



서 론
세계의 산림은 역사적으로 인구 증가, 농경지 확대, 도시화, 자원수탈, 산업협명, 전쟁, 산불, 병해충 피해 등 다양한 원인으로 많이 훼손되어 왔고(Lindsey, 2007; Pacheco et al., 2021), 최근에도 황폐화는 계속되고 있다. 2001년 이후에도 20년간 벌목, 산불, 농경지 확장 등으로 전세계 산림의 7%가 소실되었고, 연간 산림 소실률도 2001~2009년에 비해 2010~2018년에 50%나 급증하였다(He et al., 2023). 이와 같은 오랜 시간에 걸친 산림의 훼손으로 세계는 기후변화, 생태계 파괴, 생물다양성 감소, 환경오염 그리고 생활환경 악화와 같은 문제에 직면했다(Pacheco et al., 2021). 이에 인류는 산림복원을 통해 이러한 문제들을 개선하고자 시도하고, 산림 면적 확대를 위한 본 챌린지(Bonn challenge)와 같은 범국가적인 노력을 기울이고 있다. 우리나라의 산림도 외세에 의한 자원수탈, 과도한 벌채와 전쟁 등으로 황폐화 되었으나, 1970년대 이후 정부 주도의 집중적인 치산녹화 사업으로 녹화에 성공하였다(Bae, 2013; Korea Forest Service, 2020). 그러나 단기간의 인공조림을 통한 산림 조성으로 몇몇 수종들의 단순 동령림 형태를 보이고, 2020년 기준으로 전체 산림면적에서 Ⅳ·Ⅴ영급이 75.4 %, Ⅰ·Ⅱ·Ⅲ영급이 18.8 %를 차지하는 심각한 영급불균형 상태를 보이고 있다(Korea Forest Service, 2022). 하지만, 최근에 치산녹화 사업으로 조성된 산림이 점차 벌기에 도달하여 목재수확과 재조림이 이루어지고 있어 현재 시기는 산림생태계와 지속가능한 산림경영에 적합하도록 영급불균형을 조절할 수 있는 시기이다. 따라서 산림청에서는 재조림 방법으로 기존에 사용하던 인공조림과 더불어 천연하종갱신과 맹아갱신 같은 다양한 조림 방법으로 재조림을 계획하고 있으나 아직까지는 과거와 변함없이 양묘된 묘목을 일괄 식재하는 인공조림에 그대로 의존하고 있다(Korea Forest Service, 2019).
천연갱신은 지역에 적응된 수종으로 건강한 산림을 조성할 수 있고, 인공조림의 약 절반의 조림비용으로 재조림 할 수 있는 방법이다(Chokkalingam et al., 2018). 유럽에서는 천연갱신이 100여 년 전부터 많이 이루어져 왔고 지역 특성과 지형에 맞는 다양한 방법이 경제적, 생태적 그리고 기술적 관점에서 연구되어 실무에 이용되고 있으며(Kim et al., 2014), 일본도 2034년까지 조림면적의 48.2 %를 천연갱신으로 하겠다는 계획을 시행하고 있다(Ministry of Agriculture, Forestry and Fisheries, 2018). 그러나 일부 국가를 제외한 대다수의 국가에서는 많은 장점에도 불구하고 천연갱신이 인공조림에 비해 상대적으로 오랜 시간과 많은 지식 및 기술을 필요로 하기에 제대로 사용하지 못하고 성공 가능성에 대한 두려움도 가지고 있는 실정이다.
우리나라 산림 자원 중에 질적, 양적으로 높은 가치를 지니고 있는 소나무림의 갱신도 과거에는 천연갱신이 주를 이루었으나 최근에는 인공조림이 주를 이루고 있으며 산림경영을 위한 천연갱신 시업 및 관리방안에 관한 연구도 부족한 실정이다. 우리나라의 소나무 천연갱신 연구(Hyun, 1943; Kim et al., 1987; Kim et al., 1989; Bae, 1994; Park and Lee, 1996; Na et al., 2010)는 일부 시도되었으나 종합적이거나 지속적으로 이어지지 못하였다. 그러나 우리나라 산림의 수확에 따른 재조림과 산림 재해에 의한 산림복구 대상지는 앞으로도 계속 증가될 것으로 예상되고 있어 소나무에 대한 천연갱신 기술 확보는 매우 시급한 실정이다. 따라서 본 연구는 소나무 천연갱신에 필요한 중요 요소인 천연갱신 대상지의 종자 결실과 공급에 관한 기작을 구명하고자 시행되었고, 소나무 천연하종 시험지를 조성하여 종자 유입량 및 유입 패턴 등을 조사·분석하였다. 종자 비산에 관한 정밀한 연구를 위해서는 공간 패턴을 측정하고 다양한 모델 개발 등이 필요하지만(Levin et al., 2003), 본 연구는 천연갱신 처리 방법에 따른 차이와 효과 분석을 주목적으로 하기에 가장 일반적인 현상학적 방법을 이용하여 연구를 수행하였고, 우리나라 소나무 천연갱신 시업체계 마련을 위한 기초 정보를 제공하고자 한다.
