



서 론
우리 조상들은 산을 물줄기처럼 끊이지 않는 맥으로 보았고, 산은 물을 낳고 물은 산을 가르지 않는다고 여겼다. 이것이 산줄기를 인식하는 기본개념이었고 이것을 체계화 한 것이 백두대간이다(Korea Forest Service, 2006). 백두대간이란 개념은 신라말엽부터 고려초기의 옥룡기에서 시작이 되어 조선시대에 완성되었으며, 백두산에서 시작되어 원산, 낭림산, 금강산, 설악산, 오대산, 태백산, 속리산, 장안산을 거쳐 지리산까지 약 1,400 km에 이르고 우리나라의 큰 산줄기를 형성하면서 물줄기를 동서로 가르는 국토의 골간을 말한다(Shin, 2004; Hwang et al., 2015; Chae and Yun, 2019; Song et al., 2019b). 이렇듯 백두대간은 우리나라에서 역사적 의미가 강한편이며 동고서저의 지리·지형적 위치 때문에 한반도 전체의 생태계권 생물 군집의 발달과 퇴행의 과정을 거치면서 생태학적으로 특산식물이나 희귀 및 멸종위기종 등 다양한 동·식물의 서식처로서 생태적 가치 측면에서도 중요한 의미를 가진다(Korea Forest Service, 2003).
이러한 중요성이 국민들에게 알려지면서 백두대간에 관심이 부각되어 향로봉에서 지리산까지 약 701 km에 대하여 백두대간보호에 관한 법률에 의해 보호지역으로 재정되어 보호받고 있다. 하지만 최근 산림치유, 휴양 및 여가의 열풍이 일어나 백두대간을 찾는 탐방객이 급증하였고, 이에 따라 인위적 교란과 식생경관의 훼손이 심각해져 산림복원 대책이 시급한 실정이다(Kim et al., 2018).
일반적으로 산림생태계를 체계적으로 보전하고 복원하기 위해서는 대상 생태계의 구성적 기반이 되는 식생에 대한 정확한 정보수집이 요구되며(Kim et al., 2018), 정보를 바탕으로 객관적인 보전 및 관리 계획 수립을 위한 식생에 대한 분류를 통해 정보를 수집하게 된다(Park et al., 2009). 산림식생에 대한 분류는 종조성에 의한 식물사회학적 측면을 중요시하는 Z-M방식이 가장 널리 사용되고 있지만(Son et al., 2016, Kim et al., 2017), 백두대간 마루금의 식생을 대상으로 통계패키지를 이용한 군집분석이 주를 이루고 있었고, Z-M학파의 전통적 분류법에 의한 식생유형분류는 미흡한 실정이다(Song and Yun, 2019).
백두대간을 마루금의 식생을 대상으로 식물사회학적 유형분류를 한 연구는 피재-도래기재구간(Oh and Park, 2002), 정령치-복성이재구간(Choi and Oh, 2003), 남덕유산-소사재구간(Choi et al., 2004), 부봉-포암산구간(Choo and Kim, 2005), 댓재-백봉령구간(Lee et al., 2012), 구룡령-약수산구간(An et al., 2014), 조침령-신배령구간(Lee et al., 2014), 중대사-비로봉구간(Han et al., 2015), 단목령-구룡령구간(Kim et al., 2018), 미시령-단목령구간(Chae and Yun, 2019), 깃대배기봉-죽령구간(Song and Yun, 2019), 닭목령-댓재구간(Song et al., 2019b)등 세부구간에 대하여 여러 연구들이 이루어져 왔지만 하나의 권역을 대상으로 한 연구는 미흡하였다.
따라서 본 연구에서는 백두대간의 5개 권역(설악산권역, 태백산권역, 속리산권역, 덕유산권역, 지리산권역) 중 입지환경 특성에 따라 다양한 식생이 나타나는 덕유산이 포함된(Kim et al., 2010) 덕유산권역(늘재~육십령 구간) 마루금 주변의 산림식생들 대상으로 식물사회학적 식생유형분류를 통해 산림군집을 분류하고, 분류된 식생단위를 바탕으로 식생단위별 중요치, 종다양도를 산출하여 향후 체계적인 보전 및 관리를 위한 생태적 정보를 제공하는데 목적이 있다.
연구방법
본 연구는 남한의 백두대간 구간 중 경상북도 상주시에 속해 있는 청화산(970 m)과 속리산 문장대(1,031.7 m)를 가로질러 32번 국도가 지나는 늘재에서 경상남도 함양군에 속해있는 할미봉(1,026 m)과 전라북도 장수군에 속해있는 깃대봉(1,015 m) 사이를 가로질러 26번국도가 지나는 육십령까지 약 154 km에 해당하는 백두대간 덕유산권역 마루금을 연구대상지로 선정하였다.
