Article
강원도 가리왕산 지역 천연 활엽수림에서 물푸레나무 (Fraxinus rhynchophylla)의 천연하종과 치수 발생 및 고사 특성
장유림1,
정종빈2,
김현정3,
김종우1,
강규석1,
남광현1,
박필선1,*
Seed Rain, Seedling Emergence and Mortality of Fraxinus rhynchophylla in Natural Broad-leaved Forests in the Mt. Gariwang Area, Gangwon-do
You Lim Jang1,
Jong Bin Jung2,
Hyun Jung Kim3,
Jongwoo Kim1,
Kyu-Suk Kang1,
Kwang Hyun Nam1,
Pil Sun Park1,*
Author Information & Copyright ▼
1Department of Agriculture, Forestry and Bioresources, Seoul National University, Seoul 08826, Korea
2Division of Forest Ecology, National Institute of Forest Science, Seoul 02455, Korea
3Forest Resource Department, Baekdudaegan National Arboretum, Bonghwa 36209, Korea
© Copyright 2023 Korean Society of Forest Science. This is an Open-Access article distributed under the terms of the
Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits
unrestricted non-commercial use, distribution, and reproduction in any
medium, provided the original work is properly cited.
Received: Apr 06, 2023; Revised: Jun 15, 2023; Accepted: Jun 16, 2023
Published Online: Sep 30, 2023
요 약
물푸레나무(Fraxinus rhynchophylla Hance)의 천연하종 갱신과 치수의 초기 생존 특성을 파악하기 위하여 강원도 가리왕산 지역 천연 활엽수림에서 물푸레나무를 대상으로 2011-2013년 3년 간 하종량, 2012-2014년의 치수 발생, 치수의 수명 분포(Lifespan distribution), 치수 생존과 고사, 초본 식생과의 관계를 분석하였다. 하종량과 치수 발생량은 연도별 유의한 차이를 보였고(p <.05). 하종량이 증가한 이듬해는 치수 발생량이 증가하였다. 2013년 하종량과 2014년 치수 발생량은 유의한 상관 관계를 보였고(p <.05), 이는 하종 이듬해 발아하는 물푸레나무의 치수 발생 특성과 일치한다. 치수는 늦봄에서 초여름 사이에 발생하였으며, 2014년은 발생 치수 중 78%가 초여름에 한꺼번에 발생하였다. 물푸레나무 치수는 발생 직후 대부분이 고사하였다. 2012년 8월 발생한 치수 집단은 초기 두 달 동안 56%가 고사하였고, 2014년 7월 발생한 치수 집단은 1개월 동안 38%가 고사하였다. 하층의 초본 식생으로 인한 피음과 지하부 경쟁은 치수 고사의 주요 원인이었다. 천연 활엽수림의 물푸레나무는 특정 해에 종자가 많이 열리는 해거리 경향을 보였고, 다량 결실 이듬해는 임상에 다량의 치수가 발생하였으나, 발생 당년 대부분이 고사하였다. 물푸레나무는 치수 고사율이 높으나, 주기적으로 다량의 치수가 발생하고 살아남은 소수의 치수가 활착에 성공하여 갱신하는 전략을 가진다. 우리나라에는 다양한 자생 교목 수종이 서식하나 이들 수종의 천연갱신에 대한 정보는 제한적이다. 우리나라의 천연림 관리 및 복원을 위해서는 다양한 자생 수종의 천연갱신에 대한 연구가 필요하다.
Abstract
Diverse native tree species are distributed in Korean forests; however, information on their regeneration strategies is limited. This study analyzed seed rain from 2011 to 2013, seedling emergence, seedling survival, lifespan distribution, and the relationship between seedling mortality and herbaceous vegetation for 3 years, from 2012 to 2014, to understand the natural regeneration and early survival characteristics of Fraxinus rhynchophylla Hance in natural broad-leaved forests. Seed rain and emerged seedling density significantly differed among the years (p < .05). An increase in seed rain was followed by an increase in seedling emergence in the following year. Seed rain in 2013 and seedling emergence in 2014 showed a significant correlation (p < .05), confirming the trait of F. rhynchophylla seeds that germinate in the following year after seed production. Seedlings emerged in late spring and early summer. Of the seedlings, 78% emerged in the early summer of 2014. Most seedlings died immediately after emergence. Of the seedlings that emerged in August 2012, 56% died within 2 months, and 38% of those that emerged in July 2014 died within 1 month. Shade and competition against ground vegetation chiefly affected the seedling survival of F. rhynchophylla. Fraxinus rhynchophylla showed mast seeding, with a large number of seed production in a particular year. A surge of seedling emergence followed in the early summer of the next year. However, most seedlings died in the same year of emergence. Fraxinus rhynchophylla has a high seedling mortality, but mast seeding and the resulting high seedling emergence produce a few survived seedlings, enabling the continuation of the F. rhynchophylla population. Efforts and research on the natural regeneration of native tree species are required to effectively manage natural forests in Korea.
Keywords: mast seeding; natural regeneration; survival curve; lifespan distribution; seedling mortality; herbaceous vegetation
서 론
우리나라는 700종 이상의 목본 수종이 자생하고 있으며, 한반도 중부지역은 기후적으로 낙엽활엽수가 우점하는 지역에 속한다(Korea National Arboretum, 2022). 우리나라의 낙엽활엽수림은 대부분 천연림으로, 참나무속(Quercus)을 중심으로 단풍나무속(Acer), 물푸레나무속(Fraxinus), 자작나무속(Betula), 느릅나무속(Ulmus), 서어나무속(Carpinus), 벚나무속(Prunus) 등의 다양한 수종이 임분 상층을 구성하고 있다(National Geographic Information Institute, 2016). 최근 생물다양성 보전과 기후변화 대응 측면에서 자생 활엽수를 이용한 산림 조성에 대한 관심이 높아지고 있다. 낙엽활엽수림은 우리나라 천연림의 반 이상을 차지하고 있으나, 자생 활엽교목의 갱신에 대한 정보는 제한적이다. 이들 수종은 천연림에서 천연하종 또는 맹아를 이용하여 갱신한다(Kim et al., 2001).
천연하종이란 주변의 나무로부터 종자가 자연적으로 산포하는 것을 말하며, 천연하종된 종자가 발아하여 차대의 임목으로 자라난다(Lee et al., 2010). 천연림에서 천연하종 갱신은 숲의 천이와 임분 발달에 이바지한다. 울진 금강송림이나 봉화의 소나무림에서는 주변 임분에서 유래한 신갈나무(Quercus mongolica Fisch. ex Ledeb.), 물푸레나무(F. rhynchophylla Hance) 등의 낙엽 활엽수종이 하층을 점유하며 향후 상층의 주요 수종으로 자라날 것으로 예측하였다(Park et al., 2010; Kwon et al., 2018). 천연하종으로 발생하는 하층의 임목 군집은 임분의 미래 모습을 예측할 수 있게 하고 천연림 관리에 중요한 기초자료로 활용된다.
