



서 론
차나무(Camellia sinensis (L.) Kuntze)는 동백나무과(Theaceae)에 속하는 아열대성 상록활엽수이다. 우리나라는 차나무 자생지의 북방한계선 기후대에 위치해 있다. 최근 집중호우와 폭설, 가뭄, 혹한 등 기상악화 현상이 매년 반복적으로 발생하고 있어, 녹차의 수확 시기가 지연되거나 수확량이 감소하는 등 안정적인 녹차 생산이 어려워지고 있다(Hwang and Kim, 2014).
식물은 다양한 환경 스트레스를 받으며 생장하는데, 온도는 식물의 생활사에 가장 큰 영향을 미치며(Adler and Hille Ris Lambers, 2008), 식물의 분포와 개체 수, 종자휴면, 발아 등을 결정하는 중요한 환경적 요인이다(Woodward and Willians, 1987; Walck et al., 2011). 최근 지구온난화가 빠르게 진행되면서 이상기후가 빈번하게 발생하고 있다. 특히 불규칙한 한파로 인해 이상 저온 피해가 자주 발생하고 있다. 특히, 난대성 상록활엽수는 온난화로 인한 기온 상승의 영향으로 광범위한 지역에 분포지가 확대될 것으로 예측되고 있다(Jin et al., 2019).
저온에 의한 식물의 피해는 증상과 종류가 다양한데, 식물의 저온 피해 증상은 저온장해 유발온도와 지속시간, 식물 생육시기, 식물종 및 재배품종의 유형, 재배지의 지형과 토양특성에 따라 다양하다(Choi and Lee, 1976). 식물이 저온에 노출되면 식물체내 세포의 생리∙생화학적 기능 이상에 따라 형태 및 생리 대사의 변화가 발생한다. 이러한 저온에 대한 다양한 피해 양상은 내한성 평가를 위한 지표로 활용 할 수 있다. 형태적 평가 지표로는 엽록소 함량 분석이 주로 이용되는데, 저온에 처한 식물의 엽록체는 탄소동화작용으로 인한 광저해 현상(photoinhibition)으로 엽록소 및 카로티노이드 함량이 감소하는 것으로 알려져 있다(Wise and Naylor, 1987).
식물의 생리적 변화 지표로는 전해질 용출법이 널리 이용되며, 이는 온도 스트레스로 인해 세포막이 손상되어 용출되는 전해질 용출량을 측정하는 방법이다(Sharom et al., 1994; Maier et al., 1994; Iles and Agnew, 1995; Kim et al., 2010). 전해질 용출량 측정 평가법은 비선형 회귀(nonlinear regression)를 통한 모델로 다양한 식물의 내한성 평가에 적용되고 있다(Ingram, 1985; Anderson et al., 1988; Cardona et al., 1997; Kim, 2006; Kim et al., 2010; McKeller et al., 1992; Jin et al., 2019). 식물의 내한성 평가는 아미노산 및 수용성 당 함량을 측정한다. 저온 조건에서 식물은 탄수화물 분해 촉진으로 수용성 당과 ATP 함량이 증가하는 등 탄수화물 대사가 급격하게 변하는 것으로 알려져 있다(Green and Ratzlaff, 1975; Perras and Sarhan, 1984; Fernandez et al., 2012). 또한 항산화 활성도 및 지질과산화 반응 측정은 저온에 대응하는 식물의 방어 작용 원리를 이용한 내한성 평가 방법이다(Whitaker and Wang, 1987; Parkin and Kou, 1989; Nguyen and Mazliak, 1990; Anderson et al., 1995).
앞서 설명한 전해질 용출량 측정법은 식물의 저온 치사온도를 예측할 수 있다는 장점이 있지만, 식물의 내한성을 과대평가하는 경향이 있다(Jin et al., 2019). 이러한 오류를 최소화하기 위해서는 다양하고 복합적인 인자를 활용하여 저온에 대한 피해를 평가 할 수 있는 복합적인 생리적 기준을 정하는 것이 필요하다. 따라서 본 연구에서는 기후변화에 대비하고자 남부지역에서 주로 재배되고 있는 차나무 4 품종을 대상으로 저온 처리 동안에 나타날 수 있는 다양한 생리적 반응을 복합적으로 비교하여 내한성 품종을 선발하는데 이용하기 위하여 수행하였다.
