



서 론
천연기념물은 자연의 역사와 가치가 내포된 문화유산이자 자연유산으로 특히 진귀성과 희귀성, 고유성과 특수성, 분포성과 역사성을 특징으로 한다(Son et al., 2015). 우리나라 천연기념물은 문화재보호법에 따라 문화재의 보존관리 및 활용으로 원형 유지를 기본 원칙으로 하고 있다(Kim et al., 2014). 보존(preservation)은 원상태의 고유한 생태계를 유지하기 위해 이용은 물론 최대한 인위적 관리를 하지 않는 것을 말하지만(Kwon, 2005), 문화재 정책에서는 문화재 보존을 위하여 문화재의 적절한 인위적 간섭 또는 활용이 필요한 것으로 보고 있다(Lee, 2015).
우리나라의 천연기념물은 크게 동물, 식물, 지질⋅지형 천연보호구역 등으로 나뉘고 식물은 노거수, 수림지, 마을숲, 희귀종, 자생지, 분포 한계지로 구분된다(Son et al., 2016). 수림지는 그 자체로 식생이 유지되는 숲으로 특수한 지역에 발달하는 식물의 군락이나 희귀한 종류, 특수한 습성을 가진 식물군락을 보존하고자 지정하여 관리하는 숲이다(CHA, 2003). 수림지는 산업화와 도시의 무분별한 확대로 생육환경이 악화되고 있으며(Hwang, 2022) 시간의 흐름에 따른 식생의 변화, 자연재해 등으로 인한 대규모 훼손, 생육환경 변화에 따른 식생의 쇠퇴 및 변화 등 다양한 변화가 일어날 수 있어 현재의 실태를 정확히 파악하는 것은 매우 중요하다(Kim et al., 2011). 이러한 산림 군락의 발달 과정과 보호, 관리 등을 위해서는 종의 구성, 군락구조와 같은 군집생태학적 연구가 필요하다(Lee et al., 2015; Kim et al., 2017).
한편 다양한 기후대가 분포하고 독특한 식생대가 형성된 제주도는 한라산을 기준으로 북쪽에 제주시, 남쪽에 서귀포시로 구분되며(Kim, 1995; Han et al., 2013) 천연기념물 수림지는 제주시 2곳(제주 평대리 비자나무 숲, 제주 납읍리 난대림), 서귀포시 5곳(제주 물장오리오름, 제주 안덕계곡 상록수림, 제주 천지연 난대림, 제주 천제연 난대림, 제주 산방산 암벽식물지대)이 지정되었다. 이중 제주시에 지정된 제주 평대리 비자나무 숲(이하 ‘비자나무 숲’이라 한다)과 제주 납읍리 난대림(이하 ‘납읍리 난대림’이라 한다)은 화산활동 중 분출한 용암류가 만들어낸 불규칙한 암괴지대인 곶자왈 지역에 포함되어 있어 독특한 생태계가 유지되고 있는 곳이다(Kim et al., 2020). 비자나무 숲은 우리나라 비자나무 숲 중 오래된 수령을 가진 다수의 노거수를 포함한 비자나무림이며 납읍리 난대림은 제주시 서부 평지에 남아있는 유일한 상록수림으로 1993년 천연기념물로 지정되었다(Jeju Gotjawal Provincial Park, 2011; CHA, 2022). 현재 비자나무 숲과 납읍리 난대림은 산책로 조성으로 관광객 및 인근 마을 주민들이 방문하고 있으며 활용적 측면에서 인위적인 관리가 이루어져 생육환경의 변화가 발생되고 있다. 두 수림지의 식생연구는 비자나무림의 식생 구조 및 관리 방안(Lee and Oh, 2013), 보존 관리 방향 설정을 위한 기초연구(Lee et al., 2014), 식물생태학적 가치 제고(Choi and Lee, 2015)가 있으며 납읍리 난대림의 식물상 및 식생(Boo, 2006)에 관한 연구가 진행되었지만 군집생태학적 연구는 미흡한 실정이었다. 식생은 생육지의 환경과 반응하여 결정된 종들이 모인 집단이 군락의 형태로 존재하고 있어 예로부터 식생들의 유형화를 통해 대상 지역의 산림생태계 보전 및 관리의 기본 단위로 활용되고 있다(Lee et al., 2006). 따라서 본 연구는 제주도 제주시에 있는 제주 평대리 비자나무 숲과 제주 납읍리 난대림의 식생변화에 대응하는 체계적인 관리방안을 마련하기 위해 식물사회학적 방법에 따른 식생 유형을 밝히고 각 식생 유형의 군집구조에 대한 특징을 구명하고자 한다.