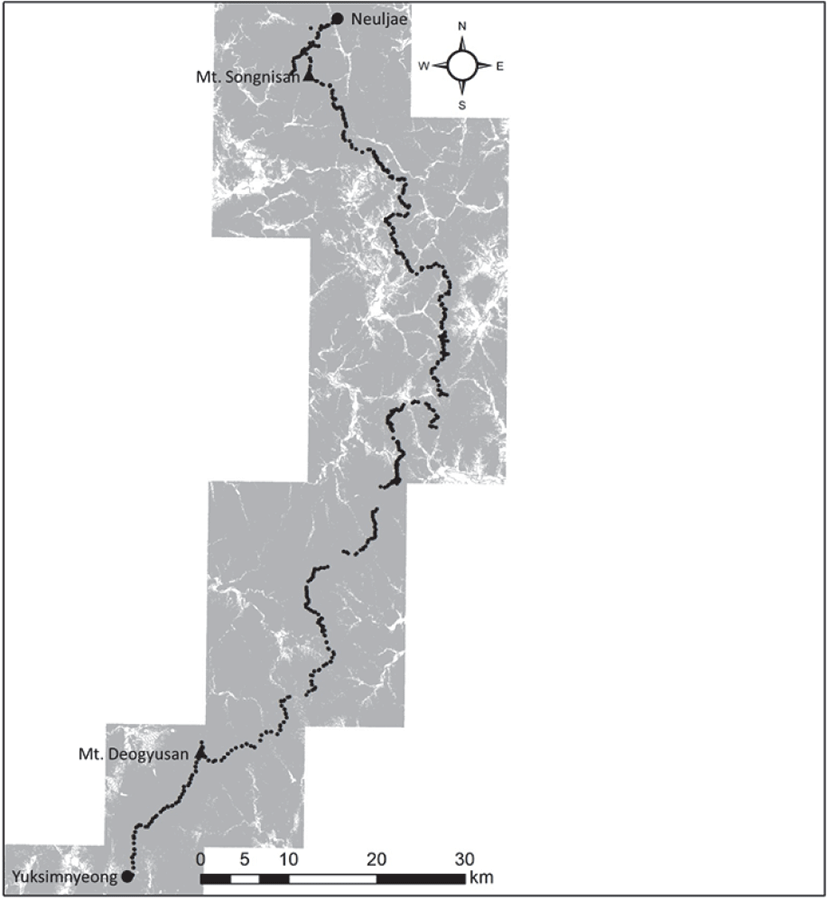
백두대간은 동사면 방향으로는 경사가 급한 해양성 기후이나 서사면 방향으로는 내륙성 기후로 완만한 경사를 이루고 있으며, 연평균 기온은 6.3°C~12.7°C, 연평균 강수량은 1,091.6 mm~1,985.1 mm으로 나타났다(ME, 2002, Song et al., 2019b). 남한지역을 대상으로 백두대간 마루금은 수치표고모델(digital elevation model; DEM) 분석에 근거한 해발고도는 최저 200 m에서 최대 1,909 m에 이르는 것으로 보고되었다(Lee et al., 2013, Song et al., 2019b). 본 조사지의 생태권역은 산악권역과 남서산야권역에 속하며(Shin and Kim, 1996), 식물구계도에서는 중부아구와 남부아구에 속한다(Lee and Yim, 2002).
본 연구는 2019년 5월부터 10월까지 백두대간 늘재에서 육십령 구간의 마루금을 대상으로 식생의 구조를 파악하기 위해 최대한 정량적인 식생자료를 수집하기 위하여 500 m 간격으로 마루금선 좌우 50 m를 이격하여 20 m × 20 m의 총 637개소의 방형구를 Z-M학파의 식물사회학적 방법(Ellenberg, 1956; Braun-Blanquet, 1964)에 따라 해발, 지형, 생태적 밀도 등 여러 입지환경요인을 고려하여 선정하였다. 식생조사법에 따라 조사구내에 출현하는 모든 식물종의 양과 생육상태에 대한 측정을 실시하였고, 양적인 측면은 표본조사구에서 출현하는 모든 종의 피도(Coverage)와 개체수(Abundance)를 조합시킨 우점도 계급(Braun-Blanquet, 1964)을 판정하여 기록하였으며, 생육상태는 종 개체의 집합 혹은 이산의 정도에 따른 군도(Sociability)를 판정하여 기록하였다. 구성종의 식물분류와 동정은 원색한국수목도감(Hong et al., 1987), 원색식물도감(Lee, 2003), 나무생태도감(Yun, 2016)을 기준으로 실시하였으며, 종명의 학명과 국명은 APG(Angiosperm Phylogeny Group)Ⅳ 시스템이 적용된 국가표준식물목록(Korea Forest Service, 2021)을 기준으로 작성하였다.
늘재 ~ 육십령 구간에서 조사된 637개소의 방형구에 대한 식생유형 분류는 대별종군과 식별종군의 용이한 파악을 위하여 PC-ORD v7.04 프로그램을 통해 정량적인 방법인 Hill(1979)의 TWINSPAN(Two-Way Indicator Species Analysis)과 정성적인 방법인 Ellenberg(1956)의 표조작법(Tabulation method)을 이용하였다. 식생유형분류를 통해 산출된 식생단위(Vegetation unit)의 층위별 점유율을 파악하기 위하여 Curtis and McIntosh(1951)의 방법을 이용하여 상대빈도(RF), 상대피도(RC)를 합산한 중요치(I.V: Importance Value)를 산출하였고, 또한 각 층위에 생육하는 수종들에 대하여 개체크기를 고려해 교목층 3, 아교목층 2, 관목층 1(Yim et al., 1980), 초본층 0.5의 가중치를 부여한 평균상대우점치(M.I.P: Mean Importance Value)를 산출하였다. 일정 면적 내 종수 또는 군집 내 일정수의 개체를 구성하는 종수를 의미하는 종풍부도와 각 종에 속하는 개체수가 고르게 분포하는가를 나타내는 균등도(Species eveness)를 동시에 나타내는 종다양도지수(Shannon and Weaver, 1949), 최대종다양도지수, 종균재도, 종우점도를 분석하였다(Brower and Zar, 1977).