재료 및 방법
인공 저온처리에 따른 차나무의 생리 반응을 비교 조사하고자 남부지역의 재배품종인 ‘참녹’, ‘보향’, ‘상녹’, ‘명녹’의 삽목묘(3년생)를 전라남도 농업기술원 차산업연구소에서 분양받아 사용하였다. 모든 식물체는 품종별 규격이 일정한 묘목을 선정하여 시판 바이오 상토(신성미네랄)를 담은 플라스틱 포트(W 11 cm × L 11 cm × H 21 cm)에 옮겨 심은 후, 활착을 위해 경상북도 봉화군 소재 국립백두대간수목원 양묘장 온실(37°00′21.1″N 128°47′32.2″E)에서 2020년 5월부터 7월까지 3개월간 순화하였다(Figure 1).

인공 저온처리에 따른 차나무 재배품종의 내한성을 비교하고자 순화 처리한 균일한 식물체를 10본씩 선발하여 실험에 사용하였다. 공시재료를 대상으로 저온처리 전 24시간 동안 상온에서 순화 처리하여 4°C에서 2시간 동안 예비 저온처리 후 실험에 사용하였다. 식물조직의 내한성은 냉각속도에 영향을 받는데, 냉각속도가 시간당 8~15°C를 넘으면 내한성이 과대평가될 수 있기 때문에(Probsting and Sakai, 1979), 저온처리는 저온인큐베이터(SH-75BS, Seyoung Scientific Co., Korea)에서 20분당 1°C씩 낮추었으며, 저온처리 조건은 상온(25°C)을 대조구로 하여, 4°C, −10°C, −15°C에서 1시간 노출 시킨 후 품종별 내한성을 평가하였다(Figure 2).

온도별로 저온 처리한 식물체를 상온(25°C)에서 24시간 동안 순화 처리 후, 저온에 대한 피해 형태를 조사하였다. 저온 피해는 정단 부위 엽 생육상태를 시각적으로 판단하여 5점 시각 척도(1=피해없음, 녹색 잎 100%, 2=10% 잎 피해, 3=10~50% 잎 피해, 4=50~90% 잎 피해, 5=모든 잎 피해, 녹색잎 10% 미만)를 이용하여 5번 반복하여 조사하였다. 생존율은 저온처리 후 약 2개월 뒤 정단부 잎의 녹색 정도, 형태, 피해 정도 등에 전반적 생육상태를 육안 조사하여 생존율을 조사하였다. 생존율은 총 식물체 수에 대한 생존 개체 수의 비를 백분율로 산출하였다(Ryn et al., 2014).
식물에 있어 저온은 세포벽 손상을 유발하며 이는 세포 내의 전해질 유출을 초래하기 때문에 저온처리에 따른 품종별 전해질 용출량 비교 분석하였다. 25°C, 4°C, −10°C, −15°C에서 인공 저온처리한 차나무 품종별로 정단부에서 3~5번째 성엽 표본조직을 원형조직(leaf disc) 형태로 채취하여 실험재료로 사용하였다. 모든 실험재료는 전해질 용출량을 측정하기 위해 저온처리한 잎을 5 g씩 절취하여 증류수 10 ml를 가하고 8시간 동안 진탕한 후 전도도 측정기(VE 4810, Korea Scientifics, South Korea)를 사용하여 저온 처리된 시료 침지액의 전해질 용출량(Lt)을 측정하였다. 저온 처리된 시료는 121°C에서 15분간 처리하여 조직을 괴사시킨 후 24시간 상온에 두었다가 괴사된 시료 침지액의 전해질 용출량(Ltm)을 측정하였다. 각 처리에 대한 측정값은 아래의 식을 이용하여 상대 전기전도도 용출량(relative conductivity, Rt, %)을 백분율로 계산하였다(Dexter et al., 1932).
전해질 용출량과 처리 온도와의 관계를 통해 식물의 저온 치사온도(lethal temperature, LT50) 값을 예측하는 방법은 많은 선행연구에서 사용되고 있다(Jin et al., 2019). 따라서 차나무 품종에 대한 저온 치사온도를 예측하기 위하여 S자 함수의 기본식 2(Yang et al., 2017)에 최소 전해질 용출값(z), 변곡점에서의 기울기(k), 온도(Tm)를 매개변수로 하고, 처리온도를 독립변수로 식 3을 이용하여 계산하였다(Kim et al., 2010).
z = baseline level of electrolyte leackage (%)
Tm = temperature at the inflection point (°C)
k = function of slope at the inflection point
T = treatment temperature (°C)
e = 2.718
스트레스에 반응하는 유리아미노산 중 proline 함량을 저온 스트레스의 생화학적 지표로 사용하고자 하였다. 25°C, 4°C, −10°C, −15°C에서 인공 저온처리한 차나무 품종별로 정단부에서 3~5번째 성엽 조직을 0.05 g 칭량한 다음 5 ml의 3% sulfosalicylic acid 용액과 함께 막자사발로 갈아준 후, 추출액(상등액)을 2 ml 취하여 ninhydrin 용액과 acetic acid 용액을 각각 2 ml씩 첨가 하여 100°C에서 반응시켰다. 이후 4°C의 차가운 물에서 반응을 정지시켜 toluene 용액을 4 ml 첨가하여 교반 후 toluene 용액층만 취하여 흡광도 측정기(Libra S22, Biochrome, England)를 이용하여 520 nm 파장에서 측정하였다.