재료 및 방법
소나무(Pinus densiflora Siebold & Zucc) 천연갱신 시험을 위해 임분의 생육상태와 형질이 양호한 강원도 삼척시 하장면 갈전리의 소나무 임분을 시험지로 선정하였다. 임분조사는 반경 10 m의 원형 표본 조사구(314 m2)를 시험지 계획을 고려하여 배치하고 13개소를 조사하였다. 임황조사는 수종, 흉고직경, 수고, 지하고, 수관폭 등을 조사하였고 지황은 지형, 경사, 방위 등을 조사하고 토양시료를 채취하여 분석하였다. 시험지는 해발 630~700 m, 경사 25~35°의 남서향 사면에 위치하고, 토성은 미사질양토이다(Table 1). 연구대상지 임황은 Ⅵ~Ⅸ영급 소나무 임분으로 임분밀도는 평균 409 본/ha이고, 평균 흉고직경은 27.4 cm, 평균 수고는 19.5 m이다(Table 2).
| Stand density (tree ha−1) | Average DBH (cm) | Average height (m) | Basal area (m2 ha−1) | Volume (m3 ha−1) | Total crown area (m2 ha−1) | Stand age-classa |
|---|---|---|---|---|---|---|
| 409 | 27.4 | 19.5 | 28.4 | 219.2 | 22,535 | VI~IX |
| 7.3~62.5 | 7.9~21.9 |
대상지의 기후는 시험지와 직선거리로 약 600 m 떨어진 곳에 위치한 기상청(Korea Meteorological Administration) 하장 자동기상관측시스템(Automatic Weather System, AWS)에서 관측된 10년간(2004~2013)의 자료를 분석하였다. 그 결과, 연평균 기온은 8.7°C로 우리나라 연평균기온 보다 4°C 낮았고, 연평균강수량은 1,223 mm로 우리나라 평균과 유사하였다. 소나무 종자의 비산시기로 알려진 9~11월의 풍향은 풍속이 0.5 m/s 미만인 정온이 약 28 %로 가장 많았고 서쪽과 북쪽에서 부는 바람이 약 40 %로 나타났다. 풍속은 서풍이 가장 강했고 최대풍속은 약 5.7 m/s이었다(Figure 1).

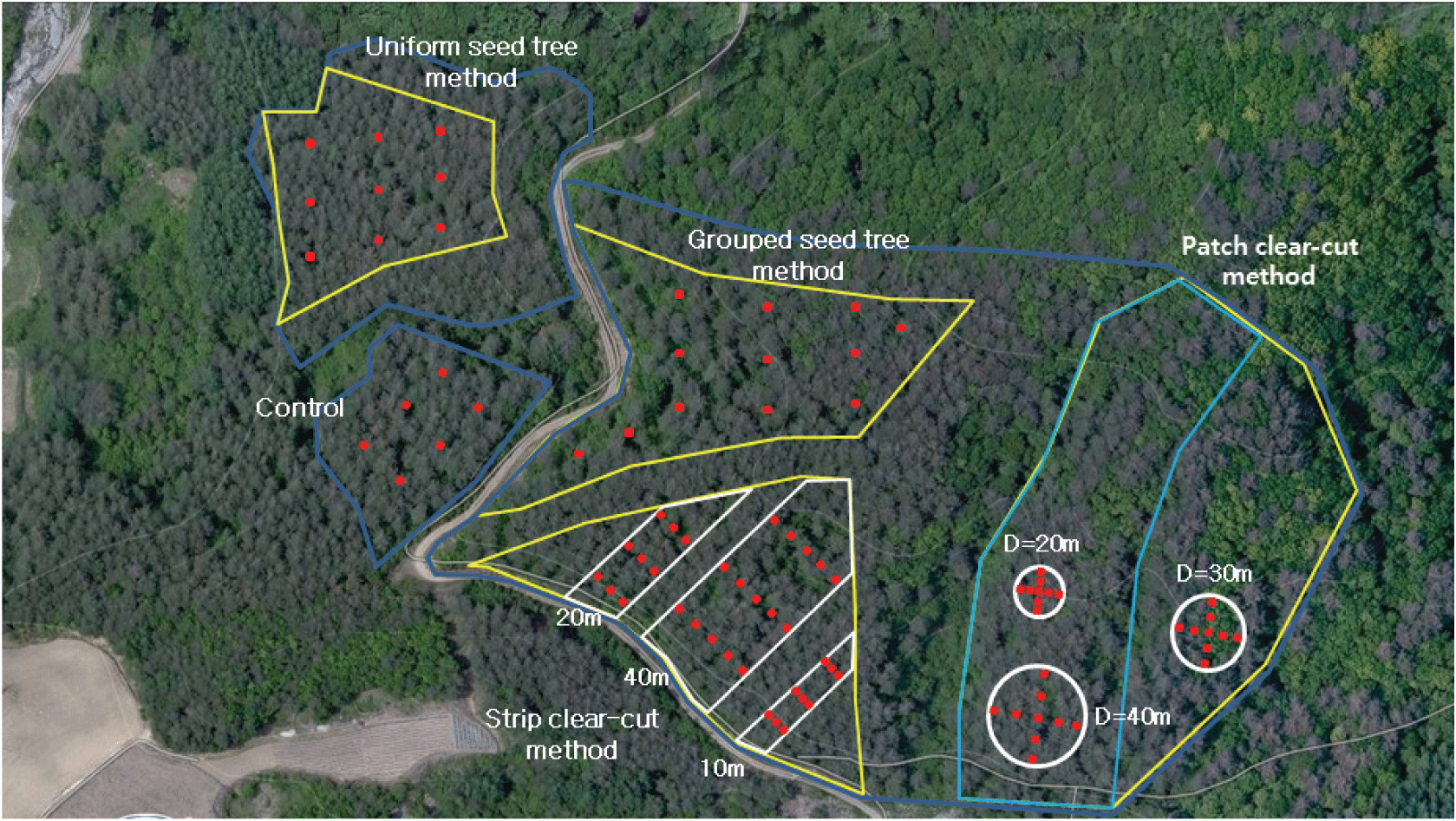
소나무림 8.6 ha를 대상으로 2012년도에 단목 모수작업, 3본 군상 모수작업, 폭 10 m 대상개벌작업, 폭 20 m 대상개벌작업, 폭 40 m 대상개벌작업, 직경 20 m 군상개벌작업, 직경 30 m 군상개벌작업, 직경 40 m 군상개벌작업을 실시하고 무처리구를 배치하였다(Table 3).