해거리(Mast seeding)란 임목이 주기적으로 특정 해에 다량 결실을 맺는 현상이다(Koenig, 2021). 임목에서 자연적으로 떨어진 종자로부터 치수가 발생하여 갱신이 일어나는 천연하종갱신은 임목의 해거리로 인한 종자생산량의 변동에 크게 영향을 받는다(Lee et al., 2010). 치수의 연도별 발생 변동은 종자 생산의 변동과 일정 기간의 간격을 두고 일치하는 경향을 보이며(Tapper, 1992), 해거리를 하는 수종은 치수 발생량의 연도별 변동성 또한 뚜렷하다(Ralph and Brinkman, 1996). 자연 상태에서 물푸레나무는 3~4년에 한 번씩 해거리를 하는 것으로 알려져 있다(Sung, 1996; Lee et al., 2010; Sung et al., 2011; Korea National Arboretum, 2016).
활착 초기의 치수는 여러 환경 변화에 취약하여 고사하기 쉽다(Kitajima and Fenner, 2000; Moles and Westoby, 2004; Manuel and Molles, 2015). 주변 식생과의 광 경쟁 또는 지하부 수분과 양분 경쟁이 흔히 치수의 고사를 야기한다. 낮은 광 환경에서 F. pennsylvanica Marshall와 Tilia cordata Mill. 치수의 고사율이 증가하는 경향을 보였고(Küßner, 2003), F. americana L.와 Acer rubrum L. 치수는 토양 조건이 양호하더라도 관목 아래 그늘에서는 성장이 저하되었다(Putz and Canham, 1992). Fraxinus excelsior 치수는 Dactylis glomerata L.와 지하부 바이오매스에서 경쟁적인 상호관계가 나타났다(Bloor et al., 2008).
임목 개체군의 크기는 새로운 세대의 갱신과 기존 임목의 고사에 의해 결정되며, 고사하는 개체가 갱신되는 개체보다 많으면 임목 개체군은 쇠퇴한다(Kimmins, 2004). 토양에 떨어진 종자는 발아에 적합한 온도와 수분 조건 등의 환경이 충족되면 발아하여 임상의 치수 집단을 구성한다. 임상에 공급된 종자로부터 발아한 치수는 개체군의 새로운 세대를 구성하는 주요 집단이다. 연약한 치수가 발아 직후 다량 고사하면 집단의 크기가 급감하는 생존곡선(Survival curve)이 나타난다(De Steven, 1991; Tanouchi et al., 1994; Ralph and Brinkman, 1996). 치수 집단을 이루는 각각의 개체는 발생 이후 환경 및 수종의 내재적인 특성에 따라 생존 기간, 즉 수명(Lifespan)이 다르다. 이를 바탕으로 개체의 생존 기간을 종합한 치수 집단의 수명 분포(Lifespan distribution)를 파악할 수 있다(Ralph and Brinkman, 1996).
물푸레나무는 전국에 분포하는 낙엽활엽교목으로, 신갈나무, 소나무(Pinus densiflora Siebold & Zucc.), 굴참나무(Q. variabilis Blume) 등과 더불어 오대산, 계방산 등 백두대간의 사면, 산복, 능선 지역에 분포한다(Kim et al., 2003; Lee et al., 2010; Kim et al., 2014). 물푸레나무의 꽃은 잡성화로 같은 나무에 양성화, 단성화가 섞여 있고 4~5월 중에 개화한다. 날개가 달린 시과가 9월에 익으며 바람에 의해 산포하여 천연하종에 의한 발아가 용이하다(Korea National Arboretum, 2016). 강원도 활엽수림에서 물푸레나무의 종자는 모수로부터 30 m의 거리까지 비산되고, 휴면 후 2~3개월 간 습윤조건에 노출되면 6월 말에 치수가 다량 발생한다(Sung, 1996; Sung et al., 2011).
물푸레나무는 우리나라 산림생태계의 주요 수종이다(Kim et al., 1999; Lee et al., 2010). 하지만, 천연활엽수림에 분포하는 물푸레나무의 해거리 및 천연하종갱신과 이로 인해 발생하는 치수 집단군의 발생 및 생존 양상에 대한 정보는 제한적이다. 본 연구는 강원도 가리왕산 지역 천연활엽수림에서 천연갱신하는 물푸레나무의 천연하종, 치수 발생 및 생존을 3년 동안 모니터링하여 종자 생산의 해거리 경향과 치수 발생을 파악하고, 천연갱신된 치수 집단의 단기간 수명 분포, 생존그래프 및 고사율과 초본 식생과의 상관관계를 분석하였다. 천연활엽수림에서 물푸레나무의 갱신 양상 및 치수 집단의 동태를 파악하여 천연활엽수림의 효과적인 관리방안을 위한 기초 자료를 제공하고자 하였다.
재료 및 방법
1. 연구 대상지 및 조사구 설치
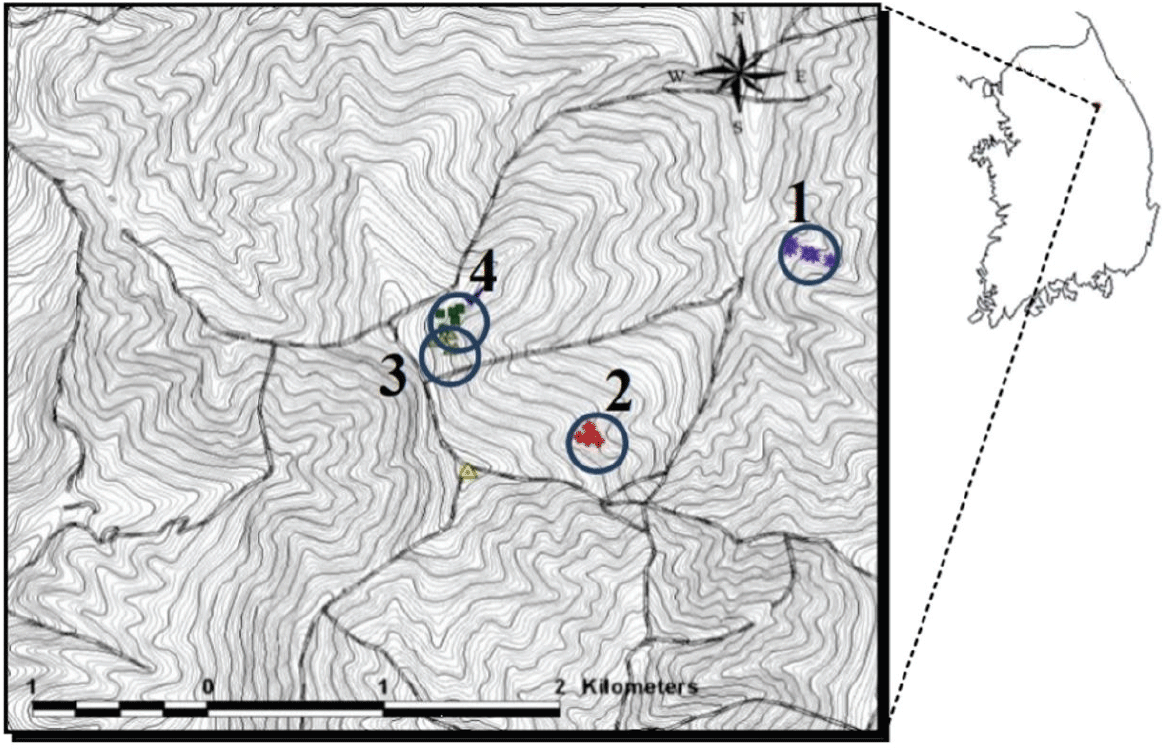
강원도 평창군 가리왕산 및 중왕산에 위치한 동부지방산림청 평창국유림관리소 관내 국유림경영단지의 천연활엽수림(북위 37°27′~37°28′, 동경 128°31′~128°32′)에 4곳의 조사지를 설정하였다(Figure 1). 조사지는 해발고도 965~1,210 m에 위치하며, 경사는 30~69%로 급했다. 연구지에 설치한 호보간이기상장치(HOBO, Onset Computer Corporation, USA)를 이용하여 측정한 2012~2014년 연평균 기온은 5.7°C, 연평균 지중(지하 5 cm) 온도는 8.2°C였다. 연중 일평균은 최소 −21.4°C(2012년 2월)에서 최대 23.4°C(2012년 8월)까지 변화를 보였다. 물푸레나무 치수가 발생하기 시작하는 5월 월평균 기온은 약 13°C, 지중온도는 약 11°C로 나타났다. 연평균 강수량은 1,353 mm이며, 7~8월에 전체 강수량의 48~61%가 집중되었다(Gangwon Provincial Office, 2014; Korea Meteorological Administration, 2015).