재료 및 방법
비자나무 숲은 제주시 구좌읍 평대리에 있으며, 다랑쉬오름(382.4 m)에서 시작하여 둔지봉(282.2 m)에서 끝나는 구좌-성산 곶자왈의 중심에 있으며 해발 125~160 m에 이르는 완만한 경사의 평지대에 위치한다(Choi and Lee, 2015). 면적은 448,753 m²로 비자나무가 우점하는 순림으로 보전 가치가 매우 높다(Shin, 2012).
납읍리 난대림은 북제주군 애월읍에 있으며, 노꼬메오름에서 시작된 애월읍 곶자왈의 끝자락 해발 80~100 m 지역에 위치한다(Boo, 2006). 면적은 33,980 m²로 제주 북서부에 유일하게 남아있는 상록활엽수림이다(Figure 1).
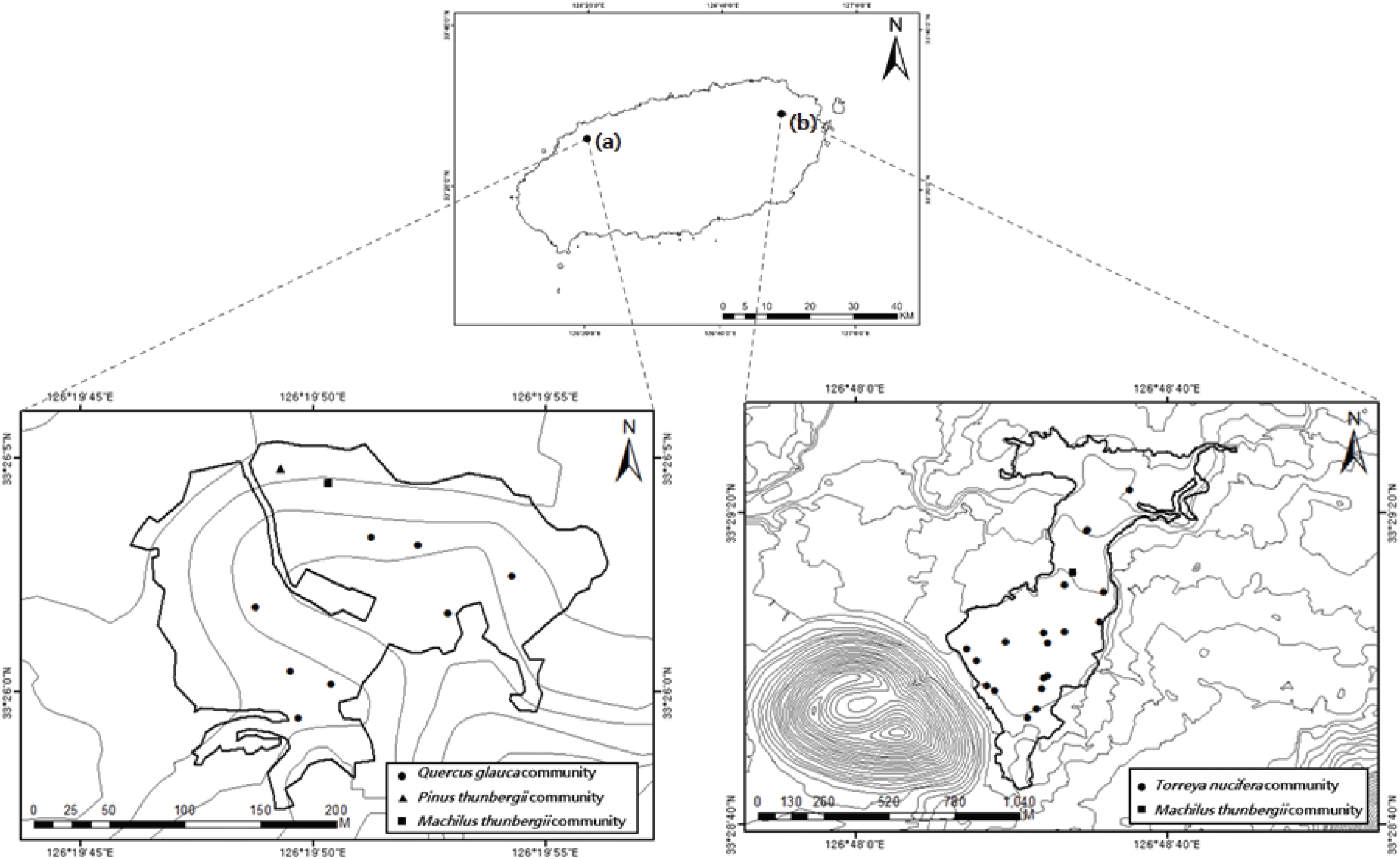
식생 조사는 비자나무 숲과 납읍리 난대림을 대상으로 2021년 5월부터 6월까지 Z-M학파의 식물사회학적 방법(Ellenberg, 1956; Braun-Blanquet, 1964)에 따라 비자나무 숲 20개소, 납읍리 난대림 10개소로 총 30개소의 식생 조사자료를 수집했다. 출현하는 모든 종을 대상으로 교목층, 아교목층, 관목층, 초본층으로 나누어 각 층에 출현하는 종에 대한 우점도, 평균수고, 평균흉고직경, 우점종 등을 기록하였고, 환경인자로는 해발고도, 방위, 경사, 암석 노출도, 토양 노출도, 낙엽층의 깊이 등을 측정하였다. 종 동정은 원색식물도감(Lee, 2003)과 나무생태도감(Yun, 2021)을 기준으로 하였으며, 명명은 국가생물종지식정보시스템(KFS, 2022a)과 국가표준식물목록(KFS, 2022b)을 기준으로 작성하였다.
자료 분석은 식생의 군락 유형을 분류하기 위해 PC-ORD v7.03 프로그램을 이용한 계층적 군화분석 방법인 Hill(1979)의 TWINSPAN(Two-Way Indicator Species Analysis)으로 Ellenberg(1956) 의 여러 표조작 과정을 함축 분석한 후 최종적으로 상재도로 나타낸 식별표를 작성하여 산림식생유형분류를 하였다. 출현하는 모든 종의 상대적 점유 정도를 파악하기 위해 Curtis and McIntosh(1951)방법을 이용하여 상대빈도, 상대밀도를 합산한 중요치[(상대빈도)+(상대밀도)/2]를 층위별로 분석하였다. 종다양도는 각 군락유형에 대한 종의 다양성, 경쟁 등을 확인하기 위해 종다양도지수(Shannon and Weaver, 1949), 최대종다양도, 균재도, 우점도를 분석하여(Brower and Zar, 1977) 식생유형별로 나타냈다.