연구대상지 내 소나무 천연갱신 처리에 따른 유입량은 직경 56.5 cm(단면적 0.25 m2)의 원형 수집망을 지표면으로부터 50 cm 높이에 설치하였다. 수집망은 처리별 대상지의 크기와 유형을 고려하여 총 87개(모수작업 21개, 대상개벌 33개, 군상개벌 27개, 대조구 6개)를 배치하였다(Figure 2). 종자 산포 연구를 위해서는 공간 패턴 측정과 모델 개발 등이 필요하고 실험 설계에서 임분 특성, 입지 트성, 기상환경, 그리고 적용 모델 등에 따라 종자 수집망의 설치 형태, 간격, 범위, 수집망 개수 등이 정해진다(Leadem et al., 1997; Nathan et al., 2002; Levin et al., 2003; Vergne, 2016). 본 연구지는 천연갱신 방법에 따른 효과 분석을 위해 조성되어 정밀한 종자 산포 모델 개발에는 적합하지 않고 천연갱신 유형별 천연갱신지의 종자 유입 차이 분석을 위하여 갱신상 전체를 대상으로 수집망을 설치하였다. 천연갱신 유형별 갱신상의 형태와 크기가 다양하여 수집망은 거리보다는 갱신상을 위치별 공간으로 분할하여 배치하였다. 사면향을 기준으로 군상개벌은 종자수집망을 십자형태로 배치하였고 나머지 갱신상에는 사면 경사 위치에 따라 상·중·하로 나누고 각 중심부와 등고선을 따라 좌우 임연부 경계의 소나무 수관부 아래에 수집망을 설치하였다. 개벌 폭이 넓은 40 m 대상 개벌 작업지와 사면 길이가 긴 군상 모수 작업지에는 수집망을 추가 배치하였다. 동일 시험지에서 본 연구와 연계하여 이루어질 천연갱신 치수의 발생과 생장 연구를 위해 하종 종자의 인위적인 유출을 최소화하려고 하였고 수집망 총 87개(모수작업 21개, 대상개벌 33개, 군상개벌 27개, 대조구 6개)를 제한적으로 배치하였다(Figure 2). 종자수집은 2014년 9월부터 2017년 2월까지는 종자비산 예상 시기인 9월부터 이듬해 2월까지 매월말 정기적으로 종자수집을 실시하였고, 2017년 9월부터 2020년 2월까지는 매년 9월초에 종자수집망을 정비하고 이듬해 2월말에 수집된 종자를 수거하여 대상지에 유입된 종자량을 집계하였다.
종자 유입량은 산술평균을 구하고 처리간 차이의 유의성은 t-검정(t-test)이나 분산분석(ANOVA)을 이용하여 검증하였으며, 통계적으로 유의성이 인정되는 평균값의 차이는 Duncan multiple range test (p<0.05)를 통해 비교하였다. 연도별, 목재수확 유형별 유입 종자의 분포 균일성 비교는 상대적인 산포도를 비교하기 위한 변이계수(Coefficient of variation, CV; 식 1)를 사용하였으며, 통계분석 프로그램은 R 4.1.1을 이용하였다.
(CV: Coefficient of variation, δ: Standard deviation, χ: arithmetic mean)
결과 및 고찰
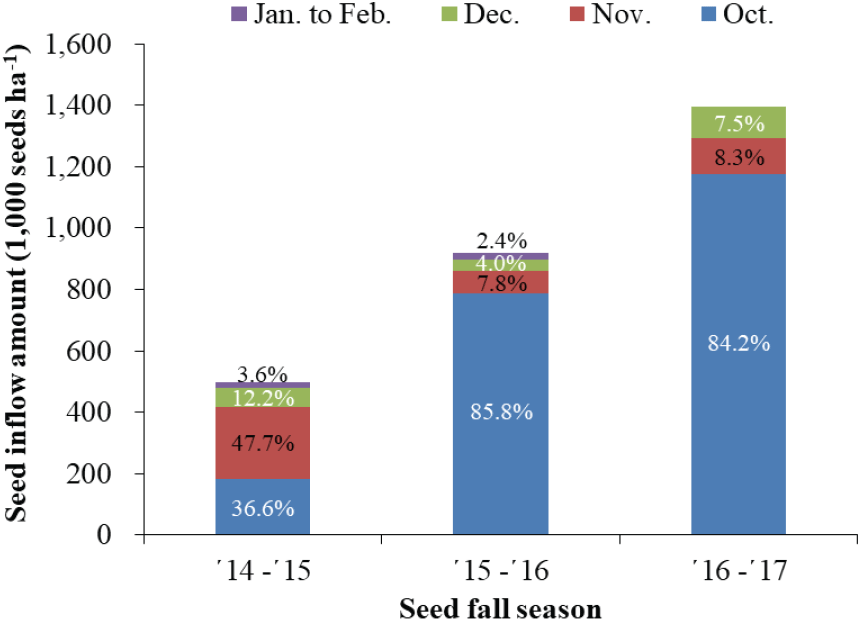
소나무 구과의 성숙 시기는 9월이고 종자채취 시기는 9월 중순 이후로 알려져 있는데(Burns and Honkala, 1990; Kim et al., 2014), 본 연구에서는 종자 비산 시기는 10월부터 이듬해 2월까지로 조사되었다(Figure 3). 성숙된 소나무 종자는 ’14~’15년도와 ’15~’16년도에는 10월에서 이듬해 2월까지 비산되었고, ’16~’17년도에는 10월부터 12월까지 비산되었다. 종자의 성숙은 지역별, 위치별 그리고 기상 상태에 따라 달라질 수 있고, 성숙한 구과에서의 종자 비산도 대기습도에 따라 질 수 있어 비산 시기는 일부 차이가 발생할 수 있다(Kim et al., 2014; Song et al., 2015). 월별 하종량을 조사한 3년 동안의 월별 하종량은 10월과 11월에 80 % 이상이 하종되어 종자 비산이 시작된 이후 1~2개월 동안의 기간에 집중되었고, 상대적으로 종자 결실이 양호했던 ’15~’16년도와 ’16~’17년도에는 종자 비산이 시작된 10월 한 달 동안에 약 85 %의 소나무 종자가 비산된 것으로 조사되었다. 종자 비산 시작 시기의 차이는 있지만 Bonner and Karrfalt(2008)의 9월~10월에 종자가 비산 된다는 보고와 유사하였다.
