



서 론
기후변화(climate change)로 인한 급격한 변화와 자연생태계의 교란은 국제적인 현안이다. 기온이 상승하며 식물의 개화 시기가 빨라지고, 동식물의 서식지가 교란되는 등 생태적 부작용이 발생한다(Kong, 2005). 특히 고산대와 아고산대에 고립되어 분포하는 종, 생육 범위가 좁거나 이동 속도가 느린 종 등은 기온 상승에 따른 생리적 스트레스의 가중으로 소멸 위험성이 높아질 것으로 예측된다(Smiley, 1968; Kong, 1998; Horikawa et al., 2009; Stokew and Yun et al., 2010).
분비나무(Abies nephrolepis)는 주로 해발고도 700-2,540 m에 생육하는 아한대의 표징종으로서 IUCN 약관심종(LC)으로 등재되어 있으며 세계적으로 한국, 만주, 시베리아에 분포한다(Zhang et al., 2013). 아고산대는 바람이 강하고, 기온은 낮으며 적설량도 불규칙하는 등 여러 기상요소의 편차가 심하여 식물이 살아가기에 척박한 환경이다(Park et al., 2015). 이러한 아고산지대에 생육하는 분비나무는 기후변화가 가속화되는 시점에 생육과 존속에 어려움을 직면하여 쇠퇴 및 집단고사 현상이 관찰되고 있다(Chun et al., 2019; Kim et al., 2021; Park et al., 2022).
분비나무와 관련된 연구로는 기후변화 민감성 평가에 관한 연구(Park et al., 2015; Lee et al., 2023), 식물의 군집구조 및 동태에 관한 연구(Chun et al., 2019; Park et al., 2020; Lee et al., 2021; Park et al., 2021; Park et al., 2022; Cheon et al., 2022; Shin et al., 2022) 등이 있다.
교란(disturbance)이란 생태계, 군집, 개체군 구조를 방해하고 가용자원 또는 물리적인 환경을 변화시키는 상대적으로 불연속적인 사건이며(Pickett and White, 1985; Duguid et al., 2013), 기후변화와 함께 연속적인 관계에 놓여 산림 내에 다양한 규모의 숲틈을 발생시킨다(Bergeron et al., 1998). 최근 덩굴성 식물종인 미역줄나무가 교란을 통해 분비나무 등 아고산 침엽수에 고사 및 쇠퇴에 영향을 끼친다는 연구가 보고되고 있다(Kim et al., 2020; Cheon et al., 2022). 덩굴의 확산 및 밀도의 증가는 수종의 발달에 부정적인 영향을 미쳐 교란 이후 숲의 재생 과정을 저해하는 것으로 여겨진다(Zahawi and Augspurger, 1999; Schnitzer et al., 2000). 미역줄나무는 교란 후 숲 틈이 발생한 환경에서 급격히 확장하는 등 지리산, 설악산, 계방산 등의 상록침엽수림에서 치수갱신과 숲 구조에서 부정적인 영향이 나타나고 있다(Kim et al., 2018; Kim et al., 2020). 또한 분비나무, 구상나무의 생육발달에 영향을 줄 것으로 예측되며, 아고산 침엽수림에서 중점 관찰이 필요한 수종으로 판단된다(Park et al., 2016).
그러나 덩굴성 식물인 미역줄나무가 분비나무림의 식생구조에 어떠한 영향을 끼치는지에 대한 세부적인 연구는 부족한 실정이다. 본 연구는 미역줄나무 출현에 따른 분비나무의 지속성에 대해 식물사회학적 방법에 따라 분석하고자 하였다. 이러한 자료를 통해 분비나무의 보전 및 관리에 기여하고자 한다.
재료 및 방법
본 연구는 분비나무 자생지인 가리왕산-중왕산, 두타산-상원산, 명지산, 발왕산, 방태산-개인산, 백덕산, 백운산-함백산-장산, 사달산, 설악산, 소백산, 오대산-계방산, 점봉산-망대암산, 청옥산-두타산, 화악산-석룡산, 황병산, 흰봉산의 16개 산지 내, 분비나무가 상층 수관을 점유하는 140개소를 대상으로 분석하였다(Figure 1).
