



서 론
희귀식물로 평가되어진 식물의 보전은 전 세계적인 수준에서뿐만 아니라 국가적인 수준에서 종다양성의 유지를 위해 절대적으로 중요하다(Pimm et al., 1988; Djebbouri and Terras, 2019). 희귀식물(Rare plants)은 전 세계적으로 분포하고 있는 개체수가 20,000개체 이하로서 개체군의 크기가 매우 작고, 분포가 넓다고 하더라도 생육 장소가 극히 제한되어 있으며(Chung, 1991), 생태적 지위에 따라 구성종의 특징적인 분포 유형이나 범위를 가진다(Grime, 1977; Korea Forest Research Institute, 1996; Dupré and Ehrlén, 2002; Grime, 2002; Pearman et al., 2008).
망개나무(Berchemia berchemiaefolia)는 갈매나무과(Rhamnaceae), 망개나무속(Berchemia)에 속하는 낙엽성 교목으로 한국, 일본, 중국에 자생하며 충청북도와 경상북도의 극히 일부 지역에 제한된 규모로 분포하는 희귀식물이다(Lee, 1979; Korea Forest Service and Korea National Arboretum, 2008; Korea National Arboretum, 2021). 1996년 산림청 지정 희귀식물 취약종(Vulnerable, VU)으로 지정되었으며(Korea Forest Research Institute, 1996), 2005년 야생동식물보호법 시행령에 의해 멸종위기동식물 II급 식물로 지정되었으나, 신규 자생지 및 추가 개체군이 발견되어 2012년 멸종위기식물 목록에서 제외되었다(Ministry of Environment and National Institute of Biological Resources, 2012).
우리나라에선 1935년 속리산 법주사 계곡에서 처음 발견된 이후, 충북 보은의 속리산, 괴산의 덕가산 그리고 제천의 월악산에 자생하는 망개나무 개체군은 천연기념물로 지정되어 지속적인 관리와 연구가 수행되고 있으나, 경상북도의 경우 포항과 청송 소재 망개나무 자생지(Kim, 2010)와 군위군 소재 망개나무림(Lee et al., 2005)의 군집구조와 개체군 구조의 연구만이 수행되었을 뿐 이들 자생지와 환경요인간의 생태적 특성을 분석한 연구는 없다.
망개나무는 내음성(shade tolerance)이 약한 양수이며, 유식물과 치수는 자생지 내 낙엽이 쌓여 있지 않아 토양이 그대로 드러나 있는 곳에 그리고 숲틈(forest gap)이 있어 햇빛이 잘 들어 올 수 있는 환경조건에서 발생한다(Lee et al., 2005). 그러나 주로 계곡부의 전석지와 바위틈에서 자라고 있어 천연갱신이 어렵다(Song et al., 2008).
망개나무의 재질은 결이 곧고 단단하며, 불에 잘 타기에 과거 농촌에서는 연료목으로 사용되거나, 농기구를 만들어 활용되었다. 이로 인해 우량한 망개나무는 벌채되고 잔류목에 세대 갱신이 이루어지면서 자생지 내 근친교배가 군집의 활력을 저하시키거나 자손 세대의 유전 다양성을 감소시키고 있다(Lee et al., 2003).
본 연구는 경북 포항 내연산 소재의 망개나무 임분을 대상으로 개체군 구조와 군집구조를 정량적으로 분석하고, 군집유형과 이에 영향을 미치는 주요 입지환경요인을 분석하여 그 생태적 특성을 규명하고자 하였다. 이를 통해 내연산 망개나무군락의 체계적 보전 및 관리 방안 수립을 위한 기초자료를 제공하고자 한다.
재료 및 방법
내연산은 행정구역상 포항시 북구 죽장면, 송라면, 청하면에 걸쳐 분포하며, 지리적으로는 북위 36°02′∼36°50′, 동경 128°59′∼129°34′에 걸쳐 분포하고 있다(Gyeongsangbuk-do Arboretum, 2007). 주봉은 향로봉(932 m)이며, 보현산(1,123 m), 천령산(775 m) 및 수석봉(821 m) 등의 평균 고도가 500 m 이상인 비교적 높은 산들이 둘러싸고 있어 지형 및 지세가 매우 복잡하고 수계가 잘 발달되어 있다.
조사가 수행된 망개나무 임분들은 지형적으로 해발고도 240∼450 m에 분포하였으며, 경사도는 11∼33°이었다. 토양은 암쇄토가 대부분을 차지하였으며, 토양산도는 pH 4.9∼5.7의 범위에 있어 전체적으로 약산성이었다.