결과 및 고찰
백두대간(늘재 ~ 육십령) 마루금 637개소의 산림식생조사 자료를 바탕으로 Z-M학파의 식물사회학적 방법에 따라 식생유형분류를 수행한 결과는 다음과 같다. 최상위 수준에서는 신갈나무군락군(Quercus mongolica community group)이 분류되었고, 신갈나무군락군은 구상나무군락(Abies koreana community), 산오이풀군락(Sanguisorba hakusan-ensis community), 여뀌군락(Persicaria hydropiper community), 굴참나무군락(Q. variabilis community), 떡갈나무군락(Q. dentata community), 층층나무군락(Cornus controv-ersa community), 신갈나무전형군락(Q. mongolica typical community)으로 세분되었다. 구상나무군락은 퍼진고사리군(Dryopteris expansa group), 가문비나무군(Picea jezoensis group)으로 세분되었고, 굴참나무군락은 떡갈나무군(Q. dentata group), 굴참나무전형군(Q. variabilis typical group)으로 세분되었다. 조사지역 내 신갈나무군락군은 총 7개의 군락, 4개 군의 분류체계를 가졌고, 9개의 식생단위로 분류되었다. 신갈나무는 우리나라 활엽수림 중 가장 많이 분포하고 있는 수종으로 보고되었으며(Lee et al., 1990, Lee et al., 1998, Yun et al., 2011), 백두대간 내에서도 가장 많은 점유면적을 차지하는 종으로 보고된 바가 있어(Chung, 1998, Cho et al., 2004, Lee et al., 2014) 본 연구와 일치한 것으로 사료되었다.
본 식생단위는 신갈나무군락군에서 종군2의 구상나무, 시닥나무의 식별종 출현에 의해 구상나무군락으로 구분되어졌으며, 구상나무군락의 하위종군으로 퍼진고사리, 나래회나무, 모시대, 박새, 각시서덜취의 식별종에 의해 퍼진고사리군으로 세분되었다. 본 식생단위에 출현하는 식물 중 상재도가 Ⅲ 이상인 종은 신갈나무, 철쭉, 가는잎그늘사초, 실새풀, 구상나무, 퍼진고사리, 나래회나무 등 총 16종이 출현하였고, 우점도가 3이상인 종은 신갈나무 철쭉, 가는잎그늘사초, 실새풀, 구상나무 등 총 17종이 출현하였다. 무생물적 환경요인은 평균해발고 1,383.6 m, 평균사면경사도 20.0°, 평균암석노출도 37.9%로 나타났으며, 전체 식생단위 중 가장 높은 평균암석노출도로 나타났다. 생물적환경요인은 교목층 평균식피율 60.0%, 아교목층 평균식피율 37.0%, 관목층 평균식피율 46.9%, 초본층 평균식피율 62.5%, 교목층 평균수고 7.9 m, 아교목층 평균수고 5.6 m, 관목층 평균수고 2.4 m, 초본층 평균수고 0.7 m, 교목층 평균흉고직경 23.3 cm, 아교목층 평균흉고직경 13.3 cm, 관목층 평균흉고직경 4.8 cm, 평균출현종수 24.0종으로 나타났다. 본 연구에서 구상나무가 군락단위로 분류된 것은 백두대간 늘재~육십령의 구간 사이에 구상나무가 자생하는 덕유산 향적봉과 중봉구간이 있기 때문이며, 다른 식생단위에서는 구상나무가 출현되지 않았다. 구상나무는 지리산, 한라산, 덕유산, 가야산 등 남부지방의 해발 1,000 m 이상의 아고산 지대에 분포하고 있는 한국 특산종으로 보고된 바(Hong and Lee, 1995, Song et al., 2019a) 본 연구와 유사한 것으로 사료되었다.
본 식생단위는 신갈나무군락군에서 종군2의 구상나무, 시닥나무의 식별종 출현에 의해 구상나무군락으로 구분되어졌으며, 구상나무군락의 하위종군으로 가문비나무, 각시취의 식별종에 의해 가문비나무군으로 세분되었다. 본 식생단위에 출현하는 식물 중 상재도가 Ⅲ 이상인 종은 철쭉, 가는잎그늘사초, 실새풀, 구상나무, 가문비나무, 산오이풀, 새, 고본, 개시호, 바위채송화, 범꼬리, 털진달래, 동자꽃, 붉은병꽃나무, 곰취 등 총 28종이 출현하였고, 우점도가 3이상인 종은 철쭉, 구상나무, 가문비나무, 새, 털진달래, 사스래나무 등 총 12종이 출현하였다. 무생물적 환경요인은 평균해발고 1,516.3 m, 평균사면경사도 16.7°, 평균암석노출도 36.7%로 나타났으며, 전체 식생단위 중 가장 높은 평균해발고로 나타났다. 생물적환경요인은 교목층 평균식피율 55.0%, 아교목층 평균식피율 15.0%, 관목층 평균식피율 46.0%, 초본층 평균식피율 89.0%, 교목층 평균수고 8.0 m, 아교목층 평균수고 7.0 m, 관목층 평균수고 1.6 m, 초본층 평균수고 0.5 m, 교목층 평균흉고직경 24.0 cm, 아교목층 평균흉고직경 22.0 cm, 관목층 평균흉고직경 3.0 cm, 평균출현종수 27.8종으로 나타났다. 가문비나무는 우리나라에서 계방산, 덕유산, 지리산에만 자생하고 있는 것으로 보고되어 있고(Han et al., 2016), 본 연구에서 군단위로 분류된 것은 백두대간 늘재~육십령의 구간사이에 가문비나무가 자생하는 덕유산 향적봉과 중봉구간이 있기 때문이며, 다른 식생단위에서는 가문비나무가 출현되지 않았다. Han et al.(2016)은 덕유산 구간의 가문비나무군락은 해발 1,444 m~1,554 m에 출현하는 것으로 보고 하였으며 이는 본 연구와 유사한 것으로 사료되었다.