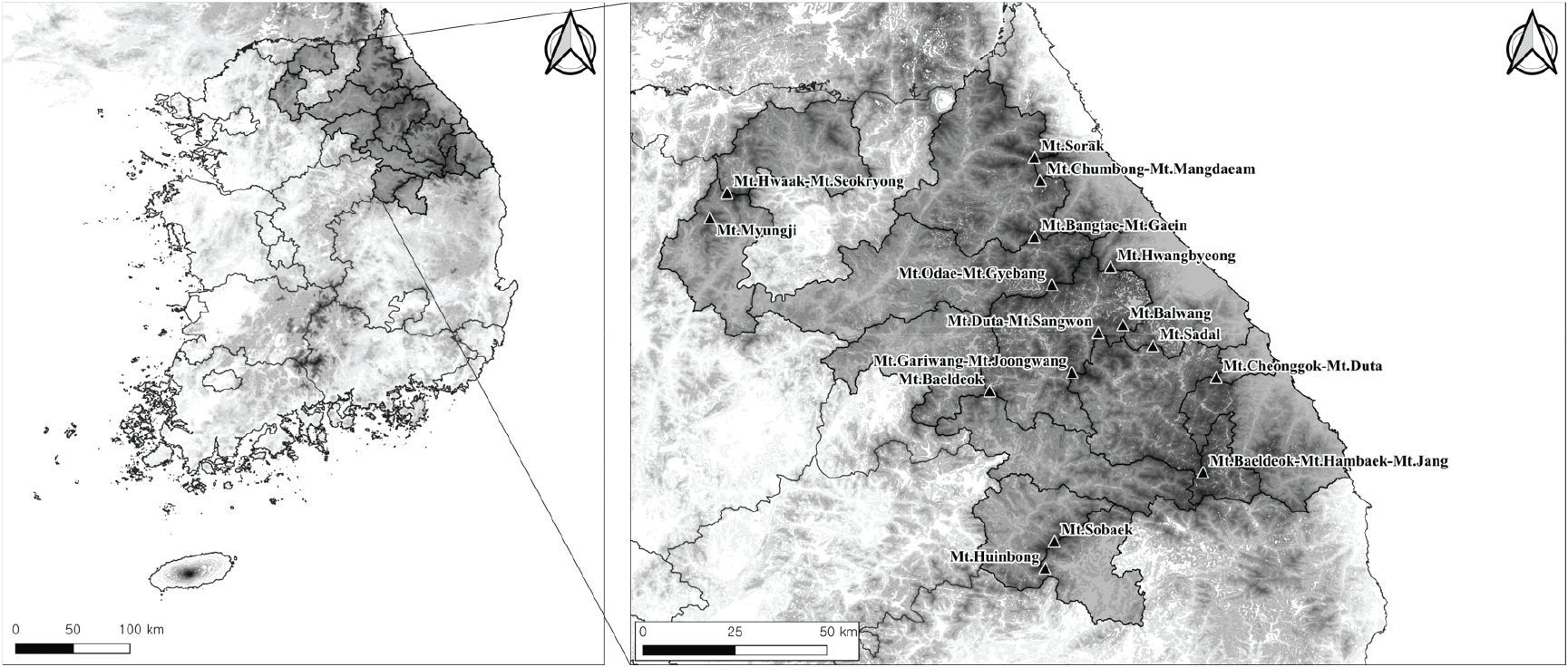
행정구역상 조사 대상지는 강원도 화천군, 인제군, 홍천군, 양양군, 강릉시, 평창군, 횡성군, 정선군, 동해시, 삼척시, 영월군, 태백시와 경기도 가평군, 충청북도 단양군, 경상북도 영주시에 속해있다. 조사지와 근접한 기상관측소인 강릉, 대관령, 북강릉, 삼척, 영월, 인제, 정선, 태백, 홍천, 영주의 최근 10년간(2013-2022) 관측값을 살펴보면, 평균기온은 11.2°C, 평균 최고기온은 16.9°C, 평균 최저기온은 6.3°C, 연평균 강수량은 1,190.0mm으로 나타났다(Korea Meteorological Administration, 2023).