저온 스트레스는 식물체 내 활성산소 작용을 억제하는 항산화 효소의 활성을 저하시키는 것으로 알려져 있다(Kuk et al., 2001). 이러한 대사적 변화를 관찰하고자 DPPH (2,2-dipenyl-1picrylhydrazyl) 분석법을 통한 항산화 활성도를 지표로 이용하였다(Hannah et al., 2005). 25°C, 4°C, −10°C, −15°C에서 인큐베이션 한 차나무를 품종별로 정단부에서 3~5번째 성엽 조직을 0.1 g으로 칭량한 다음 3 ml의 120 μM DPPH 용액에 20분간 암반응 시켰다. 이후 흡광도 측정기(Libra S22, Biochrome, England)를 이용하여 517 nm 흡광도에서 측정하였다. 항산화 활성 측정값은 아래의 식 4를 이용하여 계산하였다.
식물이 저온 스트레스를 받게 되면 세포 내 활성산소 생성이 증가하여 세포벽 내 지질막의 손상을 초래한다. 저온스트레스에 따른 지질과산화 정도를 malondialdehyde (MDA) 분석법을 이용하여 조사하였다(Heath and Packer, 1968). 25°C, 4°C, −10°C, −15°C에서 인큐베이션 한 차나무 품종별로 정단부에서 3~5번째 성엽 조직을 0.1 g 칭량한 다음 2 ml의 0.1% TCA 용액과 함께 막자사발로 갈아준 후, 추출액(상등액)을 1 ml 취하여 4 ml의 20% TCA 용액을 첨가하여 95°C에서 반응시켰다. 이후 4°C의 차가운 물에서 반응을 정지시킨 후 흡광도 측정기(Libra S22, Biochrome, England)를 이용하여 532 nm와 600 nm 흡광도에서 측정하였다. 지질과산화 반응 측정값은 다음 식 5를 이용하여 계산하였다.
결과 및 고찰
육안조사는 관찰자의 경험적 지식으로 판단하기 때문에 객관적인 판단이라 할 수 없다. 그러나 스트레스에 의한 식물의 외형변화는 내한성의 주요 평가 지표로 사용되어 왔으며, 식물의 외형변화와 전기전도도 사이에는 높은 상관관계가 있다고 보고된 바 있다(Nesbitt et al., 2002).
‘참녹’과 ‘보향’의 경우 25°C, 4°C에서 저온 피해도 지수 ‘1’로 모든 개체가 피해 없이 건전한 모습으로 조사되었다. −10°C에서는 저온 피해도 지수 ‘3’, ‘4’로 잎이 청색으로 변하여 붉은색 옅은 반점이 생기며 동해가 조사되었다. −15°C에서는 저온 피해도 지수 ‘5’로 전체 개체의 잎에 적고 현상이 관찰되어 모든 잎은 동해로 고사하는 것으로 조사되었다[Figure 3(A), 3(B)].