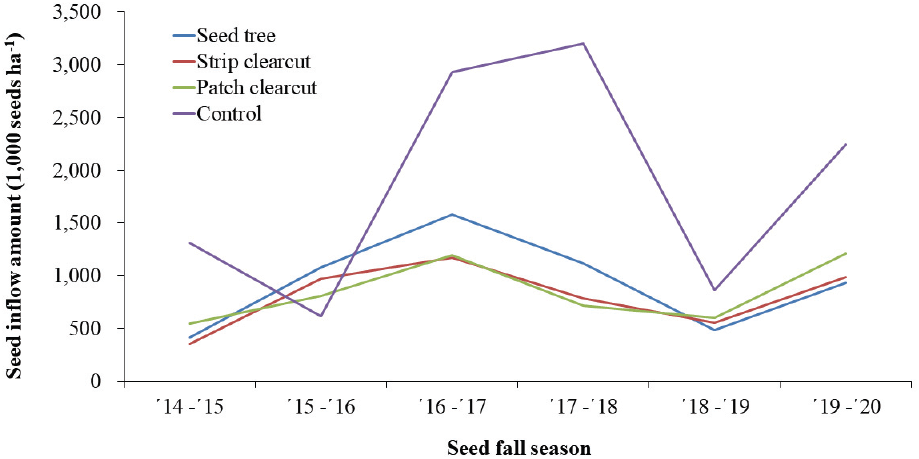
소나무는 매년 결실하지만 2~3년의 풍흉 주기를 갖는다(Kim et al., 2014). 목재수확 유형별 종자 유입량은 차이가 있었지만, 결실 풍흉에 따라 종자 유입량 변화 경향은 유사하였다(Figure 4). 그러나 목재수확이 이루어지지 않은 대조구는 처리구와 다른 경향을 보였다. 양수성 수종 성숙림의 갱신 능력은 임령보다 울폐도에 따라 달라지는 광조건의 영향이 크고(Wang and Zhou, 1989), 소나무과 수종은 솎아베기나 벌채 등으로 입목의 광합성과 영양상태가 양호해지면 개화 촉진과 종자생산량이 증가했으며(Sarvas, 1963; Cooley, 1970; Yocom, 1971; Croker, 1973), Red pine (Pinus resinosa Ait.)는 솎아베기 강도가 높을수록 종자 생산 개체 비율이 증가하였다(Godman, 1962). 목본식물은 화아원기 형성에서 종자 성숙까지 걸리는 2~3년의 기간동안 태양복사량과 강우량의 영향을 크게 받으며 여름의 더위는 암꽃 생산이나 개화를 촉진하기에(Bonnet-Masimbert, 1978) 소나무 천연갱신 시험지의 벌채에 따른 수광량 변화와 함께 임분 내 기류변화, 수관 발달 등 생육환경 변화에 따른 영향으로 처리구와 대조구 간에 다른 종자결실 경향을 보이는 것으로 사료된다.
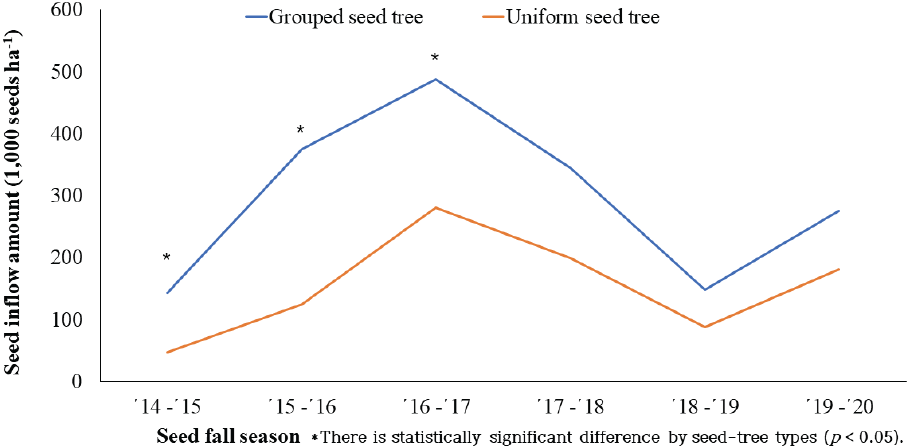
2014년 9월부터 2020년 3월까지 목재수확 유형별 종자 유입 경향을 분석한 결과, 소나무 천연갱신 모수작업지(Figure 5)는 군상작업지가 단목작업지 보다 종자유입량이 많았고 종자 결실이 좋았던 ’16~’17년도에는 작업 유형간 208 천립/ha, 결실이 나빴던 ’18~’19년도에는 61 천립/ha의 종자 유입량 차이를 보였다. 이것은 벌채지 종자공급원의 결실량 차이와 벌채지의 면적 그리고 벌채지와 접하는 임연부의 길이에 따른 차이로 판단되었다.
