 하토(15~20 cm)에서 종풍부도 및 개체수는 통계 유의성은 없었지만 큰 편차와 함께 감소하는 경향이었다; 교목(상토 0.11±0.31종 및 하토 0.26±0.84 종, p=0.013)을 제외하면 세 가지 생태 형질(종의 기원, 서식지 선호성 및 생육형) 모두 통계적 유의성은 없었다(Figure 1). 세 식생 층위의 공통 종은 7종이었고, 상토 및 하토 사이에는 오동나무, 병꽃나무, 작살나무, 국수나무의 공통종을 포함한 23종이었다. 상토에서만 발아한 종은 넓은잎외잎쑥, 덩굴곽향, 맑은대쑥을 포함한 15종이었고, 하토는 애기도둑놈의갈고리, 가시꽈리, 그리고 잔털제비꽃을 포함한 8종이었다(Table 1).
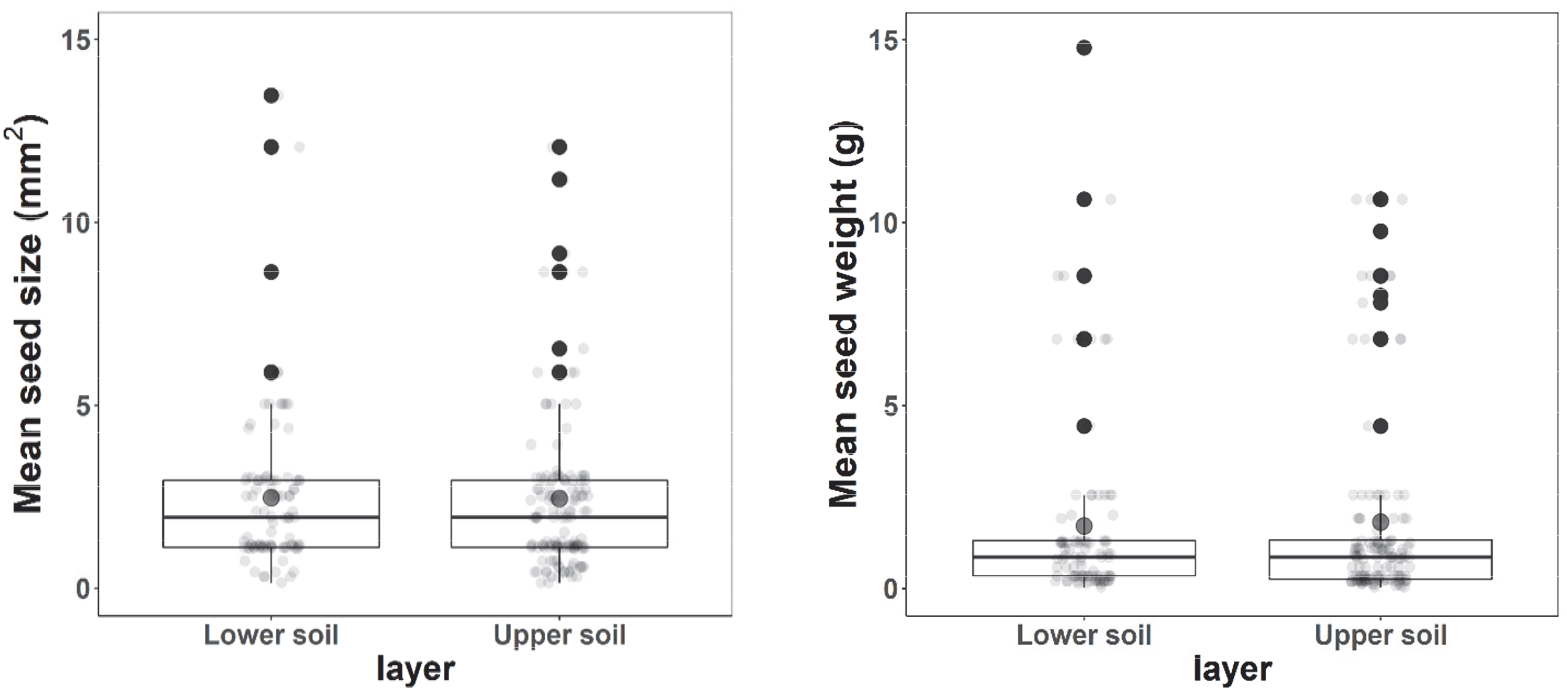
상토 및 하토에서 발아된 식물의 평균 종자 크기(상토 3.2±3.1 mm2 및 하토 3.2±3.2 mm2) 및 천립중(상토 2.1±3.0 g 및 하토 2.2±3.4 g)은 모두 유의한 차이가 없었다(Figure 2). 비교적 크고 무거운 종들에 의해 크기 및 천립중의 큰 편차가 관찰되었다; 까치박달(크기 9.1 mm2 및 천립중 8.0 g), 붉나무(크기 5.9 mm2 및 천립중 8.5 g), 산초나무(크기 8.6 mm2 및 천립중 10.6 g), 싸리(크기 5.0 mm2 및 천립중 6.8 g) 및 소나무(크기 11.2 mm2 및 천립중 9.8 g) 같은 목본 5종, 그리고 할미밀망(크기 12.1 mm2 및 천립중 4.4 g) 및 애기도둑놈의갈고리(크기 13.5 mm2 및 천립중 14.8 g) 2종의 초본식물이 포함되었다.