Figure 1.
Four study sites in natural broad-leaved forests on Mount Gariwang (Site 1) and Mount Joongwang (Sites 2, 3 and 4), Gwangwon-do.
Download Original Figure
연구지역은 평안누층군의 동고층이 분포하고, 대부분 사암으로 이루어져 있다(Korea Institute of Geoscience and Mineral Resources, 2022). 토양은 산성갈색삼림토이며, 미국 농무성의 토양분류에 의하면 청심통(CsF)에 속하고, 토성은 미사질양토이다(National Institute of Agricultural Sciences, 1975). 평균 유효토심은 50-100 cm, 낙엽층의 깊이는 6-10 cm였고, 임상은 전석지와 조릿대 밀생지역, 일반 토양이 혼재한다(Korea Forest Service, 1999).
조사지역에는 물푸레나무를 비롯한 신갈나무, 고로쇠나무(A. pictum subsp. mono (Maxim.) Ohashi), 당단풍나무(A. pseudosieboldianum (Pax) Kom.), 층층나무(Cornus controversa Hemsl.), 까치박달(C. cordata Blume), 들메나무(F. mandshurica Rupr.), 피나무(T. amurensis Rupr.) 등 40여종의 수종이 분포하고 있었다. 신갈나무는 조사지의 최우점수종으로 임분 밀도의 6.4~17.5%를 차지하고 있었다. 고로쇠나무(6.0%~10.7%), 물푸레나무(3.0%~8.0%), 느릅나무(U. davidiana var. japonica (Rehder) Nakai; 1.6~ 5.1%) 순으로 임분에서 차지하는 비율이 높았다(Eastern Regional Forest Service, 2009). 임분 내 물푸레나무의 평균 흉고직경은 7 cm 전후였고, 흉고직경 범위는 약 40 cm까지 보고되었다(Korea Forest Service, 1999). 초본층에는 얼레지(Erythronium japonicum (Balrer) Decne.), 바람꽃류(Anemone spp.), 노루오줌(Astilbe rubra Hook.f. et Thomson), 숲개별꽃(Pseudostellaria setulosa Ohwi), 참나물(Pimpinella brachycarpa (Kom.) Nakai), 현호색(Corydalis remota Fisch. ex Maxim.), 벌깨덩굴(Meehania urticifolia (Miq.) Makino) 등이 분포하고 있었다(Kim et al., 2015).
2. 하종량 조사
물푸레나무의 하종량을 조사하기 위해 조사지 당 1×1 m의 종자망을 9개씩, 총 36개의 종자망을 설치하였다. 종자망은 플라스틱 섬유재질의 방충망(1 mm mesh)으로 제작하였고, 지면으로부터 1 m 높이에 설치하여 낙하물을 수집하였다. 2011년, 2012년, 2013년 각 연도의 8월 초부터 11월 초까지 한 달 간격으로 종자망을 수거한 뒤, 실험실에서 종자를 분리하여 수종별 종자 개수를 파악하였다. 수집된 종자 중 물푸레나무 종자는 크기(3~4 cm) 및 형태(좁고 긴 날개를 가진 시과)를 토대로 동정하였다.
3. 물푸레나무 치수와 초본 조사
물푸레나무 치수 집단을 모니터링하기 위해 각 조사지에 1×1 m의 소방형구를 15개씩, 총 60개를 설치하였다. 2012년부터 2014년까지 각 년도의 5월 첫째 주(2012년 제외), 6월 첫째 주(2014년도 7월 첫째 주), 8월 첫째 주, 10월 첫째 주에 소방형구의 치수(근원경 3 cm 이하)와 초본 식생을 조사하였다. 소방형구 내에서 종자로 발아한 치수를 대상으로 개체마다 각각 번호를 부여하고, 깃발표식을 꽃아 표시하여 치수 개체별 생존 및 고사를 지속적으로 모니터링하였다(Figure 2). 조사 시기마다 각 개체 별 수종, 발생 후 최초 관찰시점, 생존 및 고사 여부를 조사하였다. 이와 더불어 같은 소방형구에서 관찰되는 초본 식생의 초고를 측정했다.
Figure 2.
Photo of seedling monitoring at 1×1 m subplot. Each seedling was monitored using numbered tags.
Download Original Figure
4. 자료 분석
단위면적당 물푸레나무 종자의 하종량과 치수 발생량을 계산하고, 연도별 하종량과 치수 발생량을 비모수 분산분석(Kruskal-Wallis test)을 이용하여 비교하였다. 사후 검정은 Bonferroni correction을 이용하였다. 하종량과 이듬해 치수 발생량 간에 비모수 상관 분석(Spearman’s rank correlation)을 수행하였다.
치수 발생량이 매우 적은 2013년을 제외한 2012년, 2014년 발생 치수 집단에 대해서는 고사율과 생존 기간을 분석하였다. 2012년 8월과 2014년 7월 발생이 최초 관찰된 집단을 각각의 발생 집단으로 보고, 이를 대상으로 조사 기간 동안의 생존여부를 바탕으로 생존곡선을 도출하였다. 2012년에 발생한 치수 전체와 2014년에 발생한 치수 전체에 대해 각 치수 개체가 고사한 시기를 확인하고 이를 통해 개체별 수명을 계산, 2014년을 시점으로 한 치수 집단의 수명 분포를 분석하였다(Ralph and Brinkman, 1996).
2014년도 7월 초에 발생한 치수 집단에 대해 8월 초, 10월 초에 치수 개체의 고사 여부를 조사하였다. 이를 바탕으로 7~8월 사이의 고사율과 8~10월 사이의 고사율을 계산하였고, 초본 식생 높이와의 상관분석을 수행하였다. 2014년 7~8월 고사율은 2014년 7월 초본 식생 초고 및 8월 초본 식생 초고와 상관관계를 분석하였다. 2014년 8~10월 고사율은 2014년 8월 초본 식생 초고 및 10월 초본 식생 초고와 상관관계를 분석하였다. 상관분석은 비모수 상관분석(Spearman’s rank correlation)을 이용하였다. 통계 분석은 R 프로그램(version 4.0.3)을 이용하였다.