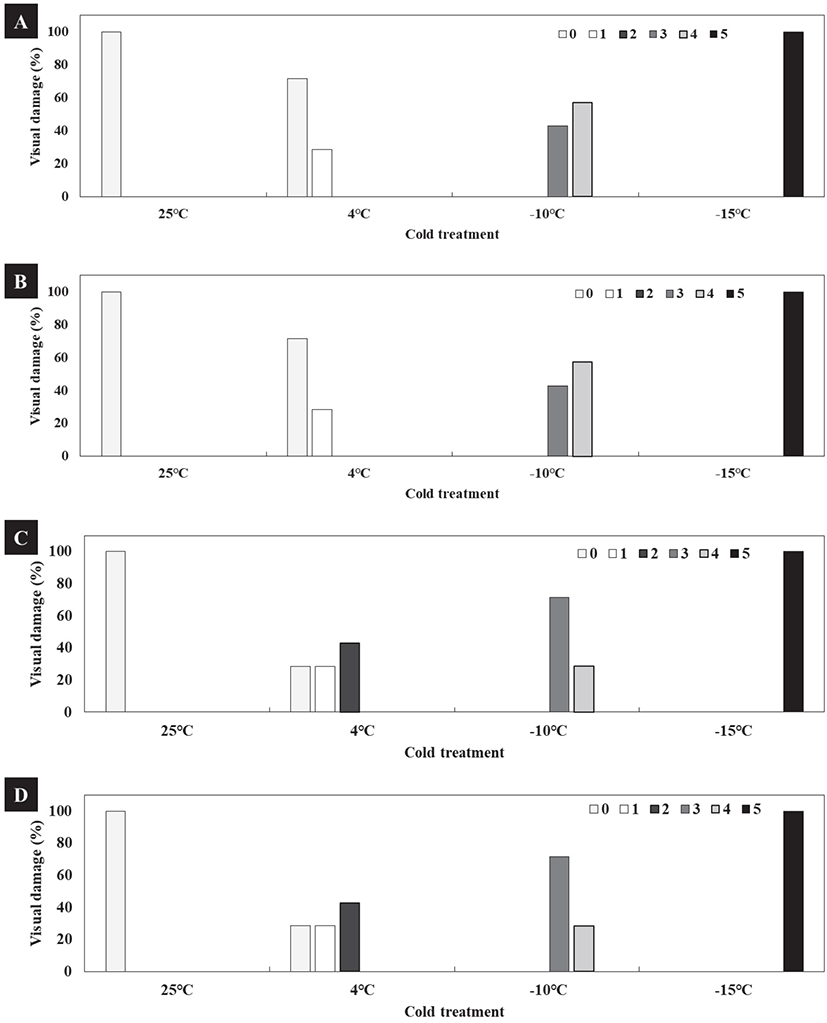
* Visual damage (%) was the value of the number of visual damaged objects compared to the number of total number of cold temperature treatment objects.
‘상녹’과 ‘명녹’의 경우 25°C에서 저온 피해도 지수 ‘1’로 모든 개체가 피해 없이 건전한 모습으로 조사되었고, 4°C에서 저온 피해도 지수 ‘2’로 청색 옅은 반점이 생기며 동해가 조사되었다. −10°C에서 저온 피해도 지수 ‘3’, ‘4’로 잎에 붉은색 짙은 반점이 생기며 동해가 조사되었다. −15°C에서는 저온 피해도 지수 ‘5’로 전체 개체의 잎이 붉게 변하는 적고 현상이 관찰되어 모든 잎은 동해로 고사하는 것으로 조사되었다[Figure 3(C), 3(D)].
이상의 저온 피해도 조사 결과, ‘참녹’과 ‘보향’의 경우 −10°C에서 잎이 청색으로 변하여 동해 피해가 발생하였고[Figure 4(A), 4(B)], ‘상녹’과 ‘명녹’의 경우 −10°C에서 전체 개체의 잎이 붉은색으로 변하여 동해가 조사되었다[Figure 4(C), 4(D)].

차나무의 동해는 청고(靑枯)와 적고(赤枯)로 분류하는데, 청고 현상은 지속적인 저온과 수분 부족으로 인해 차나무 줄기와 뿌리가 냉각되면서 수분 상승이 억제되어 발생한다. 즉 체내 수분부족으로 잎이 건조되어 퇴색된 녹색을 띄다가 고사하는 현상으로(Lee, 2004), Hwang and Kim (2012)은 찻잎의 청고 피해 잎에서 부분적인 세포사멸이 관찰됨을 보고한 바 있다.
적고 현상은 잎의 세포조직이 저온 한계를 넘어 체내에 생긴 얼음 결정체가 세포막을 파괴하여 피해 부위가 적색으로 변하면서 고사하는 현상으로, 찻잎의 적고 피해 잎에서 전체량의 가까운 정도의 세포사멸이 관찰됨을 보고한 바 있다(El-Tayeb, 2006).
본 실험에서는 ‘참녹’과 ‘보향’은 −10°C에서 저온에 의한 청고 피해가 조사되었고, −15°C에서 청고 및 적고 피해가 나타났다. 반면 ‘상녹’과 ‘명녹’은 −10°C와 −15°C에서 적고 피해가 조사되었는데, 이러한 결과는 ‘참녹’과 ‘보향’이 ‘상녹’과 ‘명녹’보다 저온에 대한 내성이 강할 것으로 판단된다.