본 식생단위는 신갈나무군락군에서 종군5의 산오이풀, 새, 고본, 개시호, 바위채송화, 돌양지꽃, 범꼬리, 바위구절초, 산부추의 식별종 출현에 의해 산오이풀군락으로 구분되어졌다. 본 식생단위에 출현하는 식물 중 상재도가 Ⅲ 이상인 종은 신갈나무, 철쭉, 가는잎그늘사초, 산오이풀, 새, 고본, 털진달래, 동자꽃, 붉은병꽃나무 등 총 19종이 출현하였고, 우점도가 3이상인 종은 신갈나무, 철쭉, 가는잎그늘사초, 실새풀, 새, 털진달래, 사스래나무 등 총 18종이 출현하였다. 본 식생단위에서 새는 Ⅲ이상의 상재도와 3이상의 우점도로 나타났는데 Song and Yun(2019)은 소백산 비로봉 초원의 새는 약 30년이 지난 후 현존식생에서 기존에 우점하던 식생보다 더 우세한 것으로 보고하였다. 따라서 본 연구에서도 산오이풀과 새의 특징종 세력변화에 대한 모니터링이 필요할 것으로 사료되었고, 본 식생단위의 식생이 주로 자생하는 덕유산 향적봉 및 제2덕유산 지역의 경관생태학적 장기적인 모니터링이 필요할 것으로 판단되었다. 또한 Kim et al.(2011)은 덕유산국립공원의 현존식생분포현황에서 아고산식생 중 초원을 하나의 유형으로 분류하였으며 이는 본 연구와 유사한 것으로 사료되었다. 무생물적 환경요인은 평균해발고 1,360.7 m, 평균사면경사도 16.5°, 평균암석노출도 21.5%로 나타났다. 생물적환경요인은 교목층 평균식피율 78.8%, 아교목층 평균식피율 25.0%, 관목층 평균식피율 51.3%, 초본층 평균식피율 68.1%, 교목층 평균수고 5.8 m, 아교목층 평균수고 5.2 m, 관목층 평균수고 1.9 m, 초본층 평균수고 0.6 m, 교목층 평균흉고직경 12.6 cm, 아교목층 평균흉고직경 9.0 cm, 관목층 평균흉고직경 2.7 cm, 평균출현종수 25.9종으로 나타났다.
본 식생단위는 신갈나무군락군에서 종군6의 여뀌, 콩제비꽃, 환삼덩굴, 개망초, 사위질빵, 영아자, 호두나무의 식별종 출현에 의해 여뀌군락으로 구분되었다. 본 식생단위에 출현하는 식물 중 상재도가 Ⅲ 이상인 종은 여뀌, 콩제비꽃, 환삼덩굴, 개망초, 사위질빵, 일본잎갈나무, 주름조개풀 등 총 23종이 출현하였고, 우점도가 3이상인 종은 여뀌, 호두나무, 소나무, 일본잎갈나무, 상수리나무 등 총 10종이 출현하였다. 본 식생단위에서 호두나무와 상수리나무가 높은 우점도로 출현하였고, 개망초, 망초, 달맞이꽃, 닭의덩굴, 방가지똥, 털별꽃아재비등의 귀화식물등이 출현하는 것을 고려한 결과 본 식생단위는 민가인근의 농경지식생이 하나의 단위로 구분된 것으로 사료되었다. 무생물적 환경요인은 평균해발고 754.8 m, 평균사면경사도 11.0°, 평균암석노출도 5.8%로 나타났다. 생물적환경요인은 교목층 평균식피율 73.3%, 아교목층 평균식피율 25.0%, 관목층 평균식피율 24.0%, 초본층 평균식피율 56.7%, 교목층 평균수고 16.3 m, 아교목층 평균수고 8.2 m, 관목층 평균수고 2.7 m, 초본층 평균수고 0.4 m, 교목층 평균흉고직경 25.3 cm, 아교목층 평균흉고직경 11.0 cm, 관목층 평균흉고직경 3.8 cm, 평균출현종수 37.0종으로 나타났으며, 전체 식생단위 중 평균출현종수가 가장 높은 것으로 나타났다.
본 식생단위는 신갈나무군락군에서 종군7의 굴참나무, 소나무의 식별종 출현에 의해 굴참나무군락으로 구분되었고, 굴참나무군락의 하위종군으로 떡갈나무, 밤나무, 아까시나무의 식별종에 의해 떡갈나무군으로 세분되었다. 본 식생단위에 출현하는 식물 중 상재도가 Ⅲ 이상인 종은 신갈나무, 가는잎그늘사초, 굴참나무, 소나무, 떡갈나무, 밤나무, 산초나무 등 총 7종이 출현하였고, 우점도가 3이상인 종은 신갈나무, 철쭉, 가는잎그늘사초, 굴참나무, 소나무, 떡갈나무, 밤나무, 아까시나무, 일본잎갈나무, 상수리나무, 산초나무 등 총 36종이 출현하였다. 무생물적 환경요인은 평균해발고 479.8 m, 평균사면경사도 18.3°, 평균암석노출도 2.3%로 나타났으며, 전체 식생단위 중 가장 낮은 평균해발고로 나타났다. 생물적환경요인은 교목층 평균식피율 75.4%, 아교목층 평균식피율 22.2%, 관목층 평균식피율 35.9%, 초본층 평균식피율 29.0%, 교목층 평균수고 13.7 m, 아교목층 평균수고 6.6 m, 관목층 평균수고 1.7 m, 초본층 평균수고 0.5 m, 교목층 평균흉고직경 21.3 cm, 아교목층 평균흉고직경 7.9 cm, 관목층 평균흉고직경 2.3 cm, 평균출현종수 18.3종으로 나타났다.