결 과
1. 연도별 하종량과 치수 발생
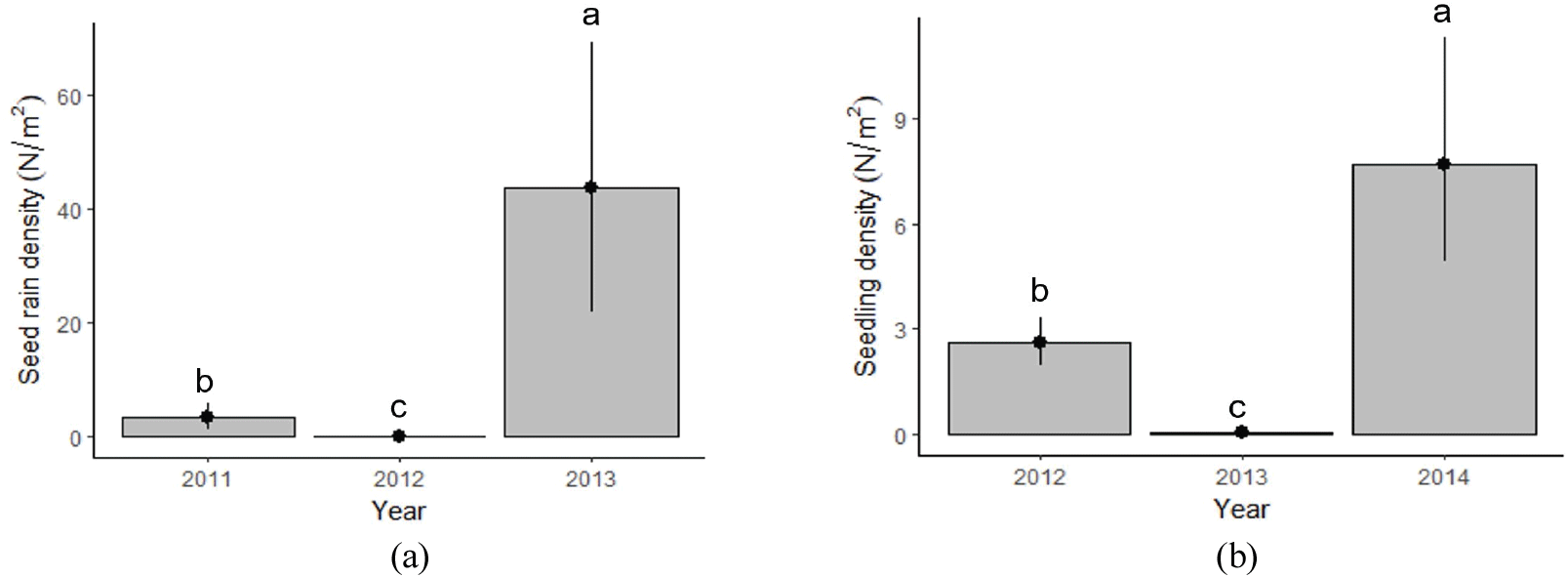
물푸레나무의 하종량과 치수 발생량은 3년 동안 연도별로 유의한 차이를 보였다[p < .05; Figure 3(a)]. 단위면적당 하종량은 2011년 3.5립/m2, 2012년 0.2립/m2, 2013년 43.9립/m2으로 연도별 유의한 변동을 보였다(p<.05). 특히 2013년도에는 다량의 종자가 임상에 떨어졌는데 이는 이듬해 2014년 다량의 치수 발생으로 이어졌다.
Figure 3.
Mean density of (a) seed rain (number of seeds/m2) from 2011 to 2013 and (b) seedling emergence (number of seedlings/m2) from 2012 to 2014 of Fraxinus rhynchophylla in the Mt. Gariwang area, Gangwon-do. Mean±SE. Different letters indicate means are significantlydifferent at p<.05.
Download Original Figure
치수 발생량은 2012년 2.6본/m2, 2013년 0.1본/m2, 2014년 7.7본/m2으로 하종량과 같이 연도별로 유의한 증감을 보였다[p<.05; Figure 3(b)]. 하종량 대비 이듬해 치수 발생량 비율은 2011년 76.2%, 2012년 47.1%, 2013년 17.5%로, 2011년에는 하종된 종자량에 비해 이듬해 가장 높은 비율의 치수가 발생하였다. 2013년에는 다량의 종자가 생산되었지만 이듬해 치수 발생 비율은 2011년과 2012년보다 낮았다.
2013년 하종량과 2014년 치수 발생량은 유의한 양의 상관관계를 보였다(p<.05; Table 1). 하종량이 거의 없었던 2012년의 이듬해인 2013년 치수 발생이 거의 없었던 반면, 하종량이 많았던 2013년의 이듬해인 2014년도에는 치수가 다량 발생하였다(Figure 3).
Table 1.
Spearman’s rank correlation coefficient between seed rain density and seedling density of Fraxinus rhynchophylla from 2011 to 2014.
|
|
Seed rain density |
|
|
2011 |
2012 |
2013 |
| Seedling density |
2012 |
0.36 |
|
|
| 2013 |
−0.11 |
−0.33 |
|
| 2014 |
0.22 |
0.30 |
0.80* |
Download Excel Table
2. 치수 발생 시기
2014년도에 발생한 대부분의 치수가 7월과 8월 초 조사에서 최초로 관찰되었다(Table 2). 특히 7월 초 78%의 치수가 발견되었고, 8월 초 조사에서 새로 관찰된 치수는 2014년도 발생 치수의 20%였다. 5월 초와 10월 초에 새로 나타난 치수는 거의 없었다. 이는 전년도에 하종된 종자가 겨울과 봄까지 휴면을 유지하다 늦봄(5~6월)과 여름철(7월)에 한꺼번에 다량으로 발아했음을 나타낸다.
Table 2.
The density of newly emerged Fraxinus rhynchophylla seedlings that were observed in the beginning of May, July, August and October in 2014.