전해질 용출량 측정법은 온도 스트레스로 인한 식물의 세포막 투과성의 변화로 누출되는 전해질 용출량을 측정하여 세포의 열 안정성(thermo-stability)을 평가하는 방법이다. 이 방법은 단시간에 간편하게 온도 스트레스에 대한 식물 치사 온도를 예측할 수 있어서 식물의 내한성 평가를 위한 지표로써 자주 이용되고 있다(Kim et al., 2014; Jin et al., 2019).
저온처리 후 품종별 잎의 전해질 용출량은 처리 온도가 낮아질수록 증가하였고, 4°C에서 −10°C 처리 구간 사이에 급격하게 증가하였다. 이를 통해 전해질 용출량의 뚜렷한 변화를 보인 4°C에서 −10°C 처리 구간 사이에 차나무 품종별 저온에 대한 내성 한계점이 있을 것으로 추정된다.
4°C 처리구에서 ‘상녹’의 전해질 용출량은 15.83%로 가장 높았으며, 반면 ‘보향’은 8.73%로 전해질 용출량이 가장 낮았다. −10°C 처리구에서 ‘상녹’의 전해질 용출량은 82.07%로 가장 높았으며, ‘참녹’과 ‘보향’은 57.47%, 58.24로 유사한 수준으로 나타났다(Figure 5).
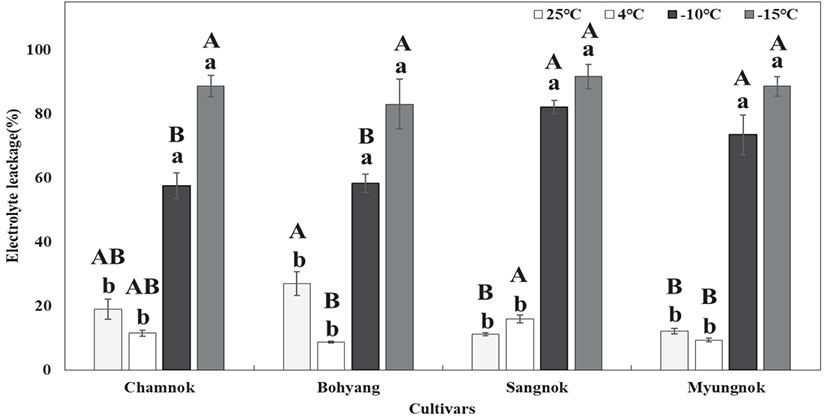
특히 4°C 처리구에서 전해질 용출량이 가장 높았던 ‘상녹’은 −10°C 처리구 에서도 가장 높게 증가하여, 다른 품종들 보다 온도변화에 민감한 수종일 것으로 판단된다. 반면, 4°C 처리구에서 전해질 용출량이 가장 낮았던 ‘보향’은 −10°C 처리구에서도 낮은 수준으로 조사되어, 다른 품종들 보다 저온에 내성이 있을 것으로 판단된다.
국내 상록활엽수종 중 내한성 수종인 동백나무와 다정큼나무는 −15°C 이하에서 저온처리 후 전해질 용출량은 70% 이상으로 나타났고, 내한성이 비교적 낮은 수종인 후박나무와 황칠나무의 경우 80%으로 전해질 용출량이 높게 나타났다고 보고하였다(Jin et al., 2019). 본 실험에서도 차나무의 모든 품종은 −15°C 처리구에서 전해질 용출량이 80% 이상으로 조사되어 일치한 결과를 나타내었다. 특히 −10°C 처리구에서 ‘상녹’, ‘명녹’은 전해질 용출량 70% 이상으로 나타나 저온에 민감한 수종일 것으로 판단되며, ‘참녹’과 ‘보향’은 전해질 용출량이 50% 이상으로 내한성이 비교적 높을 것으로 판단된다.