본 식생단위는 신갈나무군락군에서 종군7의 굴참나무, 소나무의 식별종 출현에 의해 굴참나무군락으로 구분되었고, 종군7 이하의 식별종들이 출현하지 않음으로 인하여 굴참나무전형군으로 세분되었다. 본 식생단위에 출현하는 식물 중 상재도가 Ⅲ 이상인 종은 신갈나무, 철쭉, 가는잎그늘사초, 굴참나무, 소나무 등 총 6종이 출현하였고, 우점도가 3이상인 종은 신갈나무, 철쭉, 가는잎그늘사초, 실새풀, 굴참나무, 소나무, 일본잎갈나무, 상수리나무, 산초나무 등 총 47종이 출현하였다. 무생물적 환경요인은 평균해발고 605.1 m, 평균사면경사도 20.3°, 평균암석노출도 6.5%로 나타났다. 생물적환경요인은 교목층 평균식피율 72.5%, 아교목층 평균식피율 27.1%, 관목층 평균식피율 39.0%, 초본층 평균식피율 26.3%, 교목층 평균수고 13.2 m, 아교목층 평균수고 6.3 m, 관목층 평균수고 1.8 m, 초본층 평균수고 0.4 m, 교목층 평균흉고직경 22.4 cm, 아교목층 평균흉고직경 8.5 cm, 관목층 평균흉고직경 2.5 cm, 평균출현종수 16.0종으로 나타났다. Kim et al.(2011)은 덕유산국립공원 낙엽활엽수림에서 굴참나무는 해발 600 m~900 m의 건조한 사면에 분포한다고 하였으며 이는 본 연구와 유사한 것으로 사료되었다.
본 식생단위는 신갈나무군락군에서 종군8의 떡갈나무, 밤나무, 아까시나무의 식별종 출현에 의해 떡갈나무군락으로 구분되었다. 본 식생단위에 출현하는 식물 중 상재도가 Ⅲ 이상인 종은 신갈나무, 가는잎그늘사초, 떡갈나무, 밤나무, 주름조개풀 등 총 8종이 출현하였고, 우점도가 3이상인 종은 신갈나무, 철쭉, 가는잎그늘사초, 떡갈나무, 밤나무, 아까시나무, 일본잎갈나무, 주름조개풀, 상수리나무, 산초나무 등 총 35종이 출현하였다. 무생물적 환경요인은 평균해발고 480.2 m, 평균사면경사도 16.2°, 평균암석노출도 1.9%로 나타났다. 생물적환경요인은 교목층 평균식피율 79.8%, 아교목층 평균식피율 26.2%, 관목층 평균식피율 36.0%, 초본층 평균식피율 27.6%, 교목층 평균수고 14.4 m, 아교목층 평균수고 6.8 m, 관목층 평균수고 1.8 m, 초본층 평균수고 0.5 m, 교목층 평균흉고직경 21.1 cm, 아교목층 평균흉고직경 8.3 cm, 관목층 평균흉고직경 2.1 cm, 평균출현종수 18.7종으로 나타났다. Lee et al.(2010)은 떡갈나무가 전국 표고 800 m 이하의 산기슭, 산중턱에서 자생하고 떡갈나무의 최적 수직분포대는 300 m로 보고하였으며, 이는 본 연구와 유사한 것으로 사료되었다.
본 식생단위는 신갈나무군락군에서 종군9의 층층나무, 작살나무, 십자고사리의 식별종 출현에 의해 층층나무군락으로 구분되었다. 본 식생단위에 출현하는 식물 중 상재도가 Ⅲ 이상인 종은 신갈나무, 층층나무, 작살나무 등 총 10종이 출현하였고, 우점도가 3이상인 종은 신갈나무, 철쭉, 가는잎그늘사초, 층층나무, 작살나무, 일본잎갈나무, 주름조개풀, 상수리나무 등 총 43종이 출현하였다. 무생물적 환경요인은 평균해발고 833.9 m, 평균사면경사도 20.3°, 평균암석노출도 12.4%로 나타났다. 생물적환경요인은 교목층 평균식피율 78.3%, 아교목층 평균식피율 35.0%, 관목층 평균식피율 36.8%, 초본층 평균식피율 48.8%, 교목층 평균수고 13.6 m, 아교목층 평균수고 6.7 m, 관목층 평균수고 2.0 m, 초본층 평균수고 0.6 m, 교목층 평균흉고직경 25.9 cm, 아교목층 평균흉고직경 9.2 cm, 관목층 평균흉고직경 2.8 cm, 평균출현종수 22.1종으로 나타났다. 층층나무는 해발 1,300 m 이하의 계곡부에서 주로 분포하고 있다고 보고되었다(Masaki et al., 1994). 많은 백두대간의 연구에서 층층나무는 계곡부를 대표하는 식생으로 밝혀졌지만 백두대간 마루금 내의 층층나무군락에 대한 보고는 거의 이루어지지 않았고, 계곡부가 아닌 지형에서도 층층나무는 자생하고 있기에 백두대간 마루금내의 범주화 되지 않은 미소지형들에 대한 고찰이 필요할 것으로 사료되었다(Song et al., 2019b).
본 식생단위는 신갈나무군락군에서 종군1 이하의 종군에서 식별종들이 출현하지 않음으로 인하여 신갈나무전형군락으로 구분되었다. 본 식생단위에 출현하는 식물 중 상재도가 Ⅲ 이상인 종은 신갈나무, 철쭉, 가는잎그늘사초 등 총 7종이 출현하였고, 우점도가 3이상인 종은 신갈나무, 철쭉, 가는잎그늘사초, 실새풀, 아까시나무, 일본잎갈나무, 주름조개풀, 상수리나무 등 총 54종이 출현하였다. 무생물적 환경요인은 평균해발고 939.0 m, 평균사면경사도 18.9°, 평균암석노출도 10.3%로 나타났다. 생물적환경요인은 교목층 평균식피율 73.2%, 아교목층 평균식피율 33.2%, 관목층 평균식피율 42.1%, 초본층 평균식피율 48.4%, 교목층 평균수고 11.0 m, 아교목층 평균수고 6.3 m, 관목층 평균수고 2.0 m, 초본층 평균수고 0.6 m, 교목층 평균흉고직경 20.2 cm, 아교목층 평균흉고직경 9.6 cm, 관목층 평균흉고직경 2.9 cm, 평균출현종수 15.4종으로 나타났고, 전체 식생단위 중 평균출현종수가 가장 낮은 것으로 나타났다. 신갈나무전형군락은 산의 능선부와 사면중상부의 척박한 곳에 발달하여 특별한 식별종을 수반하지 않는다고 보고되었으며(Kim and Kil, 2000; Shin et al., 2011, Song et al., 2019b.) 이는 본 연구와 유사한 결과로 사료되었다.