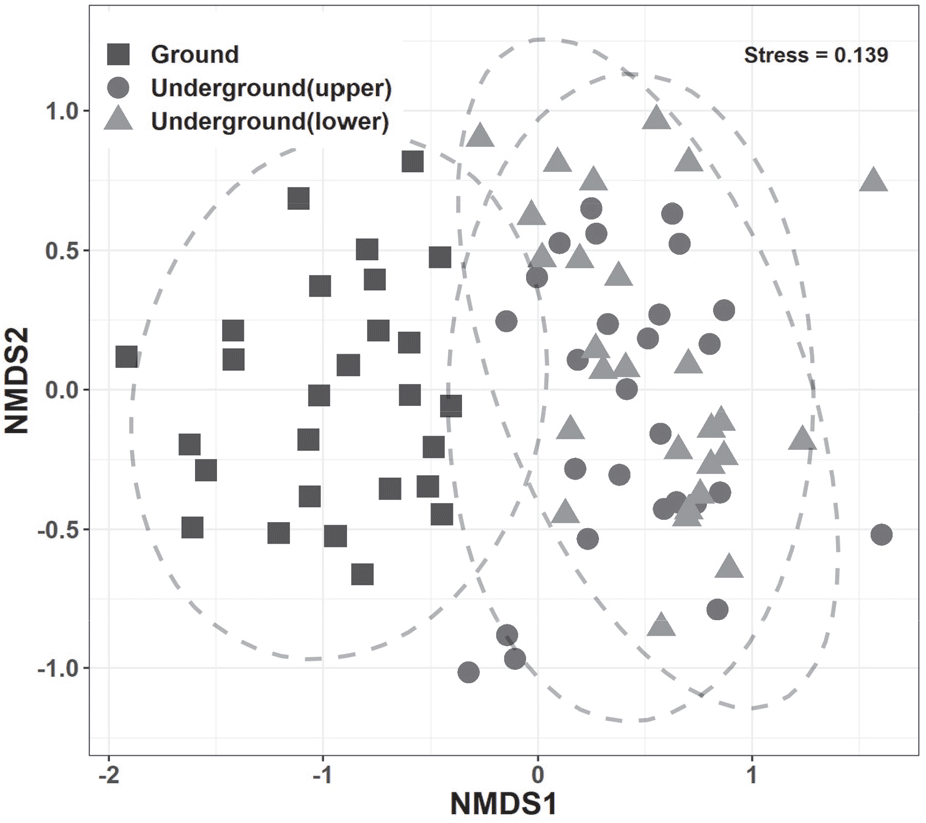
서열 분석 결과 1축의 좌측에서 우측으로 하층식생, 그리고 상토 및 하토 군집이 위치하였고, 상토 및 하토의 조사구는 서로 가까이 위치하고 퍼짐의 정도가 거의 유사하여 층위 내 이질성이 낮게 분석되었다(Figure 3). 매토종자 군집이 위치하는 1축의 우측에는 숲종인 태백제비꽃을 비롯하여 개망초, 주름조개풀, 그리고 여뀌류 등의 빈터종들이, 그리고 하측식생인 1축의 좌측에는 붉나무, 작살나무, 병꽃나무, 대사초 등으로 주로 숲종 및 가장자리종들이었다(종 서열 결과는 제외하였음).