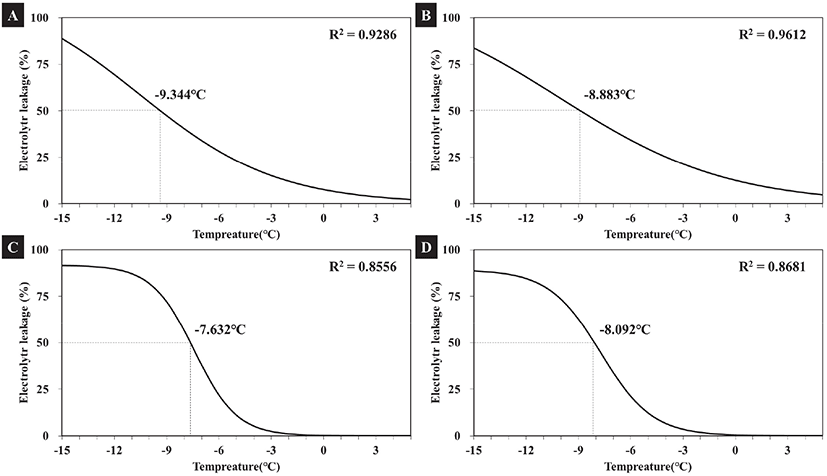
차나무 품종별 전해질 용출량과 처리온도의 관계를 비선형 회귀 분석을 통하여 도출한 결과 기 보고된 연구의 결과와 유사하게 S자 반응 곡선의 형태를 보였다(Anderson et al., 1988; McKellar et al., 1992; Maier et al., 1994; Cardona et al., 1997; Kim et al., 2010). 비선형회귀분석을 통해 저온 처리온도와 전해질 용출 관계를 평가하고자 식 3을 통하여 S자 반응곡선의 변곡점에 해당하는 온도(Tm)를 산출하였고, 이러한 온도는 최대 전해질 용출량의 50% 이상이 용출되기 시작하는 온도로서 내한성 평가에 핵심적인 기준이며, 많은 선행연구에서 예측 치사온도로 평가되고 있다(Anderson et al., 1988; Kim, 2006; Kim et al., 2010). 따라서 변곡점에 해당하는 온도(Tm)가 낮을수록 내한성이 더 높다는 의미를 가진다.
최대 전해질 용출량의 50%가 용출된 온도를 치사온도로 예측해본 결과 차나무 품종별 치사온도 범위는 −7.63°C ~ −9.34°C로 나타났다. 먼저 ‘상녹’의 예측 치사온도는 평균 −7.63°C로 다른 품종들보다 내한성이 가장 약한 것으로 나타났으며, 95% 수준에서 신뢰구간은 −7.90 ~ −7.37°C로 나타났다(Figure 6, Table 1). 즉 차나무 ‘상녹’의 경우 생존 가능한 최저온도가 −7.36°C라고 할 수 있다. 반면 ‘참녹’의 예측치사 온도는 평균 −9.34°C로 95% 수준에서 신뢰구간은 −10.03 ~ −8.65°C로 조사되어 다른 품종들 보다 내한성이 가장 강한 것으로 판단된다. 즉 ‘참녹’의 경우 생존 가능한 최저온도가 −8.65°C라고 할 수 있다. 따라서 본 실험 결과 남부지역에서 주로 재배되는 차나무 4 품종 중에서도 상대적으로 내한성의 차이를 보였으며, 이러한 결과는 동일한 지역 내에서 재배되고 생육하는 수종(Species) 간에도 내한성의 차이가 나타날 수 있음을 알 수 있다(Jin et al., 2019).
유리아미노산의 일종인 proline은 식물이 스트레스 조건에서 반응하여 축적되는 것으로 보고된 바 있다. 특히 스트레스 조건에서 단백질과 세포막의 삼투압 조절과 안정화 역할을 하며(Mohamed, 2000; Farshadfar, 2012; Yang, 2017), 식물의 잎에는 아미노산이 높은 함량으로 축적된다는 보고가 있다(Beemarao et al., 2007).
인공 저온처리 후 proline 함량은 ‘참녹’, ‘보향’, ‘상녹’,‘명녹’ 모든 품종은 처리 온도가 낮아질수록 proline 함량이 증가하였고, 그 중 ‘참녹’과 ‘보향’은 −15°C 처리에서 proline 함량이 21.6 mg/g 및 24.4 mg/g으로, ‘상녹’, ‘명녹’은 19.2 mg/g, 20.4 mg/g으로 조사되어, ‘참녹’과 ‘보향’의 proline 함량이 ‘상녹’, ‘명녹’ 보다 높은 것으로 조사되었다(Figure 7).