결과 및 고찰
비자나무 숲과 납읍리 난대림의 산림식생유형을 분류한 결과 1개의 군락군(Community group), 2개의 군락(Community), 4개의 군(Group)으로 총 4개의 식생유형으로 구분되었다(Table 1, Figure 2). 최상위 수준에서 까마귀쪽나무군락군(Litsea japonica community group)으로 분류되었고, 까마귀쪽나무군락군은 비자나무군락(Torreya nucifera community)과 종가시나무군락(Quercus glauca community)으로 분류되었다. 비자나무군락은 이나무군(Idesia polycarpa group)과 홍지네고사리군(Dryopteris erythrosora group)으로 분류되었고 종가시나무군락은 산쪽풀군(Mercurialis leiocarpa group)과 가는쇠고사리군(Arachniodes aristata group)으로 각각 분류되었으며 종군은 대별 종군을 포함하여 총 8개의 종군으로 구분되었다. 식생유형 1(까마귀쪽나무군락군-비자나무군락-이나무군)과 2(까마귀쪽나무군락군-비자나무군락-홍지네고사리군)는 비자나무 숲에 분포하였으며 식생유형 3(까마귀쪽나무군락군-종가시나무군락-산쪽풀군)과 4(까마귀쪽나무군락군-종가시나무군락-가는쇠고사리군) 종가시나무는 납읍리 난대림에 분포하고 있었다.

본 군락군단위의 표징종은 까마귀쪽나무, 마삭줄, 콩짜개덩굴, 송악, 남오미자, 후박나무 등이 나타났다. 까마귀쪽나무는 난·온대 상록활엽수로 벌채와 경작, 개발 등으로 자생지가 파괴 또는 파편화되어 소수 군락으로 자생하고 있지만 기후변화로 그 자생지가 확장될 것으로 예측되는 수종이다(Oh, et al., 2010; Yu, et al., 2020). 또한 마삭줄, 콩짜개덩굴, 송악, 남오미자 등 덩굴식물은 목본성식물의 생육을 방해할 뿐 아니라 임목의 수형에도 악영향을 줄 수 있으므로(Byeon and Yun, 2018) 덩굴식물이 피압할 경우 수림지 보호를 위해 관리가 필요할 것으로 판단되었다. 상재도가 Ⅲ 이상인 종은 까마귀쪽나무, 마삭줄, 콩짜개덩굴, 송악, 남오미자, 후박나무, 거지덩굴, 줄사초, 담쟁이덩굴, 계요등이 나타났으며, 우점도가 3 이상인 종은 마삭줄, 줄사초, 담쟁이덩굴이 나타났다.
본 군락군단위의 표징종은 비자나무, 상산, 십자고사리, 큰천남성 등이 나타났다. 비자나무 숲의 식생구조에 관한 연구(Lee and Oh, 2013)에 의하면 비자나무-상산군락으로 구분되었으며 십자고사리와 큰천남성 등이 출현하여 본 연구와 유사한 종구성이 나타났다. 비자나무속은 세계적으로 5종이 분포하고 우리나라에는 비자나무 단 1종이 자생하고 있으며(KNA, 2015) IUCN 적색목록에 약관심종(Least Concern, LC)으로 광범위하고 풍부한 분류군에 해당(KNA, 2021)되지만 향후 환경훼손, 교란으로 위협을 받을 가능성이 크므로(Oh et al., 2015) 지속적인 모니터링이 필요할 것으로 판단되었다.
본 식생유형은 까마귀쪽나무군락군에서 종군 2의 비자나무, 상산 등 식별종 출현으로 비자나무군락으로 구분되어졌으며 비자나무군락의 하위 종군으로 이나무, 새비나무, 때죽나무, 찔레꽃 등의 출현에 의해 이나무군으로 구분됐으며 상재도가 Ⅲ 이상인 종은 까마귀쪽나무, 마삭줄, 콩짜개덩굴, 송악, 남오미자, 후박나무, 거지덩굴, 줄사초, 담쟁이덩굴, 계요등, 비자나무, 상산, 십자고사리, 큰천남성, 백량금, 나도생강, 자금우, 단풍나무, 새우난초류, 이나무, 새비나무, 때죽나무가 나타났으며, 우점도가 3 이상인 종은 마삭줄, 줄사초, 비자나무, 상산, 나도생강, 자금우, 단풍나무, 이나무로 나타났다. 우리나라 비자나무림 전 지역에 관한 선행 연구(Shin et al., 2010)에 의하면 비자나무와 지위를 같이 하는 종은 찔레꽃, 송악, 마삭줄로 본 식생유형 출현 종과 일치하는 것으로 나타났다.