| Observation date |
Density of newly emerged seedlings (N/m2) |
Ratio (%) |
| 3 May 2014 |
0.07 |
1 |
| 2 July 2014 |
5.97 |
78 |
| 1 August 2014 |
1.55 |
20 |
| 2 October 2014 |
0.10 |
1 |
| Total |
7.68 |
100 |
Download Excel Table
3. 치수의 생존 기간
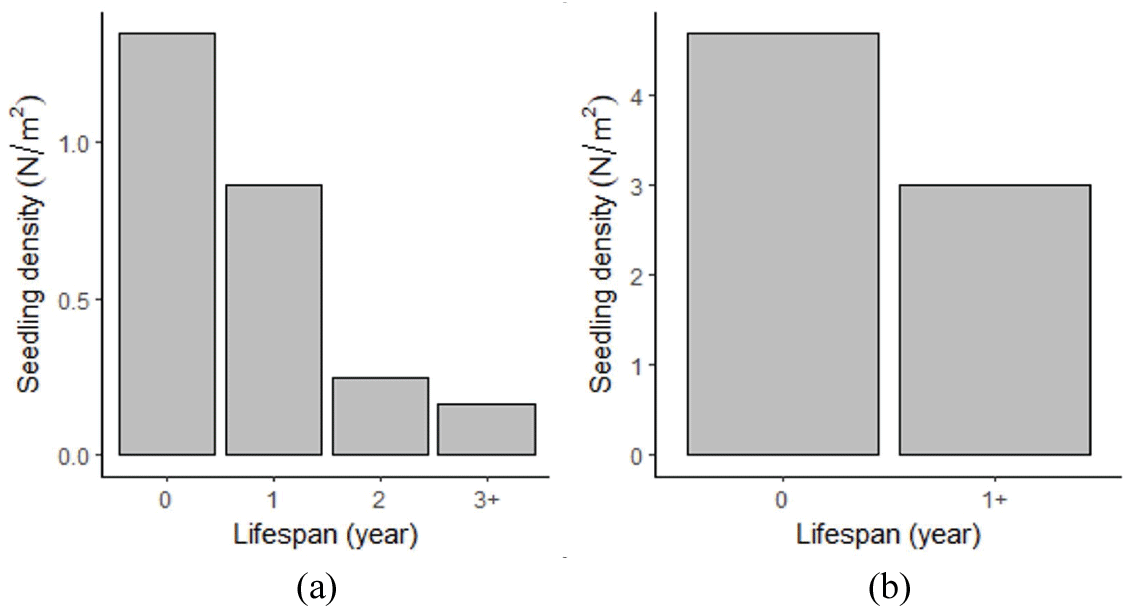
물푸레나무 치수는 발생 직후 다량 고사하였다. 2012년에 발생한 치수 집단의 밀도는 2.63본/m2였으며, 이 중 51% (1.35본/m2)가 그 해를 넘기지 못하고 고사했고, 33%(0.87본/m2)는 묘령 1년차인 2013년에 고사하여 1년 만에 사라졌다. 2012년 발생한 치수 집단의 9%(0.25본/m2)는 묘령 2년차인 2014년에 고사하여 2년의 수명을 마쳤고, 6% (1.76본/m2)만이 2년차인 2014년에 살아남았다[Figure 4(a)]. 2014년에 발생한 치수 또한 7.68본/m2 중 61%인 4.68본/m2이 발생 당해 고사했다. 즉, 과반 이상의 치수가 1년을 넘기지 못하고 고사하였다[Figure 4(b)].
Figure 4.
Lifespan distribution of Fraxinus rhynchophylla seedlings emerged in (a) 2012 and (b) 2014 observed in 2014. Seedling density on 0 year indicates number of seedlings/m2 that died in the same year of emergence. Lifespan refers to years of survival.
Download Original Figure
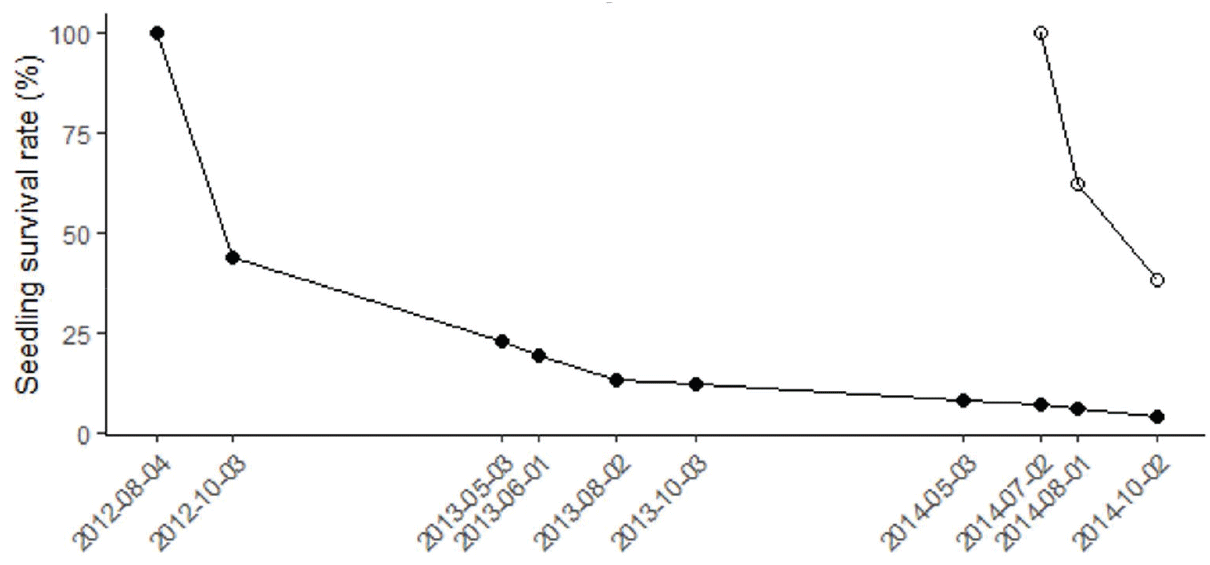
2012년 8월 발생이 관측된 발생 집단의 56%는 2개월 이후인 10월에는 사라졌다. 2014년 7월 발생이 관측된 발생 집단은 1개월 이내 38%가 고사하였고, 이후 2개월 동 강원도 가리왕산 지역 천연 활엽수림에서 물푸레나무(Fraxinus rhynchophylla)의 천연하종과 치수 발생 및 고사 특성 안 24%가 추가로 고사하여 발생 이후 3개월 동안 총 62%가 고사하였다(Figure 5).
Figure 5.
Survival rate of two emergence groups of Fraxinus rhynchophylla seedlings. ● and ○ indicate emergence groups that their first emergence was observed on 4 August 2012 and on 2 July 2014, respectively.
Download Original Figure
4. 하층 식생과 치수 고사
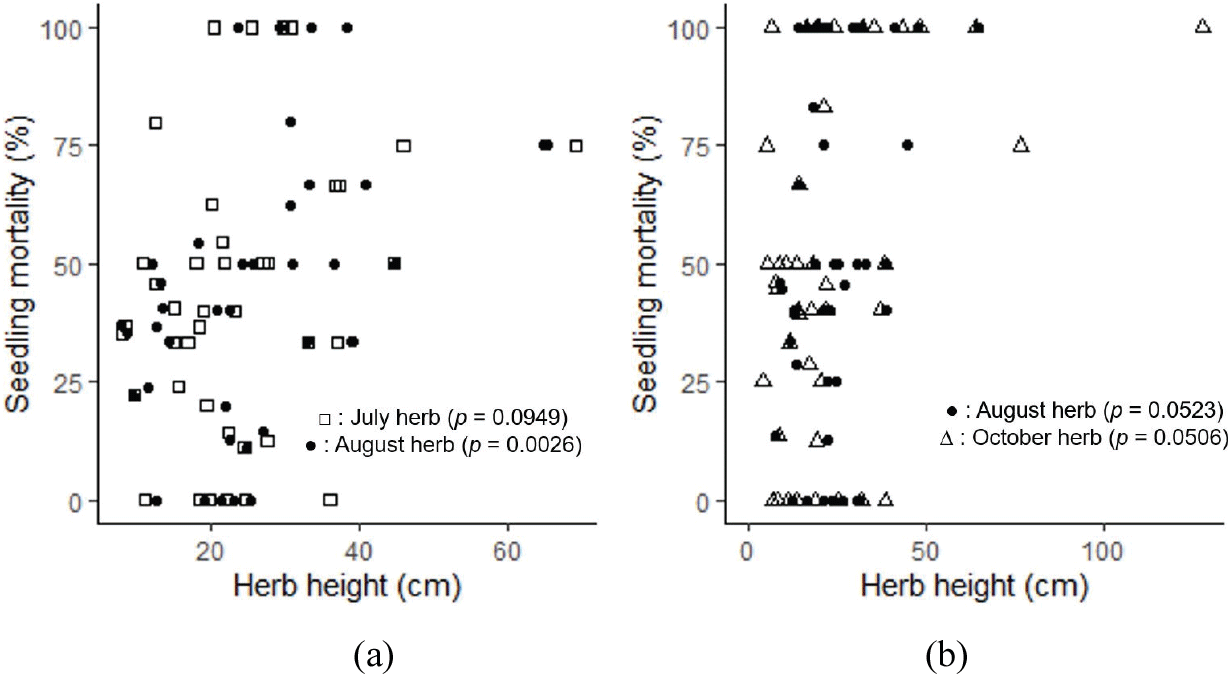
치수 발생 초기 여름철의 치수 고사율과 여름철 초본 식생의 초고는 양의 상관관계를 보였다. 2014년도에 발생한 치수 집단의 7~8월 사이의 고사율과 8월 초본 식생 초고가 유의한 상관관계를 보였다[p<.005; Figure 6(a)]. 8~10월 사이의 고사율과 8월 및 10월 초본 식생 초고 역시 양의 상관관계를 보였다[Figure 6(b)]. 발생 초기 치수의 평균 묘고는 6.1 cm, 근원경은 0.81 mm였다.