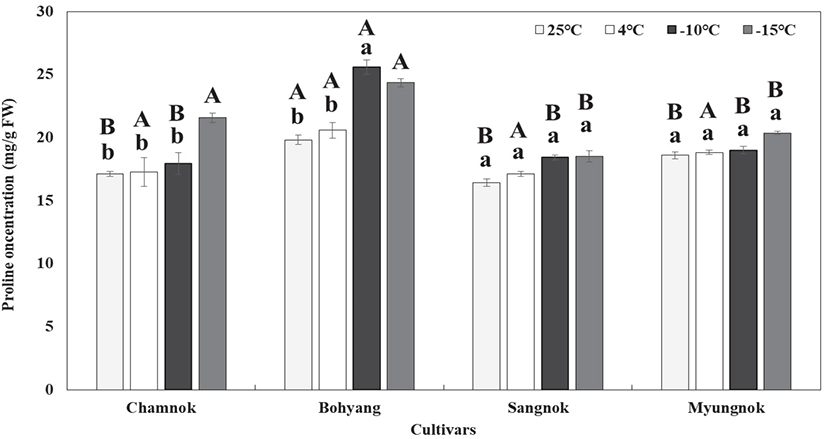
스트레스 처리 후 모든 처리 식물 잎에서는 아미노산 함량이 높아진다고 보고하였으며(Beemarao et al., 2007), 본 실험 결과와 일치하였다. 특히, 25°C처리구에서 ‘보향’은 다른 품종들과 비교했을 때 훨씬 높은 proline 함량을 보유하고 있었으며, 이러한 특징은 4°C에서 −15°C까지 같은 양상을 나타내는 것을 알 수 있었다. 즉 차나무 ‘보향’은 다른 품종들 보다 상대적으로 내한성이 높을 것으로 판단된다.
인공 저온처리에 따른 항산화 활성도를 조사하여 품종별 저온 스트레스의 내성 정도를 평가하고자 하였다. 식물이 저온과 같은 환경 스트레스를 받게 되면 세포 내 활성산소종이 과다하게 생성된다(Anderson et al., 1995). 활성산소종은 각종 스트레스 반응에 대한 매개체로서 스트레스에 대한 내성을 증진하는 역할을 하는데(Salin, 1991), 이는 식물의 활성산소종 제거를 위한 항산화 대사기능을 활발하게 하여 식물이 저온장해에 대한 저항성을 갖도록 유도한다(Lee et al., 2014).
인공 저온 처리 후 항산화 활성도 측정 결과, ‘참녹’과 ‘보향’은 온도가 낮아질수록 항산화 활성이 높게 나타났고, ‘상녹’과 ‘명녹’은 온도가 낮아질수록 항산화 활성도가 비슷하거나 낮은 값으로 조사되었다(Figure 8).
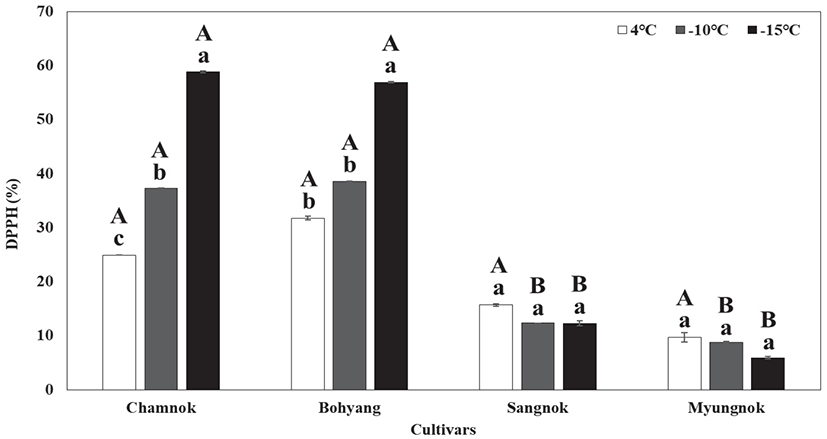
특히, ‘참녹’과 ‘보향’의 항산화 활성은 4°C에서 24.9%, 31.7%로 조사되었고, −15°C에서 58.8%, 56.9%로 조사되어, 4°C의 항산화 활성도와 비교했을 때 항산화 활성 수치가 2배 증가하는 것을 알 수 있었다. 반면, ‘상녹’과 ‘명녹’은 항산화 활성은 4°C에서 15.7%, 9.7%로 조사되었고, −15°C에서 12.3%, 5.9%로 조사되어, 4°C의 항산화 활성도와 비교했을 때 항산화 활성이 오히려 낮아지는 것을 알 수 있었다.
Kim et al.(2009)은 스트레스 내성 품종 선발 시 전해질 용출과 항산화 활성도는 높은 상관관계를 나타낸다고 하였는데, 본 실험에서도 저온 처리 시 항산화 활성도가 낮게 나타난 ‘상녹’과 ‘명녹’은 민감종으로 판단되고, 항산화 활성도가 높게 나타난 ‘참녹’과 ‘보향’은 다른 품종에 비해 내한성이 높을 것으로 판단된다.
저온 스트레스에 의해 식물 체내 생성되는 활성산소종은 세포막 지질의 불포화 지방산과 결합하여 산화량의 증가를 유발하며, 이로 인하여 세포막 지질의 구조가 변화된다(Patterson et al., 1976; Bailly et al., 1996). 또한, malondialdehyde (MDA), jasmonic acid 등과 같은 과산화작용의 부산물이 세포질 내에 축적되어 이러한 현상은 식물의 저온 저항성에 대한 지표로 이용되고 있다(Lofkowska and Holubowska, 1989; Shen et al., 1999; Lee et al., 2014).