본 식생유형은 비자나무군락의 하위 종군으로 종군 5의 홍지네고사리, 바위수국, 머귀나무, 예덕나무 등의 출현에 의해 홍지네고사리군으로 구분되어 졌다. 상재도가 Ⅲ 이상인 종은 까마귀쪽나무, 마삭줄, 콩짜개덩굴, 송악, 남오미자, 푸조나무, 거지덩굴, 줄사초, 쇠고비, 담쟁이덩굴, 천선과나무, 비자나무, 상산, 십자고사리, 큰천남성, 백량금, 나도생강, 자금우, 왜모시풀, 단풍나무, 천남성, 박쥐나무, 비목나무, 왜승마, 홍지네고사리, 바위수국이 나타났으며, 우점도가 3 이상인 종은 콩짜개덩굴, 송악, 후박나무, 거지덩굴, 줄사초, 담쟁이덩굴, 비자나무, 상산, 십자고사리, 나도생강, 자금우, 단풍나무, 홍지네고사리, 긴사상자로 나타났다.
본 군락군단위의 표징종은 종가시나무, 후추등, 도깨비쇠고비 등이 나타났다.
본 식생유형은 까마귀쪽나무군락군에서 종군 3의 종가시나무, 후추등, 도깨비쇠고비 등 식별종 출현으로 종가시나무군락으로 구분됐으며 종가시나무군락의 하위 종군으로 산쪽풀과 산유자나무의 출현에 의해 산쪽풀군으로 구분되어 졌다. 산쪽풀속 중 우리나라에는 산쪽풀 1종만이 분포하고 있으며 IUCN 적색목록 약관심종(Least Concern, LC)으로 관리가 필요한 종이다(KNA, 2021). 상재도가 Ⅲ 이상인 종은 까마귀쪽나무, 마삭줄, 콩짜개덩굴, 송악, 남오미자, 후박나무, 푸조나무, 거지덩굴, 담쟁이덩굴, 계요등, 쇠고비, 종가시나무, 후추등, 도깨비고비, 보리밥나무, 산호수, 윤판나물, 산쪽풀, 담팔수로 나타났으며, 우점도가 3 이상인 종은 까마귀쪽나무, 마삭줄, 남오미자, 후박나무, 종가시나무, 후추등, 산호수, 산쪽풀로 나타났다.
본 식생유형은 종가시나무군락의 하위 종군으로 종군 7의 가는고사리, 굴거리나무 등의 출현에 의해 가는쇠고사리군으로 구분되어 졌다. 상재도가 3 이상인 종은 까마귀쪽나무, 마삭줄, 콩짜개덩굴, 송악, 남오미자, 후박나무, 푸조나무, 종가시나무, 후추등, 보리밥나무, 산호수로 나타났으며, 우점도가 3 이상인 종은 까마귀쪽나무, 마삭줄, 후박나무, 종가시나무, 후추등으로 나타났다. 본 식생유형에서는 귀화식물인 미국자리공이 나타났다. 귀화식물은 안정된 군락에 쉽게 침입, 정착할 수 없으므로(Yang and Kim, 2005) 납읍리 난대림의 생태계가 교란을 받고 있는 것으로 판단되었다.
식생유형의 교목층(T), 아교목층(ST), 관목층(S), 초본층(H)의 중요치 산출을 통한 각 식생유형 구성종의 상대적인 중요도를 파악하였다(Table 2).