조사지의 기후정보는 Worldclim-Global Climate Data (www.worldclim.org)에서 제공하는 50년 평균 기후자료(1950∼2000년)를 추출하여 제시하였으며, 19개의 BIOCLIM 자료 중 PCA(Principal Components Analysis) 분석을 통하여 나온 대표 기후정보인 연평균기온은 3.2±0.6°C, 최온난분기의 평균기온은 24.1±0.5°C, 최한랭분기의 평균기온은 2.3±0.7°C 그리고 연평균 강수량은 1,422±55.6 mm로 나타났다(Table 1).
내연산 일대의 망개나무 개체군 구조를 파악하기 위하여 2003년 6월∼11월(5개월간)까지 총 7회에 걸쳐 조사된 망개나무 전수조사 기초자료를 수집하여 개체군의 수평적, 수직적 구조를 분석하였다. 본 조사 자료는 노거수회에서 수행하였으며, 경상북도수목원에서 시작하여 연산폭포를 지나 보경사 입구까지 계곡부를 따라 내려오면서 출현하는 망개나무 모든 개체에 대하여 흉고직경(cm), 수고(m) 그리고 맹아지수의 수를 조사하여 기록한 자료이다.
내연산 망개나무림의 현장조사는 2022년 5월과 8월의 총 2회 실시하였다. 대상임분은 비교적 층위구조가 안정되어 있으며, 망개나무 개체수가 가장 많은 6지점을 선정하여 조사하였다(Figure 1). 조사구의 크기는 교목층에 우점하는 수목의 수고를 참고하여 임분의 종 구성을 잘 대변할 수 있는 최소면적으로 10×10 m 5개소와 20×20 m 1개소로 적절하게 설정하였다.
식생조사는 현장에서 수관의 높이에 따라 교목층(tree layer, ≥8 m), 아교목층(subtree layer, 3∼8 m), 관목층(shrub layer, 1∼3 m) 그리고 초본층(herb layer, ≤1 m)으로 구분하였으며, 출현수종을 대상으로 Braun-Blanquet(1964)의 통합우점도를 적용하였다. 또한 각각의 조사구에서 좌표(GPS), 해발고도(m), 사면방위(degree), 경사도(°) 그리고 수관층위별 식피율(%)의 환경요인을 수집하였다.
각 층위별 출현한 수종의 상대적 우점도를 비교하기 위해 수종별 양적(피도), 질적(빈도)자료를 정량화하여 중요치(Importance Value, IV)를 산출하였으며(Bray and Curtis, 1957), 이를 통합해 백분율로 환산한 상대중요치(Brower and Zar, 1977)를 층위별로 비교하였다. 또한, 층위별 수목의 현존량이 다르기에 가중치를 적용한 평균상대우점치(Mean Importance Percentage, MIP)를 산출하고자(교목층 IV×3+아교목층 IV×2+관목층 IV×1)/6을 적용하였다. 자료의 정리는 엑셀 프로그램(Microsoft Office 2013)을 통해서 수행하였으며, 이를 통해 식생자료 main matrix를 생성하였다.
식물동정은 Lee(2003)을 통해 실시하였으며, 수종의 판단이 어려울 경우 사진, 수피, 잎 등의 정보를 획득하여 실내에서 동정하였다. 학명, 과명 그리고 종명표기는 국립수목원이 제시한 국가표준식물목록을 참고하였다(Korea National Arboretum, 2022).
망개나무 임분의 토양 특성을 파악하기 위해 각 조사구마다 3지점에서 표토의 유기물 층을 제거한 후 토심 0∼15 cm 깊이의 토양을 채취하였다. 일부 토심이 얕은 곳에서는 채취 가능한 깊이까지의 시료를 채취하였다.
채취한 토양 샘플은 지퍼백에 봉인하여 실험실로 운반하여 풍건시켰으며, 풍건된 토양은 2 mm 토양체(US standard No.10)를 이용하여 2 mm 이상의 석력을 걸러내어 분석을 수행하였다. 토성을 포함한 화학적 성분분석은 산림토양분석 전문기관인 한국농업기술진흥원(Korea Agricultrue Technology Promotion Agency)에 의뢰하여 분석을 수행하였으며, 분석항목은 토양의 화학적 특성인 pH, 유기물함량, 전질소(N), 유효인산(P), 양이온치환용량, 치환성양이온(K, Ca, Mg)이었다.