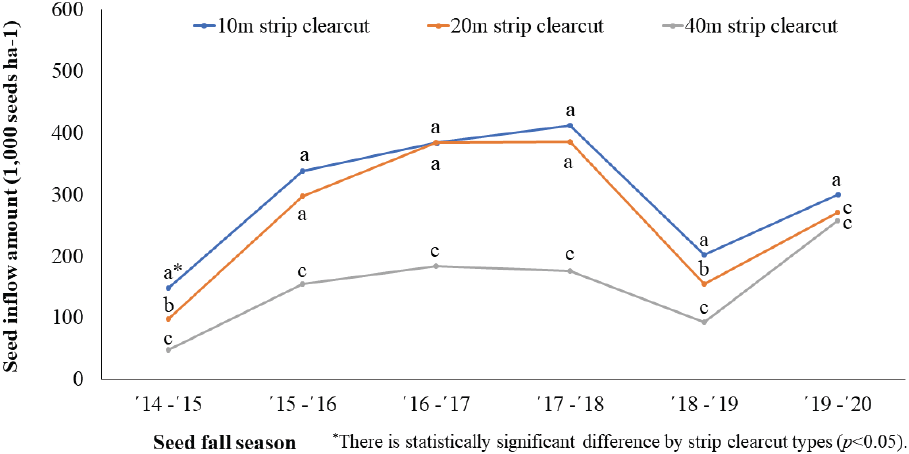
대상 개벌작업지는 10 m 대상 개벌작업지의 종자 유입량이 가장 많았고 벌채 폭에 따라 20 m, 40 m 대상 개벌작업지 순으로 나타났다(Figure 6). 처리간 종자 유입량은 10 m와 20 m 대상 개벌작업지는 유사한 결과를 보였으나 벌채폭이 넓은 40 m 대상 개벌작업지는 차이를 보이는 것으로 나타났다. Lee et al.(2018)은 소나무 종자는 80 m까지도 비산되지만, 종자 공급원으로부터 20 m까지 기하급수적으로 감소하여 40 m까지는 유사한 도달 종자의 수를 보고하였다.
군상 개벌작업지는 직경 20 m 군상 개벌작업지의 종자 유입량이 가장 많았고 벌채 폭 크기에 따라 30 m, 40 m 군상 개벌작업지 순으로 나타나 대상 개벌작업지와 동일하게 벌채 크기 증가에 따른 종자 유입량 감소 경향을 보였다(Figure 7). 종자 결실이 좋았던 ’16~’17년도에는 직경 20 m 처리구가 직경 40 m 처리구에 비해 종자 유입량이 1.9배 높았으며, 결실이 나빴던 해에는 비슷한 것으로 나타나 종자 풍흉에 따라 모수로부터 일본잎갈나무의 종자 비산거리와 밀도가 변화한다는 Masahiko et al.(2016)의 연구 결과와 유사하였다.