조사구는 반지름 11.3 m의 원형 방형구(약 400 m2)로 설정하였고, 매목조사는 방형구 내 흉고직경 6cm 이상의 수종을 대상으로 측정하였으며, 식생조사는 Z-M학파의 식물사회학적 방법(Braun-Blanquet, 1965)에 따라 조사하였다. 또한 암석노출도, 해발고도, 사면 경사도, 사면 방위각 등의 입지 환경 특성을 파악하고 분비나무 치수는 수고에 따라 0(10 cm 미만), 1(10 cm 이상 50 cm 미만), 2(50 cm 이상) 등급으로 구분하여 개체수를 조사하였다. 출현식물은 Engler의 분류체계(Melchior, 1964)의 기준에 따라 목록을 정리하였다. 구성종의 동정과 식물분류는 원색대한식물도감(Lee, 2003)을 참조하였으며, 종명의 국명과 학명은 국가표준식물목록(Korea National Arboretum, 2022) 기준을 적용하였다.
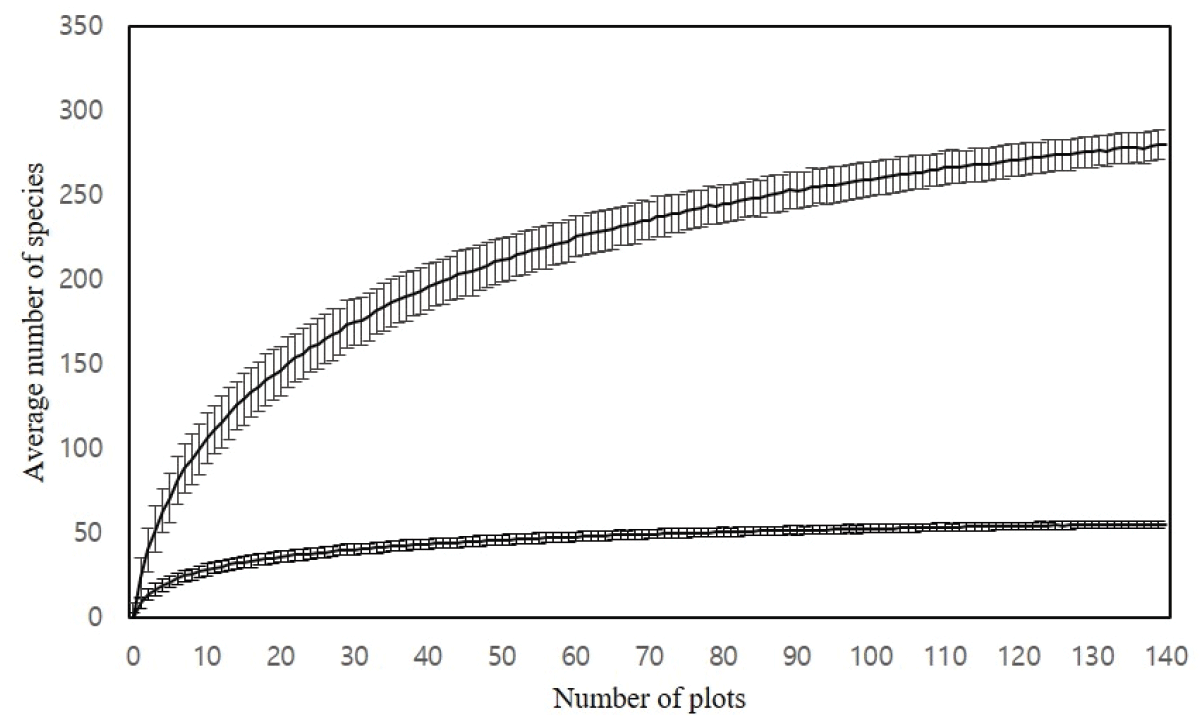
조사기간은 2021년 5월부터 2022년 10월까지 진행하였고, 야외에서 수집한 데이터는 MS-Excel 2019 프로그램을 이용하여 정리하였다. 본 연구에 앞서, 연구대상지 내 적절한 조사구 수를 만족하는지 판단하기 위해 종면적곡선을 분석하였다.