Figure 6.
Relationship between herb height and Fraxinus rhynchophylla seedling mortality rate. Mean mortality rate (a) from July to August 2014 and (b) from August to October 2014.
Download Original Figure
고 찰
1. 물푸레나무의 해거리와 연도별 치수 발생 변동
물푸레나무는 2011년, 2012년 낮은 결실 이후 2013년은 2011년 결실의 두 배 이상의 결실을 보였다(Figure 2). 물푸레나무는 천연하종갱신이 용이하지만 종자 생산은 심한 해거리를 하는 것으로 알려져 있다(Korea National Arboretum, 2016). 한반도 중부 천연림에서 물푸레나무는 3~4년의 해거리 주기가 보고되었으며(Sung, 1996; Sung et al., 2011; Korea National Arboretum, 2016), 스웨덴 활엽수림에서 F. excelsior는 3~6년의 결실주기를 보였다(Tapper, 1992; Tapper, 2006). 이처럼 물푸레나무를 비롯한 물푸레나무속 수종은 풍흉의 차가 심하고 3~4년 주기로 결실주기가 나타나는 것으로 알려져 있다.
물푸레나무속 수종의 치수 발생량은 하종의 풍흉에 크게 영향을 받는다. 2013년의 낮은 치수 발생은 2012년의 낮은 종자 생산에 기인한다(Figure 3). 또한 2013년의 낮은 치수 발생량은 2011년도에 하종된 다량의 종자가 2년 후의 치수 발생까지 이어지지 못했음을 의미한다. 이는 물푸레나무 종자는 생산된 연도에 하종하여 이듬해 높은 비율로 발아하여 치수가 발생하나, 그 이후에는 추가적으로 치수가 발생하지 않음을 보여준다.
스웨덴 활엽수림에서 F. excelsior 치수 발생량의 연도별 변동은 2년 전 하종량의 연도별 변동과 일치하는 경향을 보였으며, 이는 F. excelsior의 종자가 임상에 떨어진 후 2년 동안의 휴면 및 발아 촉진 기간을 거쳐 다량의 치수로 발생했기 때문이다(Tapper, 1992). 미국 오하이오 주 활엽수림의 물푸레나무속 치수는 10년 동안의 모니터링 기간 중 두 해만 유의미하게 높은 발생량을 보였는데 이는 물푸레나무속 특유의 해거리 경향이 치수 발생량의 연도별 변동을 초래한 것으로 볼 수 있다(Ralph and Brinkman, 1996).
2013년 종자 생산량과 2014년 치수 발생량은 높은 양의 상관관계를 보였다(Table 1). 본 연구와 동일한 지역에서 다양한 활엽수종의 실생묘 발생량을 비교한 선행연구에서는 물푸레나무 실생묘가 다른 수종들에 비해 치수 발생률이 높다고 보고되었다(Kim et al., 1999). 발아력이 높은 물푸레나무는 자연상태에서 천연하종갱신이 용이한 수종으로 평가받는다(Korea National Arboretum, 2016). 본 연구 조사지에 인접한 강원도 횡성에서 채취한 물푸레나무 종자를 대상으로 실험실에서 발아 실험을 수행한 결과 약 20°C의 온도에서 60% 이상의 발아율을 보였다(Choi et al., 2008). 종피에 상처를 낸 F. americana, F. pennsylvanica, F. angustifolia Vahl 종자에 대한 발아 실험에서는 80%가 넘는 발아율이 보고되었다(Preece et al., 1995). 본 연구에서 물푸레나무는 하종량 대비 70% 이상의 치수 발생률을 보여, 천연활엽수림에서 물푸레나무는 실험실 발아율과 큰 차이 없이 활발하게 치수가 발생함을 알 수 있다.
2. 종자 발아 및 치수 발생 시기
연구지의 물푸레나무 종자는 겨울을 지나 온도가 낮은 봄철까지 휴면을 유지하며 5~6월 이후 20°C 이상의 온도에 노출되면서 발아 및 치수 발생이 이루어진 것으로 볼 수 있다(Table 2). 물푸레나무 종자는 적정온도(20°C 내외)에서 높은 발아율을 보인다(Choi et al., 2008). 물푸레나무 속 종자는 충분한 수분 조건 하에서는 온도에 따라 배아 발달 비율 및 생장속도가 다르다. 5°C의 낮은 온도에서는 배아가 더디게 발달하고, 배아가 발달하기 시작한 이후에는 20°C의 온도에서 가장 활발히 생장한다(Villiers and Wareing, 1964). 연구지의 4월 평균온도는 6.0°C로 4월 중순에도 대기온도가 영하로 떨어지기도 한다. 5월 평균 대기온도는 12.9°C로, 5월 이후 대기온도가 5°C를 넘기 시작하고, 일 최대 대기온도가 20°C가 넘는 일수가 증가한다. 연구지에서 6월 중순에는 일평균 토양온도가 15°C이 넘는 날이 나타났다. 자연상태에서 하종된 물푸레나무 종자가 5월부터 20°C가 넘는 대기온도에 노출되고 6월에 발아에 용이한 환경이 조성되어 6-7월에 다량의 치수가 한꺼번에 발아했음을 알 수 있다. 저온습윤환경에서 휴면을 타파한 물푸레나무속 종자는 이후 적정 온도 조건에서 더욱 활발히 발아가 이루어진다(Villiers and Wareing, 1964; Finch-Savage and Clay, 1997).
3. 치수의 발생 초기 고사와 집단 동태
물푸레나무 치수는 절반 이상이 한 해를 넘기지 못하고 고사하였다(Figure 4, 5). 물푸레나무속 치수는 발생 직후 높은 고사율을 보인다. Fraxinus excelsior 치수는 발생 이후 2년 동안 높은 고사율을 보였으며, 임상의 식생 밀도가 높을수록 다수의 치수가 고사했다(Tapper, 1992). 오하이오 지역 혼효림에서 물푸레나무속 치수는 발생 후 1년 동안 96.9%의 치수가 고사하였고, 치수 수명의 중간값이 1년으로 매우 짧았다(Ralph and Brinkman, 1996). 하지만 발생 초기 살아남은 소수의 치수는 임상에 활착하여 장기간 생존한다(Grime and Hillier, 2000; Kitajima and Fenner, 2000). 본 연구의 물푸레나무 치수 또한 발생 초기에 높은 비율로 고사하였으며, 생존한 소수의 치수가 임상에 활착해서 갱신을 유지하는 것으로 볼 수 있다.