정리된 자료를 토대로 군집분석의 한 종류인 TWINSPAN (Two-Way INdicator SPecies ANaysis)을 실시하였다. TWINSPAN 분석은 모든 수종의 자료를 기초로 구분하는 분류과정으로(Hill, 1980), 이를 통해 조사가 수행되어진 망개나무 임분의 군집유형을 구분하였으며, 유형별 군집의 층위구조, 구성수종 그리고 입지환경을 간접적으로 파악하였다.
또한 망개나무 임분의 종구성을 비롯해 입지환경 요소의 유사성에 근거하여 상대적인 분포패턴과 주요 수종과 하층식생과의 연관성을 알아보고자 Ordination 분석인 DCA(detrended correspondence analysis)와 CCA를 실시하였다(Hill, 1980). 이를 통해 군집유형에 영향을 미치는 환경요인을 보다 정량적이고 객관적으로 분석하고자 하였다. 이러한 다변량 통계분석은 PC-ORD6 소프트웨어를 통하여 분석하였다.
결과 및 고찰
내연산 망개나무림에 대한 개체군의 직경급분포도, 수고급분포도 그리고 분지수분포도를 작성하였으며, 본 자생지와 행정구역이 동일하고 조사 시기가 비슷한 Lee et al.(2005)의 경상북도 군위군 장곡리 망개나무의 개체군 자료와 비교하였다(Figure 2).
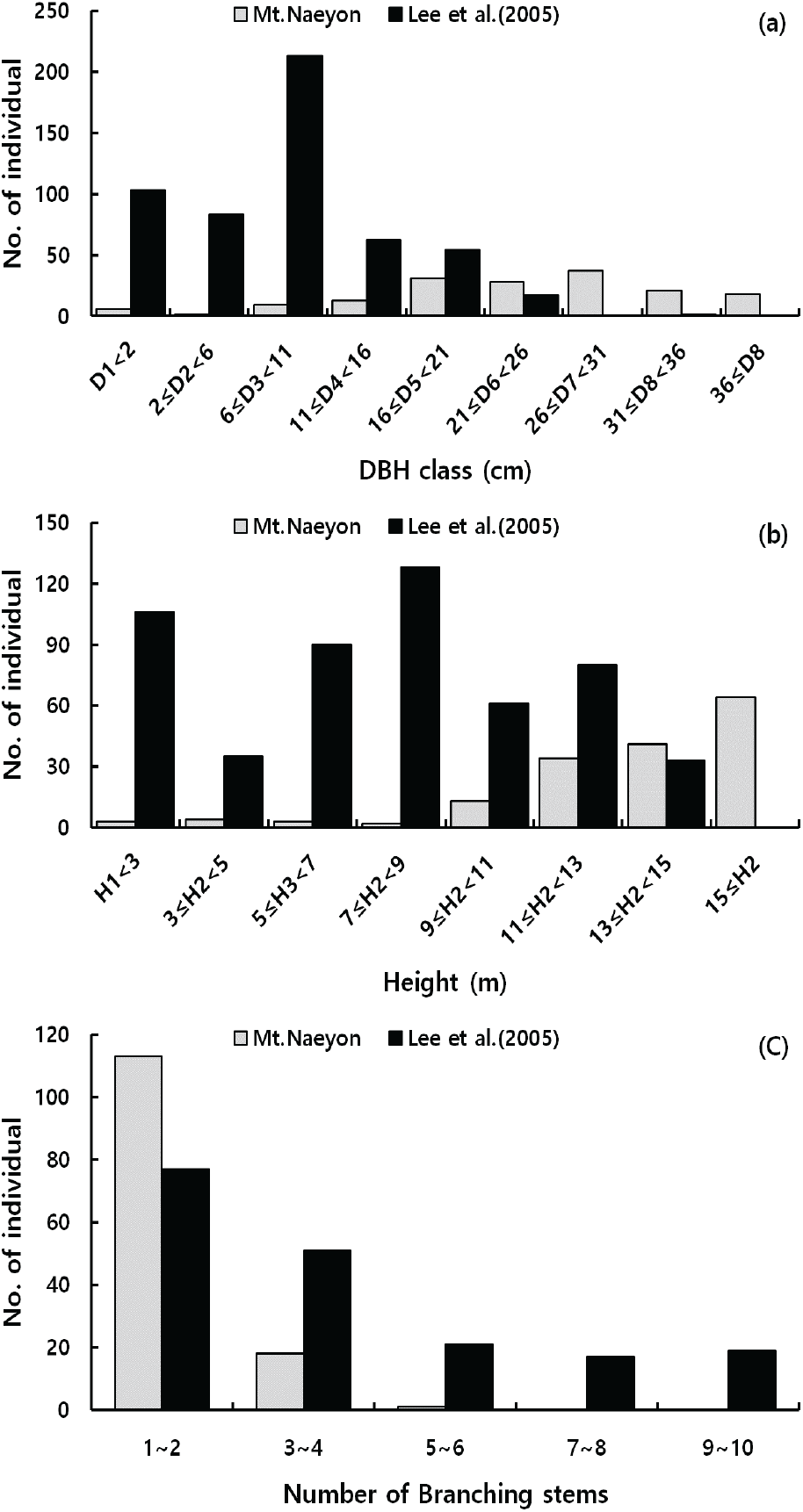
내연산 일대의 망개나무는 총 164개체가 출현하였고, 평균 흉고직경은 24.5 cm이었으며, 흉고직경 26∼30 cm 사이에 가장 많은 개체가 분포하였다. 흉고직경급별 망개나무의 출현빈도는 정규분포형의 밀도를 나타내고 있는 것으로 보아 앞으로도 당분간은 망개나무림으로 유지될 것으로 보이나, 흉고직경 5 cm 이하 유식물(seedling), 치수(sapling) 그리고 어린 나무(juvenile)와 수고 8m 이하의 하층식생(understory)의 밀도가 매우 적기에 망개나무 차대림의 발달이 어려울 것으로 판단된다.