백두대간 덕유산권역(늘재-육십령) 마루금의 산림식생을 식물사회학적 방법으로 분석하여 구분된 9개의 식생단위를 바탕으로 Curtis and McIntosh(1951)의 방법으로 식생단위별 중요치를 산출한 결과는 다음과 같다.
평균상대우점치는 신갈나무(27.1%)가 가장 높게 나타났으며 그 다음으로 구상나무(13.8%), 당단풍나무(12.8%), 가문비나무(4.8%) 순으로 각각 나타났다. 교목층 중요치는 신갈나무(41.3%)가 가장 높게 나타났으며 구상나무(26.6%), 가문비나무(10.3%), 잣나무(7.1%) 순으로 각각 나타났다. 아교목층 중요치는 당단풍나무(32.8%)가 가장 높게 나타났으며 신갈나무(25.2%), 까치박달(7.9%), 쇠물푸레나무(6.6%) 순으로 각각 나타났다. 관목층 중요치는 철쭉(17.2%)이 가장 높게 나타났으며 미역줄나무(17.1%), 당단풍나무(8.3%), 털진달래(4.9%) 순으로 각각 나타났다. 초본층 중요치는 조릿대(17.6%)가 가장 높게 나타났으며 미역줄나무(10.3%), 단풍취(8.5%), 실새풀(6.4%) 순으로 각각 나타났다.
평균상대우점치는 신갈나무(34.6%)가 가장 높게 나타났으며 그 다음으로 구상나무(18.2%), 잣나무(9.8%), 가문비나무(8.6%) 순으로 각각 나타났다. 교목층 중요치는 구상나무(38.8%)가 가장 높게 나타났으며 잣나무(21.3%), 가문비나무(18.6%), 사스래나무(12.5%) 순으로 각각 나타났다. 아교목층 중요치는 신갈나무(100.0%)가 나타났다. 관목층 중요치는 철쭉(19.9%)이 가장 높게 나타났으며 털진달래(19.4%), 쇠물푸레나무(10.0%), 사스래나무, 미역줄나무, 참싸리(8.0%) 순으로 각각 나타났다. 초본층 중요치는 새(16.7%)가 가장 높게 나타났으며 원추리(6.7%), 일월비비추(6.0%), 산오이풀(5.4%) 순으로 각각 나타났다. Ahn et al.(2010)은 지리산국립공원 천왕봉지역의 가문비나무림에서 구상나무와 잣나무의 상대우점치가 높다고 한 바 본 연구와 유사한 것으로 사료되었고, 아교목층에서는 신갈나무만 출현하는 것으로 보아 구상나무, 잣나무, 가문비나무와 지위경쟁이 일어날 것으로 사료되었다. 가문비나무와 구상나무는 기후변화 취약수종 및 희귀식물로(KFS, 2021) 가치가 매우 높아 지속적인 관찰이 필요하며 또한 초본층, 관목층의 중요치가 없거나 낮아 치수이입에 관한 모니터링도 필요할 것으로 사료되었다.
평균상대우점치는 신갈나무(36.1%)가 가장 높게 나타났으며 그 다음으로 당단풍나무(10.2%), 마가목(8.5%), 물푸레나무(8.2%) 순으로 각각 나타났다. 교목층 중요치는 신갈나무(63.1%)가 가장 높게 나타났으며 물푸레나무(16.6%), 참빗살나무(8.1%), 호랑버들(4.7%) 순으로 각각 나타났다. 아교목층 중요치는 마가목(27.6%)이 가장 높게 나타났으며 당단풍나무(25.6%), 신갈나무(18.8%), 쇠물푸레나무, 고로쇠나무, 피나무(9.4%) 순으로 각각 나타났다. 관목층 중요치는 철쭉(20.1%)이 가장 높게 나타났으며 털진달래(11.0%), 조록싸리, 미역줄나무(9.5%), 신갈나무(7.6%) 순으로 각각 나타났다. 초본층 중요치는 새(8.2%)가 가장 높게 나타났으며 가는잎그늘사초(5.8%), 실새풀(5.7%), 원추리(5.6%) 순으로 각각 나타났다.
*Other 359 companion species omitted
평균상대우점치는 일본잎갈나무(19.9%)가 가장 높게 나타났으며 그 다음으로 상수리나무(5.5%), 호두나무(5.5%), 당단풍나무(5.4%) 순으로 각각 나타났다. 교목층 중요치는 일본잎갈나무(43.0%)가 가장 높게 나타났으며 상수리나무, 호두나무(11.8%), 소나무, 거제수나무(9.3%), 층층나무, 벚나무류(7.3%) 순으로 각각 나타났다. 아교목층 중요치는 산뽕나무(17.3%)가 가장 높게 나타났으며 청단풍나무(13.9%), 느티나무, 떡갈나무(9.8%), 다래, 대왕참나무(8.3%) 순으로 각각 나타났다. 관목층 중요치는 생강나무(15.6%)가 가장 높게 나타났으며 고광나무(9.2%), 청단풍나무, 다래(7.4%), 층층나무(3.5%) 순으로 각각 나타났다. 초본층 중요치는 오미자(10.5%)가 가장 높게 나타났으며 여뀌(8.4%), 닭의장풀(5.0%), 이삭여뀌(3.1%) 순으로 각각 나타났다. 본 식생단위에서는 다른 식생단위와는 다르게 다래, 왕머루, 오미자 등의 덩굴성식물의 중요치가 높게 나타났는데, 이는 조림지 식생인 일본잎갈나무가 교목층에 우점하는 것으로 나타났으며, Byeon and Yun(2016)은 인공조림지가 동령림임으로 숲의 구조가 간단하며 수광량이 높아 덩굴성 식물의 이입률이 높아진다는 연구결과와 유사한 것으로 사료되었다(Kim, 2017).