유년기의 높은 고사율과 이후 고사율의 감소로 인한 역J자 형태의 생존그래프는 K 선택 전략의 식물종에서 주로 나타난다(Manuel and Molles, 2015). K 선택 전략은 다량의 종자와 치수를 발생하여 교란과 스트레스 환경으로 인한 시공간적 불확실성에 대비하여 갱신을 도모한다(Grime, 1989). 본 연구에서 살아남은 물푸레나무 치수 집단 또한 초기 높은 고사 이후 생존곡선이 완만해지면서 살아남은 개체가 임상에 활착하고 줄기를 경화하여 생존하고, 이들 개체가 생존을 지속하며 전체적인 생존그래프가 K전략 식물종과 같은 역J자 형태로 나타나게 된다(Figure 5).
임상은 종자의 발아와 치수의 활착에 영향을 미친다(Keizer et al., 1985: Kitajima and Fenner, 2000). 본 연구지에서 암석 및 지의류로 이루어진 임상은 일부 종자가 발아하였으나 치수의 뿌리가 토양에 활착하지 못하고 대부분 고사하는 경향을 보였다.
4. 하층 식생이 치수 생존에 미치는 영향
하층 초본 식생의 높은 초고는 물푸레나무 치수의 고사와 양의 상관관계가 있는 것으로 나타났다(Figure 6). 하층 초본식생이 번성하면 치수가 요구하는 광을 차단하여 치수의 갱신에 부정적으로 작용하며 임목의 갱신을 억제한다(Looney et al., 2017). 또한 초고가 높은 식생은 치수를 피음하여 치수 생장에 부정적 영향을 끼친다(Iwamoto and Sano, 1998; Park et al., 2012). 경북 봉화군 소나무 모수림 시업지에서 천연하종으로 발생한 소나무 치수는 시업 초기 풀베기작업이 필요하였다(Byeon and Yun, 2018). 본 연구에서도 초고가 높은 초본식생이 치수가 필요로 하는 광을 차단해 초기 고사율을 높인 것으로 보인다. 조릿대가 우점하는 임상 또한 치수 발생이 거의 없었다. 밀도가 높은 조릿대가 임상에 도달하는 광을 차단하여 치수의 생장을 제한한 것으로 보인다.
지하부 경쟁 역시 물푸레나무 치수의 활착을 방해하는 것으로 알려졌는데(Bloor et al., 2008), 영국 Buff Wood 및 Harlton Cambridgeshire에서 채취한 발아 직후의 F. excelsior 근계는 단일 직근이 주로 발달하여 측근이 미약하고 발달 정도가 지상부보다 부족하였다(Wardle, 1961; Thomas, 2016). 발달이 미약한 치수의 근계는 낙엽 및 하층 식생에 의해 쉽게 임상에서 이탈된다. 연구지의 초본 식생은 지상부의 광 뿐 아니라 지하부의 공간을 점유하여 물푸레나무 치수의 생존을 방해하였다. 이렇듯 울폐된 자연 임상에서 물푸레나무 치수는 높은 고사율로 인해 천연하종갱신이 어렵지만, 수관을 조절하여 빛 환경을 개선하고 임상에 바닥긁기(Scarification)를 실시하여 하층 환경을 개선하면 갱신을 도울 수 있을 것이다(Sung et al., 2011).
결 론
본 연구는 강원도 천연 활엽수림에 자생하는 물푸레나무 개체군의 해거리 경향, 천연 갱신된 치수의 발생 및 생존을 분석하여 천연림에서 물푸레나무 치수의 발생과 생존 특성을 파악하고자 하였다. 3년 동안 물푸레나무는 종자 생산량은 해마다 달랐으며, 해거리 경향을 보였다. 종자 하종은 이듬해 치수 발생으로 이어져 종자 생산량과 이듬해 치수 발생량은 양의 상관관계를 보였다. 물푸레나무 치수는 종자 하종 이듬해 초여름(5~7월) 온도가 상승함에 따라 종자가 발아하여 치수가 발생했다. 임상에서 물푸레나무는 하종량 대비 이듬해 치수 발생 비율이 최대 76.2%에 달했고, 하종량의 변동은 이듬해 여름 치수 발생량에 직접적으로 영향하였다. 치수 발생 직후 다량의 치수가 고사하여 당해년도에 치수 고사율이 높았다. 초본 식생의 초고는 피음 환경을 조성하여 치수의 생장에 부정적으로 작용하며 고사율에 영향을 주었다. 자연 상태의 물푸레나무는 해거리로 특정 해에 다량의 종자를 임상에 공급하며, 이는 이듬해 높은 치수 발생으로 이어지나 초기 고사율이 높았고, 울폐한 하층 식생은 물푸레나무 치수 생존에 부정적으로 작용하였다. 천연활엽림에서 물푸레나무의 천연하종갱신을 위해서는 종자의 다량 생산이 이루어지는 이듬해 임상의 광 환경 및 하층 환경을 개선하여 치수의 생존률을 높일 필요가 있다. 본 연구는 3년에 걸친 하종량 조사 기간 동안 물푸레나무 종자 생산량의 풍흉을 확인하였다. 이후 지속적인 결실량 모니터링을 통해 기존에 보고된 물푸레나무의 3~4년 해거리 주기를 환경 영향과 연계하여 확인할 필요가 있다. 또한 향후 수종의 천연갱신에 대한 연구를 수행하기 위해서는 목표 수종의 생태적 특성, 종자의 크기와 모양, 치수의 특징 및 각 개체에 대한 모니터링 방안을 수립하는 것이 중요하다. 특히 발아한 지 얼마 안 된 치수는 임상의 다른 식생과 구별이 어려워 관찰과 주의가 요구된다.
감사의 글
본 연구는 산림청(한국임업진흥원) 산림과학기술 연구개발사업(S211314L020120, FTIS 2022461B10-2324-0201)과 국립산림과학원 위탁연구용역의 지원을 받았습니다. 조사를 도와주신 분들과 연구지 출입에 협조해주신 평창국유림관리소에 감사드립니다.
References
Bloor, J.M.G., Leadley, P.W. and Barthes, L. 2008. Responses of
Fraxinus excelsior seedlings to grass-induced above-and below-ground competition. Plant Ecology 194: 293-304.


Byeon, S.Y. and Yun, C.W. 2018. Understory species composition and
Pinus densiflora natural regeneration in
Pinus densiflora stands regenerated by seed-tree method. Journal of Korean Forest Society 107: 25-34.
Choi, C.H., Seo, B.S., Tak, W.S., Cho, K.J., Kim, C.S. and Han, S.U. 2008. Comparison of seed germination response to temperature by provenances in
Fraxinus rhynchophylla. Journal of Korean Forest Society 97(6): 576-581.
Eastern Regional Forest Service. 2009. Report on Development of Eco-Friendly Silvicultural Techniques and Sustainable Forest Management Techniques [X]. Eastern Regional Forest Service, Gangneung. pp. 198.
Finch-Savage, W.E. and Clay, H.A. 1997. The Influence of embryo restraint during dormancy loss and germination of
Fraxinus excelsior seeds. pp. 245-253. In: Ellis, R.H., Black, M., Murdoch, A.J. and Hong, T.D. (Ed.). Basic and Applied Aspects of Seed Biology: Proceedings of the Fifth International Workshop on Seeds. Springer. Dordrecht, Netherlands.