그러나 Lee et al.(2005)의 조사된 군위군의 개체군 구조는 내연산과 정반대로 나타났는데, 흉고직경 6∼10 cm 사이에 가장 많은 개체가 출현하는 역J자형의 양상으로 후계목의 보충이 계속될 것이며, 수고 8 m 이하의 하층식생의 밀도가 상층식생보다 높아 망개나무의 우세가 계속될 것으로 보고하였다.
이들 망개나무림의 가장 큰 차이점은 수관의 상층부가 열려있어 빛에너지가 들어 올 수 있는 숲틈(forest gap)의 존재와 임상에 낙엽이 쌓여 있지 않아 토양이 그대로 드러나 있는 토양층의 발달이었다. 실제 개엽기(5월 초)에 망개나무 유식물과 치수의 출현이 잦았으나 울폐된 수관층에 의한 광량의 차단과 빈약한 토양층으로 인한 뿌리의 활착 지연은 망개나무림의 재생을 저해하는 요인으로 보고되었다(Kim et al., 2012).
또한 출현한 모든 망개나무 개체의 맹아로부터 발생된 분지수를 측정한 결과, 분지수는 최고 5개까지 나타났으며, 대부분 2개 이하로 나타났다. 망개나무 자생지로 알려진 경북 군위(Lee et al., 2005), 충북 괴산의 군자산(Lee et al., 2012)과 속리산(Kim et al., 2012)의 경우에도 망개나무는 대부분 맹아에 의한 갱신이 이루어지고 있었는데, 이는 과거로부터 망개나무 자생지가 교란이나 인간의 간섭이 지속되어 왔으며, 그 재생방법으로 맹아가 발생되었음을 대변하는 것이다(Kang et al., 1982).
내연산 망개나무림의 층위에 따른 구성종의 우점도를 알아보고자 각 층위별 구성종의 출현종수와 중요치(IV)를 산출하여 비교하였다(Table 2).
망개나무 임분 6개소에서 출현한 식물은 총 39분류군이었으며, 평균 18.5종이 출현하였다. 층위별 종수는 교목층이 5.2±1.2종이었으며, 아교목층은 2.3±1.2종이, 관목층은 6.0±3.2종이 그리고 초본층은 9.8±1.8종이 출현하였다.
교목층(Tree layer)에서 망개나무의 중요치(IV)는 62.3%로 상층수관의 대부분을 형성하는 우점수종이었으나 아교목층에서는 2.1%로 그 출현빈도가 매우 낮았으며, 관목층과 초본층에서는 출현하지 않았다. 그러나 천이후기단계 종으로 보고되는 졸참나무(5.4%), 쪽동백나무(5.1%), 서어나무(5.0%)가 교목층에서 비교적 높은 중요치로 출현하였으며, 아교목층에서 쪽동백나무(35.6%), 까치박달(16.7%), 서어나무(3.1%)가, 그리고 관목층과 초본층에서도 이들 수종들의 유식물과 치수의 출현이 나타나고 있었다. 이는 양수인 망개나무가 향후 숲틈과 같은 교란체제(disturbance regime)가 없는 평형상태에 도달하였다면 음수인 서어나무나 까치박달로 대치가 됨으로써 중요치가 감소될 것이다.