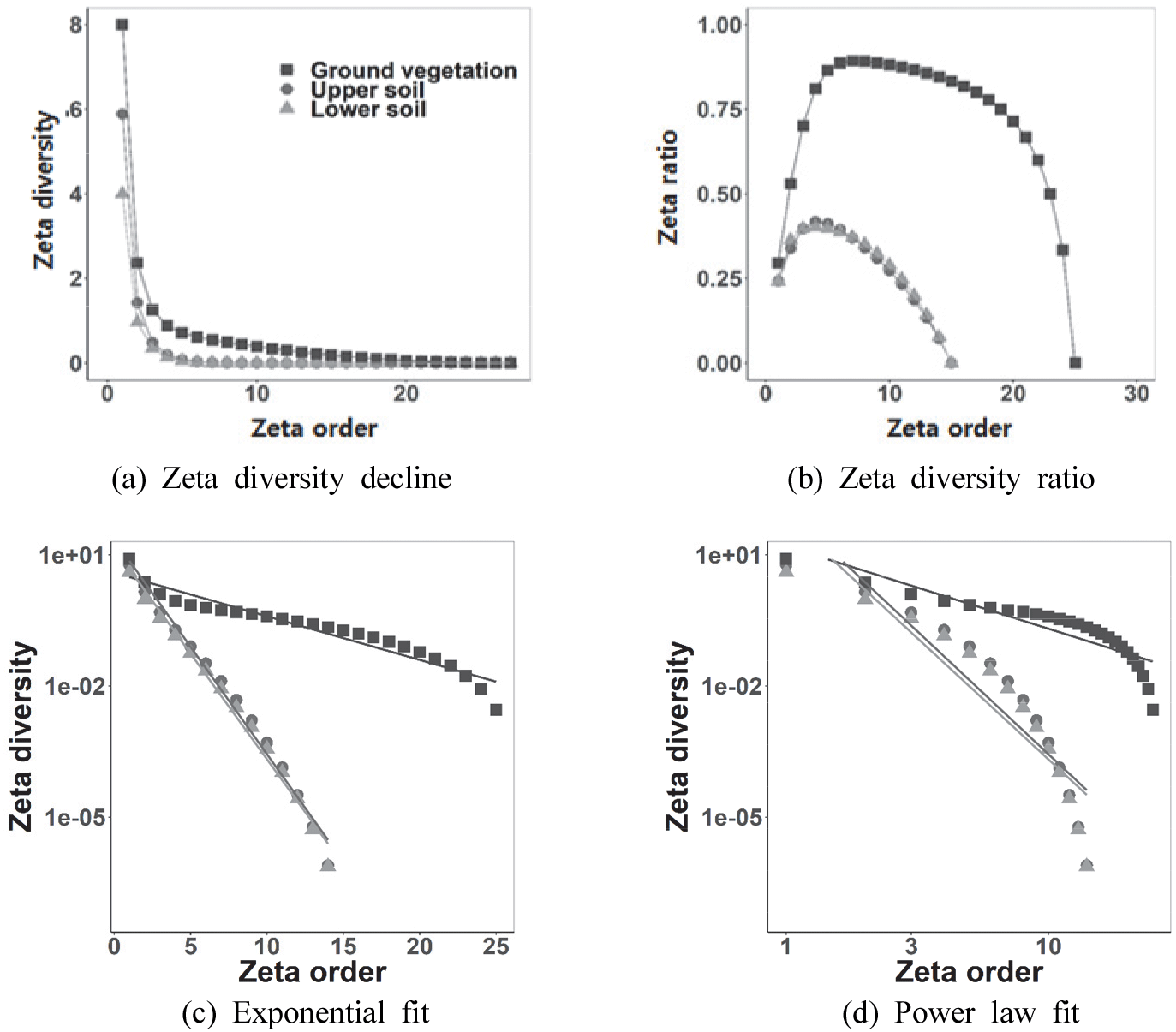
제타다양성 감소는 세 식생 층위 모두 유사한 형태로 진행되었다[Figure 4(a)]. 제타 2 순차에서 하토가 상토보다 적은 공통 출현종의 감소가 관찰되었고, 제타 3 순차에서는 거의 동일한 값을 보였다. 상토와 하토에서 제타 비율은 유사하게 증가한 후 감소하여, 군집 내 희귀종과 일반종의 기여도가 동일함을 보였다[Figure 4(b)]. 제타 다양성 변화 양상에 대한 지수 및 멱함수 적합은 지상 및 지하 식생 모두 지수 함수로 적합되어 환경 여과 및 지위 구조에 따른 상호작용 영향 보다는 임의성이 높은 특성을 보였다[Figure 5(c) 및 5(d)]; 지수함수와 멱함수의 AIC 값은 하층식생에서 각각 0.9와 23.2이었고, 상토에서 17.7과 48.8이었으며, 하토에서 15.1과 47.6이었다.