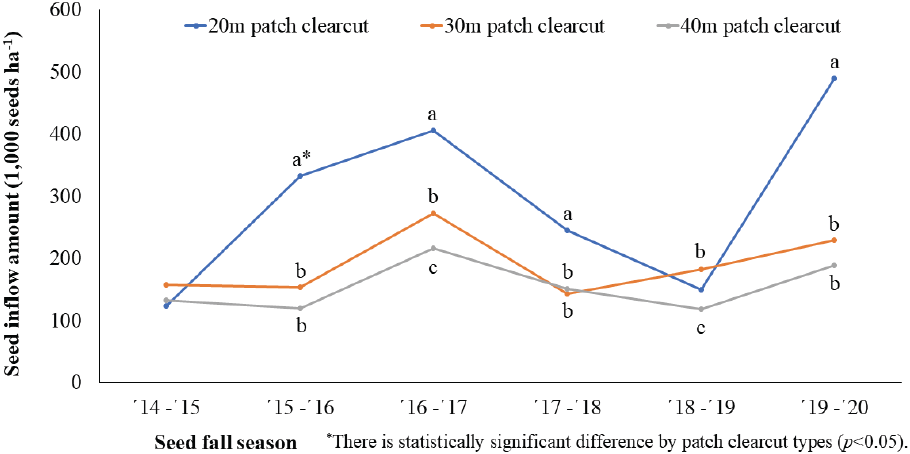
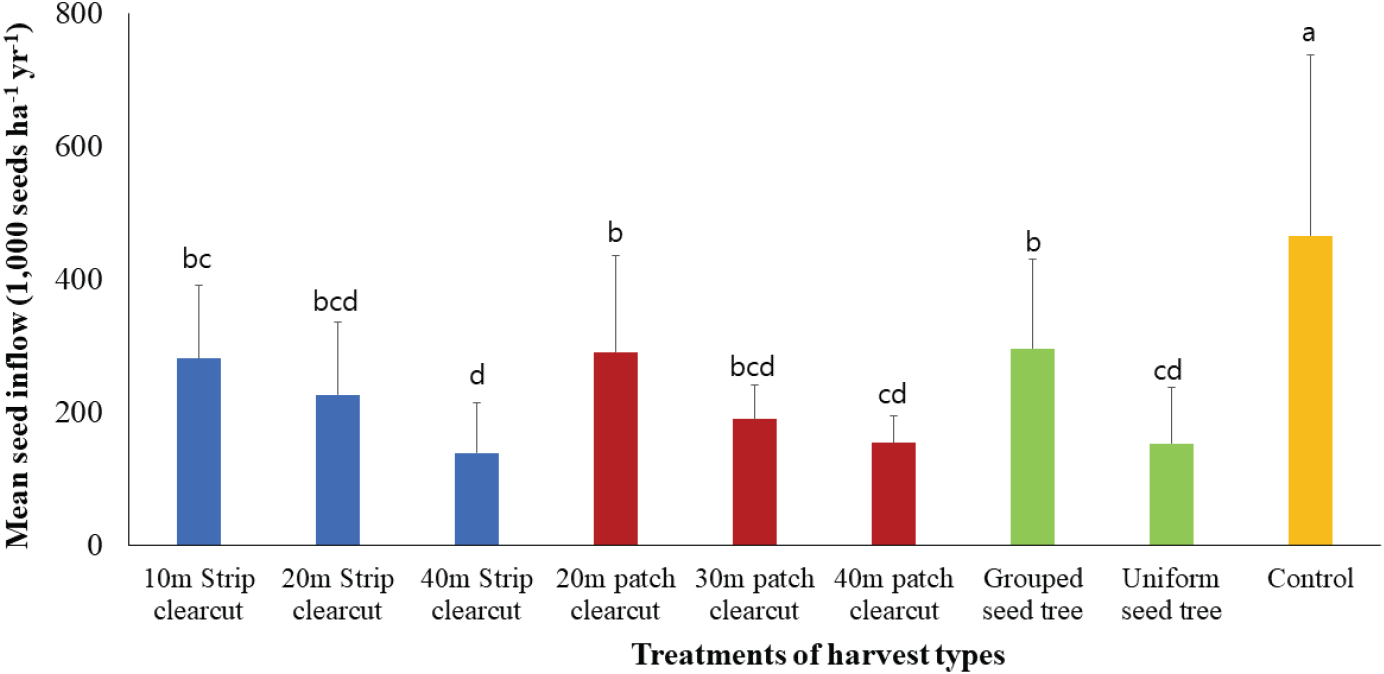
연평균 종자 유입량은 무처리구가 466 천립/ha/yr로 가장 많았고, 천연갱신 처리구에서는 군상 모수작업지 296 천립/ha/yr으로 가장 많고 40 m 대상개벌작업지가 136 천립/ha/yr으로 가장 적었다(Figure 8). 처리간에는 전체적으로 통계적 유의성을 보였지만, 군상 모수작업지와 20 m 군상 개벌작업지, 20 m 대상작업지와 30 m 군상작업지, 그리고 단목 모수작업지와 40 m 군상 개벌작업지에서는 통계적 차이를 보이지 않았다. 각각의 벌채 유형별 처리간에는 모두 통계적 유의성이 나타났으며 대상 개벌작업지와 군상 개벌작업지는 벌채 규모가 커질수록 평균 종자 유입량이 감소하는 경향을 보였다. 모수작업지는 군상 배치가 단목 배치에 비해 93.5 % 종자 유입량이 많은 것으로 나타났는데 이것은 갱신상의 형태와 면적에 따라 인접 수림대에 분포하는 종자공급원인 모수의 본수 차이가 원인인 것으로 판단된다. 그리고 본 연구지의 처리별 연평균 종자 유입량은 Lee et al.(2018)이 70년 된 소나무림의 연간 종자 생산량으로 보고한 250~270 천립/ha/yr과 유사하였다.