평균상대우점치는 굴참나무(12.0%)가 가장 높게 나타났으며 그 다음으로 신갈나무(11.2%), 떡갈나무(11.0%), 소나무(10.8%) 순으로 각각 나타났다. 교목층 중요치는 굴참나무(20.8%)가 가장 높게 나타났으며 소나무(19.5%), 신갈나무(13.1%), 떡갈나무(11.9%) 순으로 각각 나타났다. 아교목층 중요치는 떡갈나무(15.8%)가 가장 높게 나타났으며 신갈나무(14.2%), 개옻나무(8.4%), 잔털벚나무, 졸참나무(6.9%) 순으로 각각 나타났다. 관목층 중요치는 생강나무(10.6%)가 가장 높게 나타났으며 진달래(9.6%), 졸참나무(6.8%), 개옻나무(5.2%) 순으로 각각 나타났다. 초본층 중요치는 가는잎그늘사초(24.6%)가 가장 높게 나타났으며 큰기름새(11.5%), 주름조개풀, 고사리(2.7%), 조록싸리(2.3%) 순으로 각각 나타났다. 본 식생단위는 교목층과 아교목층에 신갈나무, 굴참나무, 졸참나무, 떡갈나무 등의 참나무류가 높은 중요치로 나타나고 있어 참나무류간 지위경쟁이 심할 것으로 사료되었다.
평균상대우점치는 신갈나무(24.5%)가 가장 높게 나타났으며 그 다음으로 소나무(12.3%), 굴참나무(11.5%), 쇠물푸레나무(5.6%) 순으로 각각 나타났다. 교목층 중요치는 신갈나무(33.9%)가 가장 높게 나타났으며 소나무(23.6%), 굴참나무(20.4%), 졸참나무(6.2%) 순으로 각각 나타났다. 아교목층 중요치는 신갈나무(26.2%)가 가장 높게 나타났으며 쇠물푸레나무(12.9%), 개옻나무(7.7%), 굴참나무(6.1%) 순으로 각각 나타났다. 관목층 중요치는 철쭉(16.0%)이 가장 높게 나타났으며 진달래(11.7%), 생강나무(9.9%), 쇠물푸레나무(9.4%) 순으로 각각 나타났다. 초본층 중요치는 가는잎그늘사초(23.4%)가 가장 높게 나타났으며 큰기름새(6.4%), 둥굴레(3.8%), 쇠물푸레나무(3.1%) 순으로 각각 나타났다. 굴참나무는 신갈나무, 소나무, 상수리나무 등과 혼생하는 종으로(Byeon and Yun, 2016; Lee et al., 2021), 본 식생단위의 교목층에서 높은 중요치를 차지하는 수종과 일치한 것으로 사료되었다. 굴참나무군락에서 신갈나무의 교목층, 아교목층의 중요치가 높게 나타났는데 이는 굴참나무에서 신갈나무로의 천이 단계상(Lee et al., 2002; Kim et al., 2010)에 있는 것으로 사료되었다.
평균상대우점치는 떡갈나무(10.5%)가 가장 높게 나타났으며 그 다음으로 상수리나무(10.3%), 신갈나무(9.8%), 밤나무(8.2%) 순으로 각각 나타났다. 교목층 중요치는 상수리나무(20.1%)가 가장 높게 나타났으며 신갈나무(14.1%), 떡갈나무(13.6%), 일본잎갈나무(10.7%) 순으로 각각 나타났다. 아교목층 중요치는 떡갈나무(12.2%)가 가장 높게 나타났으며 신갈나무(9.4%), 밤나무(9.1%), 졸참나무(8.3%) 순으로 각각 나타났다. 관목층 중요치는 생강나무(10.1%)가 가장 높게 나타났으며 진달래(7.3%), 조록싸리(6.5%), 산초나무(5.1%) 순으로 각각 나타났다. 초본층 중요치는 가는잎그늘사초(12.1%)가 가장 높게 나타났으며 큰기름새(7.2%), 주름조개풀(6.3%), 조록싸리(4.4%) 순으로 각각 나타났다. Choung et al.(2000)은 대구 인접지역의 떡갈나무군락에서 교목층에서는 떡갈나무와 상수리나무, 관목층에서 조록싸리, 산초나무, 초본층에 가는잎그늘사초, 큰기름새가 우점하고 있다고 보고하였으며 이는 본 연구와 유사한 것으로 사료되었다.
평균상대우점치는 신갈나무(16.6%)가 가장 높게 나타났으며 그 다음으로 층층나무(10.3%), 당단풍나무(7.9%), 물푸레나무(7.0%) 순으로 각각 나타났다. 교목층 중요치는 신갈나무(32.6%)가 가장 높게 나타났으며 층층나무(15.9%), 물푸레나무(10.4%), 일본잎갈나무(6.0%) 순으로 각각 나타났다. 아교목층 중요치는 당단풍나무(22.6%)가 가장 높게 나타났으며 층층나무(9.4%), 고로쇠나무(8.8%), 산뽕나무(6.6%) 순으로 각각 나타났다. 관목층 중요치는 노린재나무(9.6%)가 가장 높게 나타났으며 철쭉(8.1%), 미역줄나무(6.9%), 생강나무(6.1%) 순으로 각각 나타났다. 초본층 중요치는 조릿대(20.3%)가 가장 높게 나타났으며 단풍취(4.0%), 가는잎그늘사초(3.2%), 주름조개풀(2.7%) 순으로 각각 나타났다. 본 식생단위에서는 평균상대우점치, 교목층 중요치, 아교목층 중요치에서 신갈나무와 더불어 층층나무가 높은 중요치로 나타났는데, Lee et al.(2014)는 백두대간 북부권역 및 남부권역에서 신갈나무와 함께 층층나무가 우점한다고 하였으며 이는 본 연구와 유사한 결과를 나타내었다.