총 6개소의 망개나무 임분에서 출현한 39분류군의 정량자료를 활용하여 TWINSPAN 분석을 수행한 결과, 제 1수준(Level 1)에서 고유치(eigenvalue) 0.374의 정보량으로 지표종인 단풍취(+)에 의해 그룹Ⅰ과 그룹II의 총 2개 그룹으로 분류되었다(Table 3).
식물종의 출현빈도 차이가 그룹간의 2배 이상일 때 지표종이 되며(Sasaki et al., 2015), 이는 그룹별 식물군집의 환경기울기를 간접적으로 보여주는 것이다(Lee et al., 1994). 지표종인 단풍취는 차광정도와 습도가 높을수록 출현빈도가 높고 생육이 더 양호하였기에(Ahn et al., 2018) 그룹Ⅰ과 그룹II의 군집구분은 수관층의 식피율과 이에 파생된 토양수분 구배가 가장 중요한 환경요인으로 작용하게 된 것으로 판단된다.
TWINSPAN 분석을 통해 구분된 그룹별 구성종과 층위별 종조성을 분석한 결과는 다음과 같다. 본 조사는 망개나무 개체수가 가장 많은 임분에서 수행되어졌기에 모든 그룹(Ⅰ, II)의 교목층에서 망개나무는 가장 높은 중요치를 보여주며 최우점하고 있었다. 또한 각 군집명은 교목층의 망개나무와 두 번째로 높은 중요도를 보인 차우점종을 바탕으로 명명하였으며, 망개나무-졸참나무군집(그룹 Ⅰ)과 망개나무-서어나무군집(그룹 II)으로 명명하였다.
망개나무-졸참나무군집(그룹 Ⅰ)의 교목층은 망개나무(59.3%), 졸참나무(10.9%), 물푸레나무(6.8%) 등의 양수성 수종들이 흉고직경 26 cm 이상의 대경목으로 분포하였으며, 망개나무(최대직경 38 cm)와 졸참나무(최대직경 80 cm) 등의 우세목이 임분 내 모자이크 형상으로 분포하고 있었다.
양수들이 교목층에 우점하고 있는 사실은 망개나무-졸참나무군집의 입지 조건이 과거로부터 수목 성장을 위한 충분한 광량이 제공되었음을 보여주는 것이다. 실제 본 군집에서 출현한 망개나무 65개체 중에서 34개체(약 55%)가 맹아로부터 발생(최대 분지수 4개)되었는데, 이 맹아 개체들은 과거 교란이나 인간의 간섭에 따른 매우 열악한 조건하에 있었음을 보여주는 결과로 사료되며, 이러한 교란의 결과 발생된 숲틈(forest gap)은 수평 및 수직적으로도 빈 공간을 형성하여 양수들의 성장과 생육에 충분한 광량을 제공했을 것으로 판단된다.
또한 숲틈으로 들어오는 직사광과 산란광은 토양층의 수분구배에 영향을 주어 하층식생에서 우점하고 있는 주요 수종의 분포패턴에 영향을 줄 것이다. Park and Jang (1998)은 당단풍나무와 생강나무 우점군집이 약간 건조한 토양수분 상태였으며, 까치박달 우점군집은 주로 산지 하부 및 계곡 평탄부에 위치하여 비교적 습윤한 토양수분 상태를 유지한다고 보고하였다.
이처럼 망개나무-졸참나무군집(그룹Ⅰ)은 입지조건과 자생지의 교란에 의한 발달단계, 종조성이 다른 군반(patch)이 모여 모자이크 형상을 이루고 있었으며(Whittaker, 1977), 시 · 공간적 다양성을 나타내고 있었다(Sousa, 1984).
망개나무-서어나무군집(그룹 II)은 주로 활엽수림의 하층 구성종으로 내음성이 강한 단풍취를 식별종으로 그룹화 되었는데, 이는 앞서 수관층의 틈(gap)이 군집형성에 중요한 요인이었던 그룹Ⅰ과는 반대로 수관층의 높은 식피율과 이로 인해 임분내 광조건이 불량하여 비교적 습윤한 토양 상태을 대변하는 것이다.