Grime, J.P. 1989. The stress debate: symptom of impending synthesis? Biological Journal of the Linnean Society 37(1-2): 3-17.
Grime, J.P. and Hillier, S.H. 2000. The contribution of seedling regeneration to the structure and dynamics of plant communities, ecosystems and larger units of the landscape. pp. 361-374. In: Fenner, M. (Ed.). Seeds: The Ecology of Regeneration in Plant Communities. CABI. London, England.
Iwamoto, S. and Sano, J. 1998. Standing crop of
Sasa and the growth patterns of seedlings in a deciduous broad-leaved secondary forest. Nihon Ringakkai Shi. Journal of the Japanese Forestry Society 80: 311-318.
Keizer, P.J., van Tooren, B.F. and During, H.J. 1985. Effects of Bryophytes on seedling emergence and establishment of short-lived forbs in Chalk Grassland. Journal of Ecology 73: 493–504.
Kim, C.H., Oh, J.G., Kang, E.O., Yun, C.S., and Lim, J.K. 2014. Community distribution on mountain forest vegetation of the Gyebangsan Area in the Odaesan National Park, Korea. Korean Journal of Ecology and Environment 47(3): 135–145.
Kim, G-T., Choo, G-C. and Baek, G-J. 2003. Structure of forest community at Daedeoksan-Geumdaebong Nature Ecosystem Preservation Area in Baekdudaegan. Korean Journal of Environment and Ecology 17: 9-17.
Kim, H.J., Jung, J.B., Sung, J.H., Han, A.R. and Park, P.S. 2015. Flowering phenology and the growth of three native Anemone species in a montane deciduous forest on Mt. Joongwang, Korea. Horticulture, Environment, and Biotechnology 56: 849-857.
Kim, J.H., Yang, H.M. and Jin, G. Z. 1999. The pattern of natural regeneration by three different silvicultural systems in a natural deciduous forest. Journal of Korean Forest Society 88(2): 169-178.
Kim, J.H., Yang, H.M., Jin, G.Z., Lee, W.S. and Kang, S.K. 2001. The aspect of natural regeneration for major tree species in the natural deciduous forest. Journal of Forest Science, Kangwon National University 17(1): 1-17.
Kimmins, J. P. 2004. Forest Ecology: a foundation for sustainable forest management and environmental ethics in forestry. 3
rd Ed. Prentice Hall. New Jersey, USA. pp. 611.
Kitajima, K. and Fenner, M. 2000. Ecology of seedling regeneration. pp. 331-359. In: Fenner, M. (Ed.). Seeds: The Ecology of Regeneration in Plant Communities. CABI. London, England.
Koenig, W.D. 2021. A briefhistory of masting research. Philosophical Transactions the Royal Society of Publishing B 376: 20200423.
https://doi.org/10.1098/rstb. (2020. 4. 23).
Korea Forest Service. 1999. Report on Demonstration Research for the Improvement of National Forest Management by Industrial and Academic Cooperation (X). Korea Forest Service, Daejeon.
Korea Institute of Geoscience and Mineral Resources. 2022. Geo Big Data Open Platform: Pyeongchang.
https://data.kigam.re.kr/map/. (2022. 5. 22).
Küßner, R. 2003. Mortality patterns of
Quercus,
Tilia, and
Fraxinus germinants in a floodplain forest on the river Elbe, Germany. Forest Ecology and Management 173: 37-48.
Kwon, H.J., Kim, J.W., Cha, D.W. and Oh, C.H. 2018. Characteristics of plants according to the management characteristics of “Nationally Important Agricultural Heritage” Uljin Geumgang Forest region. Proceedings of Korean Society of Envrionment and Ecology Confernce, Kongju National University, Gongju, 2018. 10. 19. pp. 4-5.
Lee, D.K., Kwon, K.W., Kim, J.H. and Kim, G.T. 2010. Silviculture: sustainable ecological management of forests. Hyangmunsa. Seoul, Republic of Korea.
Looney, C.E., D′Amato, A.W., Palik, B.J., Slesak, R.A. and Slater, M.A. 2017. The response of
Fraxinus nigra forest ground-layer vegetation to emulated emerald ash borer mortality and management strategies in northern Minnesota, USA. Forest Ecology and Management 389: 352-363.
Manuel, J. and Molles, C. 2015. Ecology: concepts and applications. McGraw-Hill Education. New York, USA.
Moles, A.T. and Westoby, M. 2004. Seedling survival and seed size: a synthesis of the literature. Journal of Ecology 92(3): 372-383.
National Geographic Information Institute. 2016. National Atlas: 2. National Geographic Information institute, Ministry of Land, Infrastructure and Transport. Suwon, Korea.
Park, P.S., Kim, K.Y., Han, A., Jang, W., Son, Y., Yi, M. J., Park, B.B. and Son, Y. 2010. Development processes and growth pattern of
Pinus densiflora stands in central eastern Korea. Journal of Plant Research 123: 453-462.

Park, S.G., Yi, M.H., Yoon, J.W. and Sin, H.T. 2012. Environmental factors and growth properties of
Sasa borealis (Hack.) Makino community and effect its distribution on the development of lower vegetation in Jirisan National Park. Korean Journal of Environment and Ecology 26(1): 82-90.
Preece, J., Bates, S. and Sharon V.S. 1995. Germination of cut seeds and seedling growth of ash (
Fraxinus spp.) in vitro. Canadian Journal of Forest Research 25(8): 1368-1374.
Putz, F.E. and Canham, C.D. 1992. Mechanisms of arrested succession in shrublands: root and shoot competition between shrubs and tree seedlings. Forest Ecology and Management 49(3-4): 267-275.
Ralph, E.J.B. and Brinkman, J.A. 1996. Ten years of tree seedling establishment and mortality in an Ohio deciduous forest complex. Bulletin of the Torrey Botanical Club 123(4): 309-317.
Sung, J.H. 1996. Ecophysiological Characteristics for Natural Regeneration of
Fraxinus rhynchophylla hance. (Dissertation). Seoul National University. Seoul, Republic of Korea.
Sung, J.H., Lee, D.K. and Park, P.S. 2011. Seed biology, stand characteristics, and regeneration of
Fraxinus rhynchophylla Hance. Journal of Sustainable Forestry 30(5): 392-405.
Tanouchi, H., Sato, T. and Takechita, K. 1994. Comparative studies on acorn and seedling dynamics of four
Quercus species in an evergreen broad-leaved forest. Journal of Plant Research 107: 153-159.
Tapper, P.G. 1992. Demography of persistent juveniles in
Fraxinus excelsior. Ecography 15(4): 385-392.
Tapper, P.G. 2006. The replacement of
Alnus glutinosa by
Fraxinus excelsior during succession related to regenerative differences. Ecography 16(3): 212-218.
Thomas, P.A. 2016. Biological flora of the British Isles:
Fraxinus excelsior. Journal of Ecology 104(4): 1158-1209.
Villiers, T.A. and Wareing, P.F. 1964. Dormancy in Fruits of
Fraxinus excelsior L.. Journal of Experimental Botany 15(2): 359-367.
Wardle, P. 1961. Biological Flora of the British Isles:
Fraxinus excelsior L. Journal of Ecology 49(3): 739-751.