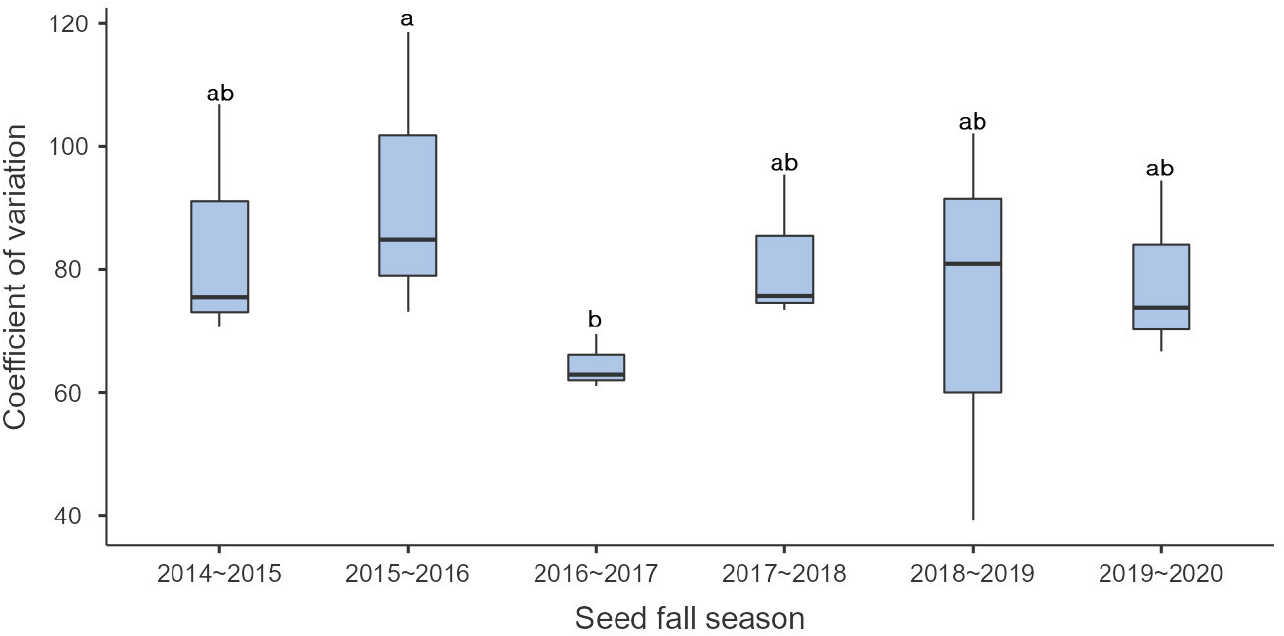
연도별 소나무 유입 종자의 분포 균일성과 종자 유입량을 비교·분석한 결과, 연도별 유입 종자의 분포 균일성은 통계적 유의성이 나타났고 종자 결실이 가장 좋았던 ’16~’17년도의 변이계수는 64.5로 가장 낮아 유입 종자가 갱신상에 균일하게 분포하고, 결실이 저조할 때(’14~’15년도 및 ’18~‘19년도)에는 갱신상에 유입된 종자의 분포가 균일하지 못하고 위치에 따른 변이도 커지는 것으로 나타났다(Figure 9). 이러한 경향은 Chung et al.(2021)과 Masahiko et al.(2016)가 일본잎갈나무 천연갱신지에서 종자 풍흉에 따른 유입 종자의 분포 균일성을 모니터링한 연구 결과와 동일하였다.
목재수확 유형별 유입 종자의 분포 균일성은 벌채작업을 하지 않은 무처리구의 유입 종자 분포가 변이계수 35.3으로 가장 균일한 것으로 분석되었다. 작업구 중에서는 군상 개벌 작업의 변이계수가 67.4로 종자 분포 균일성이 가장 좋았고, 수확지 내의 종자 공급을 위해 우수한 형질의 모수를 잔존시키지만 기본적으로 벌채 면적이 넓었던 모수 작업의 변이 계수가 가장 높아 종자 분포 균일성이 상대적으로 저조하였으며 각 유형 간에는 통계적 유의성을 보였다(Figure 10).
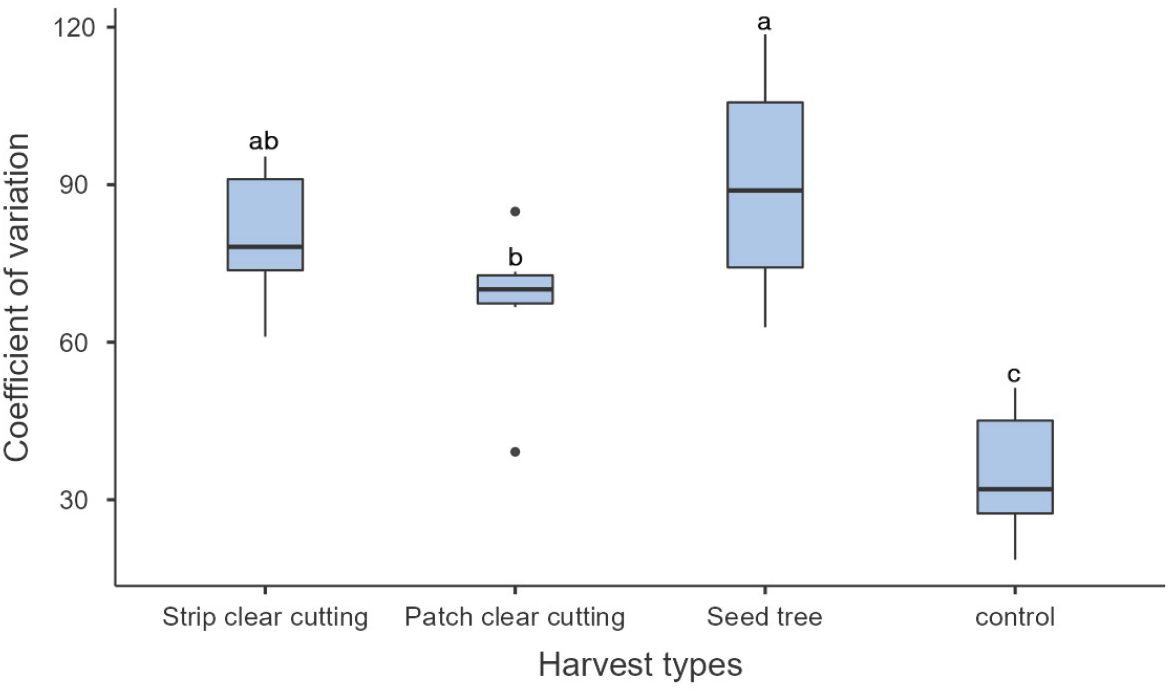
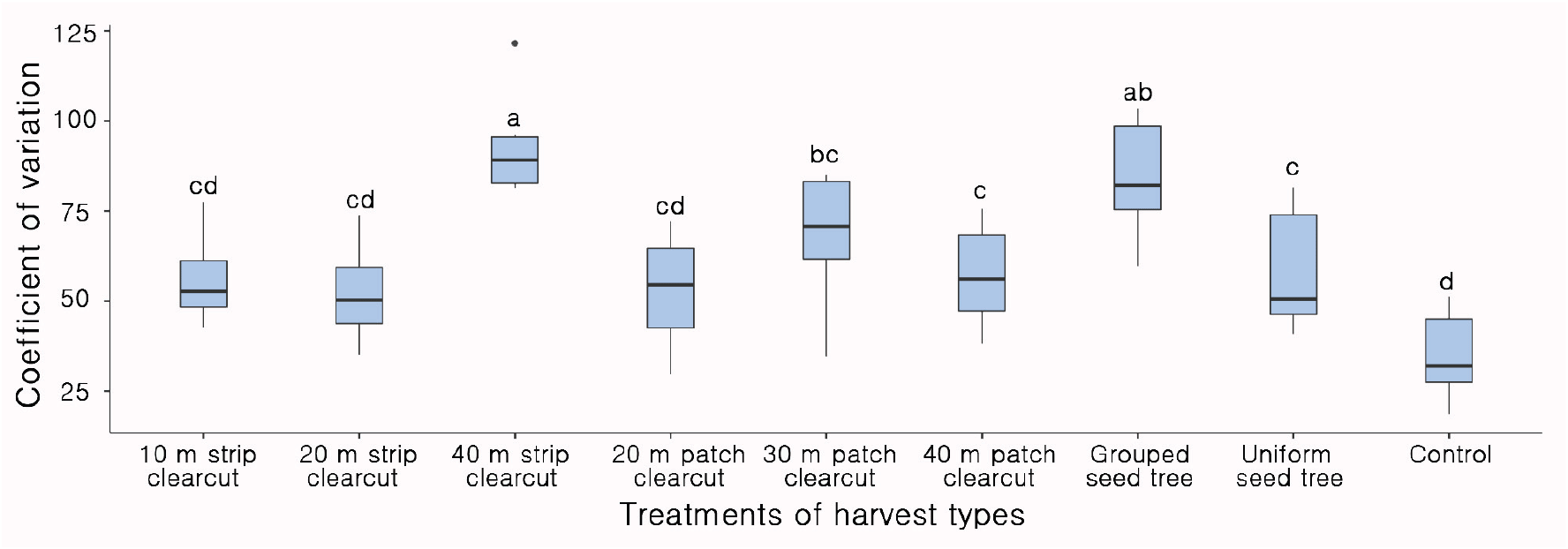
목재수확 유형의 처리별 유입 종자의 분포 균일성은 무처리구가 가장 좋았고 처리구에서는 20 m 군상 개벌작업지(52.2), 20 m 대상 개벌작업지(52.9) 그리고 10 m 대상 개벌작업지(56.1)의 변이계수가 낮았고 통계적 유의성이 없었다(Figure 11). 40 m 대상 개벌작업지(93.2)와 군상 모수작업지(84.1)는 변이 계수가 높아 종자의 분포 균일도가 가장 저조하였다(Figure 11). 