평균상대우점치는 신갈나무(38.1%)가 가장 높게 나타났으며 그 다음으로 물푸레나무(6.7%), 당단풍나무(5.3%), 물푸레나무(4.6%) 순으로 각각 나타났다. 교목층 중요치는 신갈나무(63.7%)가 가장 높게 나타났으며 물푸레나무(8.3%), 졸참나무(4.1%), 잣나무(3.6%) 순으로 각각 나타났다. 아교목층 중요치는 신갈나무(25.7%)가 가장 높게 나타났으며 당단풍나무(14.9%), 물푸레나무(7.8%), 쇠물푸레나무(7.6%) 순으로 각각 나타났다. 관목층 중요치는 철쭉(18.2%)이 가장 높게 나타났으며 노린재나무(11.2%), 조록싸리(9.7%), 미역줄나무(9.0%) 순으로 각각 나타났다. 초본층 중요치는 조릿대(12.8%)가 가장 높게 나타났으며 가는잎그늘사초(9.6%), 실새풀(9.0%), 대사초(4.1%) 순으로 각각 나타났다. 본 식생단위는 교목층과 아교목층에서 신갈나무의 중요치가 가장 높게 나타나 교란이 없는 한 신갈나무가 우점할 것으로 사료되었고, 아교목층의 당단풍나무, 물푸레나무, 관목층의 철쭉, 초본층의 실새풀, 대사초 등이 출현한 것으로 보아 다른 연구에서 나타난 신갈나무군락의 공통적인 식별종들이 포함된 것으로 나타났다(Chung et al., 2000; Kim et al., 2009, Song et al., 2019b).
백두대간 덕유산권역(늘재 ~ 육십령) 마루금의 산림식생을 식물사회학적 방법으로 분석하여 구분된 9개의 식생단위를 기준으로 종다양도(H'), 최대종다양도(Hmax'), 종균재도(J'), 종우점도(1-J')를 분석한 결과이다. 종다양도는 생물군집 속성을 간접적으로 해석할 수 있으며, 생태적 지위 구성에 있어 종다양성이 높은 군집일수록 보다 복잡하고 안정되며 성숙되었다고 볼 수 있다(Kim et al., 1996). 또한 군집 구조상 복잡성, 외부교란 요인으로부터의 안정성, 그리고 천이 진행과 발달과정상 성숙도는 종다양도와 정비례하는 경향이 있다(Odum, 1969; Loucks, 1970; Bazzaz, 1979). 종균재도는 값이 클수록 안정된 상태에 도달하고 있는 것으로 평가되며(Cho et al., 2012), 종우점도는 0.9 이상일때 1종우점, 0.3~0.7일 때 1종 또는 2종, 0.1~0.3일 때는 다양한 종이 분포하는 것으로 보고되었다(Whittaker, 1965)각 식생단위별 종다양도를 살펴보면 식생단위 2(신갈나무군락군-구상나무군락-가문비나무군)가 2.234로 가장 높은 종다양도로 나타났고 식생단위 1(신갈나무군락군-구상나무군락-퍼진고사리군)이 2.206, 식생단위 4(신갈나무군락군-여뀌군락)가 2.102, 식생단위 3(신갈나무군락군-산오이풀군락)이 2.080, 식생단위 5(신갈나무군락군-굴참나무군락-떡갈나무군)가 1.864, 식생단위 8(신갈나무군락군-층층나무군락)이 1.803, 식생단위 7(신갈나무군락군-떡갈나무군락)이 1.726, 식생단위 6(신갈나무군락군-굴참나무군락-굴참나무전형군)이 1.723, 식생단위 9(신갈나무군락군-신갈나무전형군락)가 1.656의 순으로 각각 나타났다. 각 식생단위의 최대종다양도를 살펴보면 식생단위 4가 3.603으로 가장 높게 나타났고, 식생단위 2(3.314), 식생단위 3(3.238), 식생단위 1(3.140), 식생단위 8(2.992), 식생단위 5(2.856), 식생단위 7(2.840), 식생단위 6(2.723), 식생단위 9(2.660)의 순으로 각각 나타났다. 각 식생단위의 균재도를 살펴보면 식생단위 1이 0.704로 가장 높게 나타났고, 식생단위 2(0.675), 식생단위 5(0.648), 식생단위 3(0.643), 식생단위 6(0.630), 식생단위 9(0.620), 식생단위 7(0.604), 식생단위 8(0.599), 식생단위 4(0.583)의 순으로 각각 나타났다. 각 식생단위의 우점도를 살펴보면 식생단위 4가 0.417로 가장 높게 나타났고 식생단위 8(0.401), 식생단위 7(0.396), 식생단위 9(0.380), 식생단위 6(0.370), 식생단위 3(0.357), 식생단위 5(0.352), 식생단위 2(0.325), 식생단위 1(0.296)의 순으로 각각 나타났다. 식생단위 9의 경우 전체 식생단위 중 가장 낮은 종다양도로 나타났는데 본 식생단위는 신갈나무군락군의 신갈나무전형군락으로 앞선 중요치 결과에서도 신갈나무의 중요치가 가장 높아 안정된 신갈나무군락으로 보았다. 이는 순군락의 종다양도가 혼합된 군락보다 낮다(Kim et al., 2003; Cho and Lee, 2013)는 결과와 일치한 것으로 사료되었다.