인공 저온 처리 후 지질과산화 반응 측정 결과 ‘참녹’, ‘보향’, ‘상녹’, ‘명녹’은 온도가 낮아질수록 지질과산화 반응에 의한 MDA 축적 함량이 증가하였다(Figure 9). 특히, 저온에 내성을 보인 ‘참녹’과 ‘보향’은 −10°C에서 MDA 축적 함량이 각각 0.9 μM, 0.8 μM로 나타나, 저온에 민감성을 보인 ‘상녹’(1.3 μM)과‘명녹’(1.6 μM)을 비교했을 때 MDA 축적 함량이 훨씬 적은 것으로 조사되었다.
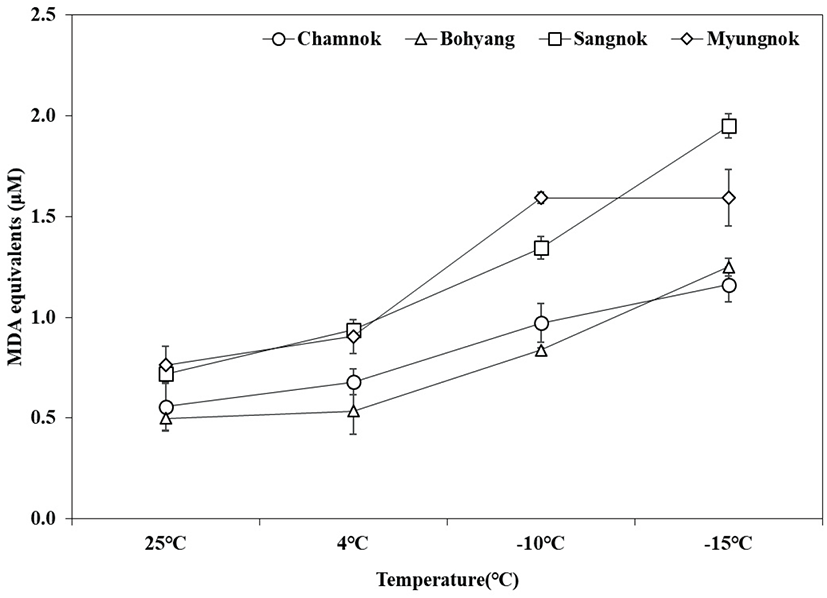
Kuk et al.(2001)은 저온 처리 후 지질과산화 작용의 지표인 MDA 생성량은 내성 및 민감성 품종 간 유의적인 차이를 보이며 민감성 품종의 MDA 생성량이 크게 증가 한다고 보고하였는데, 본 실험에서도 온도가 낮아질수록 저온처리에 민감성을 보인 ‘상녹’과 ‘명녹’은 내성을 보인 ‘참녹’, ‘보향’에 비해 MDA 축적 함량이 높은 값으로 조사되어 유사한 결과를 나타내었다.
결 론
본 연구는 기후변화에 대응하여 차나무 내한성 품종을 선발하고자 남부지역 주 재배 품종인 ‘참녹’, ‘보향’, ‘상녹’, ‘명녹’을 대상으로 인공 저온처리에 따른 내한성을 비교 평가하였다. 인공 저온처리에 따른 4 수종 차나무 품종의 전해질 용출량을 측정한 결과 처리 온도가 낮아질수록 그 값이 증가하였다. 전해질 용출량과 저온 처리 온도의 관계를 비선형회귀 분석을 통해 치사온도를 예측한 결과,‘참녹’은 −9.34°C, ‘보향’ −8.88°C로 예측되어 저온에 대한 저항성이 ‘명녹’(−8.09°C), ‘상녹’(−7.63°C) 보다 상대적으로 높게 나타났다. 이러한 결과는 저온피해도, 항산화 활성도, 지질과산화 반응과도 일관성 있게 나타나 차나무 품종 ‘참녹’과 ‘보향’은 ‘상녹’과 ‘명녹’에 비해 상대적으로 내한성이 강한 것으로 판단된다. 이러한 지표들은 기후변화에 대응한 차나무의 내한성 평가와 선발에 중요한 생리적 지표로 이용할 수 있을 것으로 판단된다. 또한 본 연구 결과는 내한성 차나무 외 다른 내한성 식물의 육성에도 활용될 수 있을 것으로 판단된다.