이는 망개나무-서어나무군집(그룹 II)의 층위별 종조성에서도 확인할 수 있었다. 교목층에서 양수인 망개나무의 중요치가 65.3%로 상층 수관의 대부분을 형성하는 우점종이었으나 아교목층에서는 4.2%로 그 출현 빈도가 매우 낮았으며, 관목층과 초본층에서 치수나 유식물의 출현은 없었다. 그러나 음수인 서어나무는 교목층과 아교목층에서 각각 9.0%, 6.3%의 비교적 높은 중요치를 보였으며, 하층식생인 관목층과 초본층에서 치수와 유식물이 출현하였는데, 만약 망개나무-서어나무군집이 평형상태에 도달하였다면 망개나무는 천이후기종으로 보고되는 서어나무로 대치되어 그 중요치는 계속 감소 될 것으로 사료된다.
DCA 분석을 실시한 결과, 고유치(eigenvalue)는 제 1축이 0.4660, 제 2축이 0.0963으로 총 6개 임분은 그 정보량에 따라 각각의 좌표축 상에 배열되었다(Figure 3, Table 4).
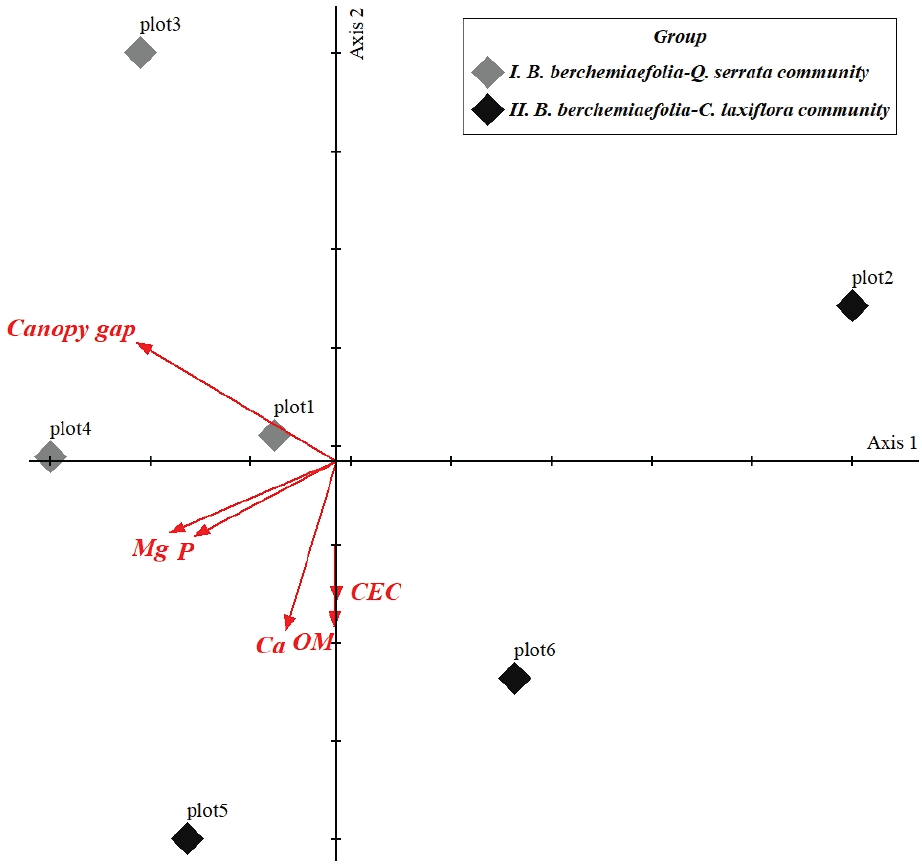
이는 TWINSPAN에 의한 군집의 분류(그룹Ⅰ과 II) 경향과 대체로 유사하였는데, 제 1축을 기준으로 왼편에는 그룹Ⅰ이, 제 2축을 기준으로 아래쪽에는 그룹II의 임분들이 위치하고 있었다. 이렇게 인접해 분포하는 임분들은 종조성 및 임분구조의 유사성을 보여주는 것으로, 결국 입지환경을 반영하는 것이다.
각 그룹에서 멀리 떨어져 위치한 plot2는 임분간 거리가 매우 인접하였던 그룹Ⅰ의 임분들(plot1, plot3, plot4)과 종조성이 다르게 나타났는데, 이는 그룹Ⅰ의 임분들이 주로 산지 하부에 집중되어 위치하였고, 과거 교란의 영향으로 맹아 개체들의 출현율(약 55%)이 높았으며, 수관틈이 18.3%이었던 것에 반해, 비교적 평탄부에 위치한 plot2의 식피율은 95%로 망개나무(최대직경 44 cm)가 수관층의 대부분 형성하였으며, 아교목층에선 천이후기종인 서어나무의 출현빈도가 높은 비교적 온전한 상태였기 때문으로 판단된다.