본 연구에서는 벌채 폭이나 벌채 직경이 20 m까지는 종자 분포 균일성이 유사하다가 벌채 크기가 증가할수록 균일성이 낮아져, 소나무 모수림인 임연부로부터 멀어질수록 종자 낙하량은 급격하게 감소하여 모수의 수고 이상에서는 종자가 발견되지 않았고(Korea Forest Research Institute, 1985) 모수로부터 10~15 m 범위에는 충분한 양의 종자가 도달한다는 연구결과와 유사하였다(Kim et al., 2021). 따라서 벌채 폭이 좁을수록 모수와의 거리가 가깝고 모수의 밀도가 높아져 벌채지의 평균 종자 밀도는 증가하지만 벌채 폭의 확대화 함께 감소하는데(Oya et al., 2020) 소나무와 같이 바람을 매개로 하는 종자의 비산 거리는 종자공급원의 높이와 바람 조건의 영향을 가장 크게 받기에(Nathan, 2011) 천연갱신에 활용하기 위해서는 다양한 조건에서의 종자 유입 연구와 종자 공급 이후의 숲의 발달조건을 고려한 추가적인 연구가 필요할 것으로 사료된다.
결 론
본 연구는 목재수확 유형에 따른 종자 유입 특성을 구명하여 천연갱신을 통한 소나무 후계림 조성 기술 개발의 기초자료를 제공하기 위해서 수행되었다. 소나무의 종자 비산 시기는 10월에서 이듬해 2월까지 이루어졌으며, 10~ 11월에 전체 유입 종자의 80 % 이상이 유입되는 것으로 나타나 갱신상은 천연갱신 대상지의 종자 성숙 및 비산시기를 미리 파악하여 조성이 되어야 할 것으로 판단되었다. 목재수확 유형별 종자 유입량과 유입된 종자의 분포 균일성은 벌채 면적이 좁은 10 m 대상 개벌작업지와 20 m 군상 개벌작업지에서 양호하였다. 모수를 많이 남기는 목재수확 방식이 천연갱신에 유리할 것으로 판단 할 수 있으나, 성공적인 천연갱신 후계림 조성을 위해서는 종자유입 이후 발아, 천연갱신 치수 발생⋅생존 및 생장을 고려한 종합적인 검토 과정을 통해 적절한 목재수확 방식을 결정할 필요가 있다. 따라서, 임목 수확에 따른 수광량 변화, 임분내 풍환경, 수림대 수관 발달 등 천연갱신지의 생육환경 변화에 관한 다양한 정보 수집 및 연구가 지속적으로 수행되어야 할 것이다.