식생유형 1은 교목층(T)에서 비자나무(39.5), 후박나무(13.1), 이나무(12.7), 곰의말채나무(8.5)등으로, 아교목층(ST) 단풍나무(18.9), 아왜나무(12.4), 송악(10.1), 비자나무(9.9)등으로, 관목층(S)은 상산(40.0), 새비나무(7.3), 단풍나무(5.7), 비자나무(4.5)등으로, 초본층(H)은 마삭줄(9.8), 아왜나무(4.5), 콩짜개덩굴(3.0), 비자나무(2.3)등으로 나타났다. 식생유형 2는 교목층에서 비자나무(57.5), 후박나무(8.1), 팽나무(6.9), 머귀나무(6.9)등으로, 아교목층 단풍나무(22.0), 비자나무(10.7), 비목나무(7.3), 아왜나무(5.9)등으로, 관목층 상산(22.6), 까마귀쪽나무(9.2), 아왜나무(8.8), 콩짜개덩굴(7.2)등으로, 초본층 송악(5.0), 콩짜개덩굴(4.0), 아왜나무(3.3), 생달나무(2.2)등으로 나타났다.
비자나무 숲(식생유형 1과 2)은 교목층에 비자나무, 아교목층에 단풍나무, 관목층에 상산이 우점하는 구조를 보이고 있다. 이들 종은 비자나무림의 기원에 관한 근거를 반영하는 진단종으로 판단되고 있다(Choi and Lee, 2015). 교목층의 경우 비자나무의 중요치가 높아 당분간은 비자나무가 상층을 이루고 있을 것으로 판단되었으나 초본층에서 비자나무의 중요치가 낮고 마삭줄, 콩짜개덩굴 등 덩굴성 수종의 중요치가 높게 나타났다. 전남 화순 개천산 비자나무림 선행연구에 의하면 칡, 송악, 마삭줄, 담쟁이덩굴 등 덩굴식물에 의해 비자나무림의 피해가 발생한다고 하였다(Park, 2007).
식생유형 3은 교목층에서 종가시나무(51.9), 후박나무(30.1), 팽나무(11.3) 등의 순으로 나타났다. 난대림은 종가시나무, 후박나무, 팽나무가 조사지 전체에 분포 하고 있다는 연구(Boo, 2006)와 일치하는 것으로 나타났다. 아교목층 종가시나무(27.0), 후추등(15.8), 까마귀쪽나무(14.5), 콩짜개덩굴(14.2) 등으로, 관목층 종가시나무 (24.9), 까마귀쪽나무(19.5), 후추등(15.4), 후박나무(9.0)등으로, 초본층 산쪽풀(15.0), 마삭줄(7.8), 후추등(7.6), 남오미자(6.6)등으로 나타났다. 식생유형 4는 교목층에서 종가시나무 (44.8), 후박나무(30.5), 곰솔(24.7)로 나타났으며 모든 층위에서 고루 분포하고 있는 종가시나무와 후박나무가 유지될 것으로 보이나 양수인 곰솔의 경우 쇠퇴될 것으로 판단되었다. 아교목층 종가사지나무(33.6), 콩짜개덩굴(19.7), 후박나무(14.4), 굴거리나무(9.1)등으로, 관목층 종가시나무(19.1), 까마귀쪽나무(16.8), 후박나무(9.8), 푸조나무(6.3)등으로, 초본층 후추등(14.8), 마삭줄(9.7), 송악(7.4), 큰족제비고사리(5.8)등으로 나타났다. 우리나라 상록활엽수림은 도서 지역의 인구 증가와 연료림 채취로 인해 대부분 파괴되었으며 지리적으로 격리된 섬에 소규모로 남아있어 난대생태계 보존 및 복원대책이 필요한 상황이다(Oh and Kim, 1996). 납읍리 난대림(식생유형3과 4)은 종가시나무, 후박나무, 까마귀쪽나무, 굴거리나무 등이 출현하여 상록활엽수림의 모습을 보이고 있지만 아교목층까지 올라온 콩짜개덩굴, 송악, 후추등, 마삭줄 등의 덩굴성 수종이 햇빛을 차단하여 정상적인 생육에 영향을 미치므로(Hyum et al., 2014) 덩굴성 식물의 관리가 필요할 것으로 판단되었다.