또한 DCA 분석을 통해 수관틈(canopy gap)과 치환성양이온인 Ca 및 Mg, 유기물 함량(OM), 유효인산(P) 그리고 양이온치환능(CEC)이 임분들의 분포에 영향을 미치는 주요 환경요인으로 확인되었다. 이는 경상북도 망개나무 개체군이 수관개방율과 토양의 유기물 함량, 치환성양이온 Ca와 밀접한 관계를 지닌다는 보고와 같았다(Kim et al., 2011). 그러나 이러한 환경요인들이 임분들을 군집화하는데 그 역할이 미비하였는데, 이는 상관 상으로 균질해 보이는 군집일지라도 각 군집의 입지 조건이 다양한 환경요인과의 상관관계에 의해 이질적인 공간분포를 나타내고 있기 때문이다(Lee and Jo, 2000).
그룹Ⅰ의 임분들은 수관틈(Canopy gap)과 강한 양(+)의 상관관계를, 그룹II는 치환성양이온 Ca, 유기물함량(OM) 그리고 양이온치환능(CEC)과 양(+)의 상관관계를 보여주었지만 군집화되지는 않았다. 즉, 서로 상반된 환경요인의 영향하에 구성종의 분포가 크게 두 개의 패턴으로 나뉘었으며, 대표적인 교목성 수종은 앞서 TWINSPAN 분석 시 각 그룹별 망개나무 다음으로 중요치가 높았었던 그룹Ⅰ의 망개나무, 졸참나무와 그룹II의 망개나무와 서어나무이었다.
토양층에서 그룹II의 치환성 Ca, 유기물함량(OM) 그리고 양이온치환능(CEC)은 그룹Ⅰ보다 다소 높게 나타났다. 수목 성장에 중요한 영향을 미치는 유기물이 그룹II에서 비교적 높았던 이유는 망개나무의 주요 자생지가 산지 계곡부의 경사지와 평탄부가 만나는 습성 입지조건으로 상부 낙엽층이 쉽게 분해되어 표토층에 유입되었기 때문으로 판단된다. 비록 토양유기물 함량에 직접적으로 영향을 미치는 낙엽층에 대한 조사는 이루어지지 않았지만, 그룹간의 우점 수목의 본수와 흉고직경은 큰 차이를 보이지 않았기에 토양유기물 함량의 차이는 주요 자생지의 입지 조건 차이 때문으로 해석된다(Park and Jang, 1998).
치환성 Ca와 양이온치환능(CEC)이 높게 나타난 것 역시 입지 조건으로 해석될 수 있는데, 그룹II는 높은 유기물함량과 습윤 조건으로 상부 낙엽층이 쉽게 분해되어 물과 양분이 잘 흡수될 수 있는 반면, 그룹Ⅰ의 임분들은 산지 하부에 분포하였으며, 비교적 넓은 수관틈으로 들어오는 빛과 바람의 영향으로 인해 건조한 미기후가 조성되어 낙엽층 분해가 잘되지 않아 표토층으로 물과 양료의 유입이 적었기 때문으로 사료된다.
CCA 분석을 통해 망개나무림을 구성하는 주요 교목성 수종인 망개나무, 졸참나무 그리고 서어나무와 하층식생 구성종 간의 연관성을 알아보고자 CCA 분석을 수행하였으며, 상층과 하층의 구성종들 사이에는 강한 연관성이 있었는데, 이는 앞서 설명한 종조성이 다른 군반이 모여 모자이크 형상을 이루고 있는 망개나무 자생지의 공간적 이질성을 보여준다(Figure 4).