고 찰
본 연구는 광릉숲 토양에서 10 cm 간격의 상토(0~5 cm) 및 하토(15~20 cm), 두개 깊이에 대한 매토종자 군집의 변화를 다루었다. 광릉숲에 위치한 묵논에서 천이 진행에 따른 지상 및 지하 식물 군집 사이의 이질성의 증가가 보고된 바 있다(Cho et al., 2018). 이러한 매토종자 군집의 특성은 우리 연구 장소 같은 성숙림에서 지상부 식생으로부터 토양 내부로의 종자 이입이 제한적임을 말한다. 따라서 우리의 결과는 제한적인 종자 이입을 통한 토양 내 수직이동에 따른 매토종자 군집의 변화 과정을 나타내고 있다. 물론 종자의 수직이동에 영향을 줄 수 있는 깊이에 따른 토양의 물리성, 그리고 종자 쇠락(seed depletion)에 영향을 줄 수 있는 pH 변화 같은 화학성에 대한 자료는 부족하다. 이러한 한계가 존재하지만, 지역의 종급원(Species pool)으로서 매토종자의 보전 기능을 고려하면(Vandvik et al., 2016), 토양 내 식물 군집 특성의 일반성 및 지역성에 대한 다양한 접근이 필요하다. 예를 들면, 천이 진행과 다양한 토양 깊이 구배에 따른 매토종자 군집의 변화 양상에 대한 접근이 있을 수 있다. 그리고 또한 식생 복원 또는 보전 역량 관점에서, 군집 조합 및 해체 과정에 대한 지식 수준을 향상시켜야 한다(Helsen et al., 2015).
광릉숲의 매토종자는 목본 식물의 다양성 및 발아 밀도가 높았고, 빈터종보다는 가장자리 종이 풍부하였다. 일반적으로 숲 식생에서 매토종자는 목본 식물의 비율은 현저히 낮은데, 목본성 식물의 종자는 크고 무거우며 주로 낙엽층에 분포하기 때문이다(Marquis, 1975; Graber and Thompson, 1978; Schiffman and Johnson, 1992). 또한 수관이 닫힌 구조를 가진 산림은 매토종자보다 치묘 발아(seedling banks)를 통해 개체를 공급한다(Fröborg and Eriksson, 1997). 빈터종은 현재 또는 과거 숲에 성숙한 식물(mature plants)이 존재하지 않고도 매토종자에 포함되는 것이 가능하다(Peterson and Carson, 1996). 우리의 결과에서 외래종은 바람에 분산되는 특성을 지닌 3종(Erigeron annuus, Conyza canadensis, Erechtites hieracifolia)이 6개 소방형구에서 1개씩 출현하였다. 오랜 기간 보존 관리된 광릉숲의 매토종자에서 외래식물 밀도는 일반적으로 매우 낮았는데, 성숙림에서의 특성으로 이해할 수 있다(Leckie et al., 2000).
광릉숲의 성숙림에서 토양 깊이 증가에 따른 매토종자 군집의 다양성과 발아 밀도는 감소하는 경향이었지만 교목 집단을 제외하면 통계적으로 유의성은 없었다. 그리고 상토와 하토 군집의 종자 형질 역시 뚜렷한 차이가 없었다. 연구에 적용된 하토의 깊이는 15 cm~20 cm로서, 20 cm 이내 수준에서는 뚜렷한 토양 식물군집의 쇠락이 관찰되지 않을 수 있다. 매토종자 군집의 구조와 동태는 지상 식생과 비교를 포함하여 다양한 수준(예, 교란 후 천이 및 개별 생태계)에서 진행되어 왔다. 일반적으로 매토종자 군집에서 종다양성과 발아 개체의 밀도는 천이 진행(Royo and Ristau, 2013) 및 토양 깊이 증가(Bossuyt et al., 2002; Godefroid et al., 2006)에 따라 감소하는 경향이 알려져 있다. 스텝 기후에서 강한 바람의 영향을 많이 받는 사구 환경에서 불안정한 토양 환경이 매토종자의 풍부성에 영향을 주는 것이 보고되었다(Qian et al., 2016). 온대지역에서 미세부터 일정 규모 이상 크기의 교란으로서 서릿발(frost heaving)은 불안정한 토양 환경을 형성한다(Groffman et al., 2011). 대륙성 기후의 저지대에 위치하여 비교적 눈이 적게 내리거나, 내리더라도 빠르게 녹는 광릉숲에서 숲 내 서릿발은 일반적이며, 광범위한 현상으로 관찰된다. 따라서 낙엽층 및 부식층 아래에서 서릿발 현상에 따른 토양 불안정성과 뒤따른 종자들의 수직 이동은 연구에 적용된 상토와 하토 사이의 낮은 군집 이질성의 배경일 수 있다.