군락의 종다양도와 성숙도는 일반적으로 정비례하는 경향을 보이는데(Loucks, 1970) Shanon의 종다양도지수를 분석한 결과 식생유형 1은 2.866, 식생유형 2는 2.716로, 식생유형 3는 2.222, 식생유형 4는 2.326로 나타났다(Table 3). 제주도기념물 제10호로 지정된 동백동산의 종다양도는 1.0268~1.2102(Kwak et al., 2013)로 비자나무 숲과 납읍리 난대림이 성숙한 군락으로 판단되었다. 식생유형별 종다양도의 경우 비자나무 숲(식생유형 1, 2)이 납읍리 난대림(식생유형 3, 4) 보다 종다양도가 높은 것으로 나타났다. 종다양도는 이질적이거나 복잡한 생육환경에 국소적인 교란이 발생할 때에 높게 나타나는데(Krebs, 1985; Barbour et al., 1987) 비자나무 숲은 수관을 우점하고 있는 노거수를 다양한 시술(Lee et al., 2014), 수관층의 과도한 택벌로 인한 광환경 및 수분환경 변화 또는 수목 생장에 양호한 환경조건이 이루어져 다양한 종이 출현했기 때문인 것으로 판단되었다(Choi and Lee, 2015). 납읍리 난대림은 종가시나무와 같은 상록활엽수가 우점한 군락으로 수관 하부에 내음성이 강한 소수의 종이 제한적으로 나타나는 특성을 보여 종 다양성이 낮게 나타난 것으로 판단되었다(Park 2012, Kim et al., 2020).
| Vegetation unit | H’ | Hmax’ | J’ | 1-J’ |
|---|---|---|---|---|
| VU1 | 2.866 | 3.615 | 0.792 | 0.208 |
| VU2 | 2.716 | 3.505 | 0.775 | 0.225 |
| VU3 | 2.222 | 3.164 | 0.703 | 0.297 |
| VU4 | 2.326 | 3.234 | 0.719 | 0.281 |
결 론
본 연구는 천연기념물 수림지인 제주 평대리 비자나무 숲과 제주 납읍리 난대림의 군집구조를 파악하고자 수행하였다. 식물사회학적 식생유형분류 결과 최상위 수준에서 까마귀쪽나무군락군으로 분류되었다. 식생유형 1과 2는 비자나무 숲, 식생유형 3과 4는 납읍리 난대림에 분포하고 있으며 식생유형별 종조성의 차이를 보이는 것으로 나타나 식생유형별 차별화된 관리 방안의 마련이 필요할 것으로 판단되었다. 비자나무 숲의 층위구조는 교목층에 비자나무, 아교목층에 단풍나무, 관목층에 상산이 우점하는 구조를 보이고 있었으며, 납읍리 난대림의 경우 종가시나무가 우점하는 군락에 후박나무, 곰솔, 팽나무 등과 함께 군락을 형성한 것으로 판단되었다. 군락군 수준에서 마삭줄, 공짜개덩굴, 송악, 담쟁이덩굴 등 덩굴식물의 중요치가 높아 주요 구성종인 비자나무, 종가시나무 개체군 구조에 미치는 영향이 높으므로 장단기적으로 식생구조를 고려한 생태학적 관리가 필요할 것으로 판단되었다.
결론적으로 천연기념물 수림지는 인간의 간섭을 지속적으로 받아 왔고 또 앞으로도 간섭을 받게 될 것으로 예상되는 바, 식생단위별 종조성을 중심으로 수림지 보존을 위한 식생학적 관리 방안의 체계화 그리고 장기적인 식생변화 모니터링이 필요할 것으로 판단되었다.