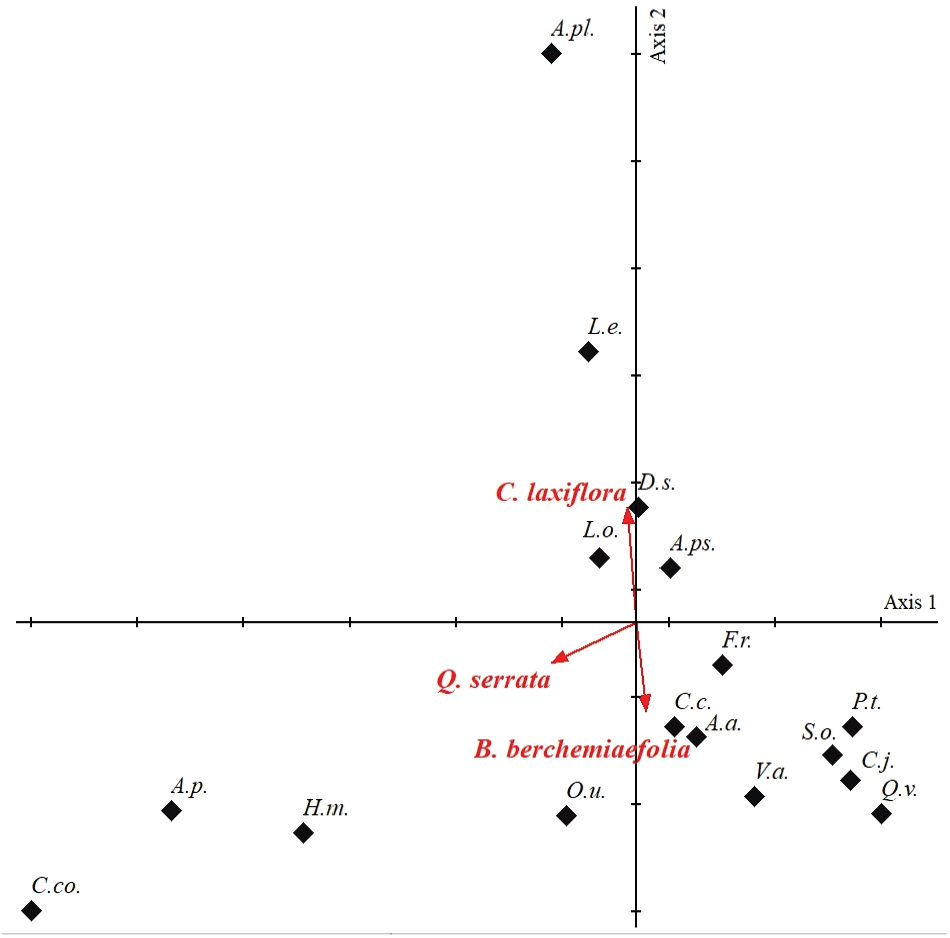
중심에서 기원하는 주요 교목성 수종을 바탕으로 망개나무(B. berchemiaefolia)는 주름조개풀(O.u.), 남산제비꽃(V.a.), 쪽동백나무(S.o.), 굴참나무(Q.v.), 작살나무(C.j.) 그리고 담쟁이덩굴(P.t.)과, 졸참나무(Q. serrata)는 까치박달(C.co.), 고로쇠나무(A.p.), 산수국(H.m.)과 그리고 서어나무(C. laxiflora)는 박쥐나무(A.pl.) 및 비목나무(L.e.)와 밀접한 연관성을 나타내었다.
결 론
경상북도 포항시 내연산 소재의 망개나무 임분을 대상으로 개체군 구조 및 층위별 종조성을 정량화하고, 다변량통계기법을 활용하여 군집유형과 입지환경요인과의 상관관계를 분석하였다.
내연산 망개나무는 총 164개체가 출현하였으며, 흉고직경은 1.1∼76.4 cm의 범위로 평균 24.5㎝인 정규분포형의 밀도분포를 보였다. 수고기준 관목층(≤3 m) 이하에서 망개나무는 총 5개체(약 3%)만 출현했기에 향후 차대림의 발달은 어려울 것이며, 출현한 망개나무의 37.1%는 2개 이상의 맹아개체로 과거부터 현재에 이르기까지 망개나무 임분들이 매우 열악한 조건하에 생육하고 있음을 대변하는 것이다.
망개나무 임분들은 망개나무-졸참나무군집(그룹Ⅰ)과 망개나무-서어나무군집(그룹II)의 2개의 그룹으로 분류되었다. 또한 수관틈(Canopy gap)의 크기수준, 토양의 화학적 특성인 유기물함량(OM), 치환성 Ca 그리고 양이온치환능(CEC)이 망개나무 임분들의 분포에 영향을 주는 주요 입지요인이었다. 이중 수관틈의 여부와 이에 파생된 토양의 수분상태는 내음성이 약한 양수인 망개나무, 졸참나무 그리고 음수인 서어나무의 종조성과 유지 여부에 가장 중요한 입지 조건으로 판단되었다.
결과론적으로, 내연산 망개나무군집이 평형상태에 도달하였다면 망개나무는 음수이면서 아교목층 이하에서 비교적 높은 우점도를 보이며 출현하고 있는 서어나무나 까치박달로의 천이가 진행될 것으로 사료되지만, 숲틈과 같은 임분 내의 교란체제가 변하지 않는다면 망개나무, 졸참나무와 같은 양수들은 분포지의 입지 조건에 따라 계속하여 우점할 것이다.