본 연구의 결과에서 매토종자 군집의 쇠락은 뚜렷하지 않았으며, 관련된 하토 내 종조성의 단순화 역시 관찰되지 않았다. 종에 따른 종자 수명, 그리고 토양 깊숙히 이동하는데 관여하는 종자의 크기 같은 결정론적 요인이 중요하다면, 종자의 생태 형질과 토양의 여과 기능에 의해 하토의 종조성은 상토보다 낮은 변이성을 보였을 것이다(Royo and Ristau, 2013). 상토와 하토에서, 동일한 제타다양성 감소 양상은 구성 종의 생태 형질뿐만 아니라 다양성의 구조화 양상 역시 유사하다는 것을 나타냈다. 이에 더하여, 활용된 토양 층위의 개별 군집은 서로 독립적 양상으로서 멱함수 보다는 지수함수의 적합도가 높아 확률론적 과정이 우세함을 보였다.
군집 구조와 종조성 변화와 관련한 다양한 생태 기작들은 주로 군집 내에서 발생하는 상호작용을 포함하는 결정론적 과정에 대한 연구들이 많았다. 그러나 우점하는 생태학적 기제(결정론 및 확률론)의 시공간 상 변동은 종 교대와 뒤따른 군집 구조 및 다양성 변화에 대한 다양한 접근을 필요로 하고 있다(Goldberg, 1987; Bullock et al., 1995). 앞서 언급한 바와 같이, 매토종자 군집의 쇠락 경향성은 종자 수명에 영향을 주는 크기와 무게의 결정론적 과정이 중요하게 지적되어 왔다(Schiffman and Johnson, 1992). 이와 더불어 지상과 지하 식물 군집에서 종조성의 이질성 증가와 함께 결정론과 확률론적 과정의 상대적 중요성 변화가 교란 후 매토종자 군집의 천이에서 관찰되고 있다(Royo and Ristau, 2013). 우리의 결과는 활용된 20 cm 이내의 토양 내의 종조성은 비교적 적은 변화와 더불어 종자 형질보다는 이입 및 소실의 임의 과정(stochastic processes)이 우점하는 것으로 평가할 수 있다.
결 론
종급원으로서 매토종자 군집의 생태 구조 및 다양성에 기초한 기능 평가는 지역의 다양성 이해와 보전, 그리고 군집 조합 및 해체 과정에 대한 높은 이해도를 요구하는 복원생태학의 관점에서 중요한 학술 및 실행 지식을 쌓을 수 있는 활동이다. 광릉숲의 매토종자는 초본의 구성 비율이 낮고 목본 식물의 다양성 및 발아 밀도가 높았고, 빈터종보다는 가장자리 종이 풍부하였다. 토양 깊이 증가에 따른 매토종자 군집 변화는 뚜렷하지 않았지만, 감소 추세가 관찰되었다. 연구에 적용된 토양 깊이(20 cm 이내)에서는 교목 식물의 풍부도만이 뚜렷한 감소를 나타냈다. 적용된 토양 층위의 종조성과 다양성은 확률론적 생태 과정이 우점하는 것이 확인되었다. 이러한 우점하는 생태 과정은 시공간적 변이를 나타내므로 매토종자 군집 조합에 대한 구조화 양상의 변화 이해에 필요한 다양한 활동이 필요하다. 주요 및 구성 종들의 변화 기작에 대한 충실한 이해는 종 및 식생의 보전, 그리고 복원 활동의 핵심 지식일 것이다. 예를 들어, 교란 후 군집 조합의 초기 조건에서 결정론적 또는 확률론적 과정의 상대적 중요성과 변화 양상에 대한 지식은 식생 복원에 필요한 재료의 준비와 종 도입(식재) 방식 같은 핵심 영역에 중요한 정보를 제공한다.