미역줄나무의 출현에 따른 식생구조 구조를 파악하기 위해 미역줄나무의 출현 유무와 미역줄나무가 출현하는 층위의 2가지 기준에 따라 3가지 유형으로 구분하였다. 미역줄나무가 교목·아교목층에 출현하는 유형(tree·subtree presence: TP), 관목층에 출현하는 유형(shrub presence: SP), 모든 층위에서 출현하지 않는 유형(absence: A)으로 분류되었다. 구분한 유형의 환경요소와 임목구조를 살펴보았다. 환경요소에서 암석노출도, 사면방위는 유형별 비율을 나타냈고, 해발고도, 경사는 분산분석(ANOVA)을 실시하였다. 임목구조는 평균값이 통계적으로 차이가 있는지 파악하기 위해 분산분석을 실시하였으며, 사후검정으로 Scheffe의 검정방법을 이용하였다.
유형별 종조성의 이질성을 파악할 수 있는 MRPP-test (Multi Response Permutation Procedures test)는 각 집단 간의 유의성 및 유사계수를 이용하여 군집의 이질성을 검증할 수 있으며(Berry et al., 1983; McCune and Grace, 2002), 거리 척도는 Sørensen을 사용하였다. 지표종 분석(indicator species analysis)은 확률화 검정(randomization test)을 이용하여 계산하였고, 지표종은 현재 특정지역에서 현존하며 우점하는 종으로(Dufrene and Legendre, 1997) 유의성(p<0.05) 있는 수종을 분석하였다.
현장조사 자료를 토대로 분비나무 치수를 ha당 본수로 환산하여 평균을 산출하였다. 종간 상대적 우세를 비교하기 위해 Curtis와 McIntosh(1951)의 기준으로 상대중요치(IV)를 계산하였으며 평균상대중요치(Mean importance value, MIV)는 층위별로 가중치를 부여한 (교목층 IV×3+아교목층 IV×2+관목층 IV×1+초본층 IV×0.5)/6.5로 산출하였다. 산림식생의 종 경쟁 및 다양성 등을 분석하기 위해 식생조사 자료를 바탕으로 층위별 종다양도(Shannon diversity index, H’), 최대종다양도(H’max), 균등도(Evenness, J’), 우점도(Dominance index, D)를 분석하였다(Shannon and Weaver, 1949; Brower and Zar, 1977). 산림식생의 안정성을 판단하기 위해 유형별로 분류한 군집 내 흉고직경 6 cm 이상의 목본류에 대한 흉고직경급 분포를 나타내었다.
종면적곡선, MRPP-test, 지표종분석, 종다양도는 PC-ORD7을 이용하였고, 분산분석은 IBM SPSS Statistics 25을 통해 산출하였다.
결과 및 고찰
본 연구대상지 내 나타난 관속식물은 67과 170속 286종 3아종 38변종 7품종으로 총 334분류군이 분포하는 것으로 조사되었다(Table 1). 이는 증거표본에 따른 한국 관속식물 2954종류(Korea National Arboretum, 2016)의 11.3%에 해당한다. 양치식물이 6과 16속 29종 2변종으로 31분류군으로 나타났으며, 나자식물이 3과 5속 6종 1변종으로 7분류군, 피자식물은 58과 149속 251종 3아종 35변종 7품종으로 총 296분류군으로 조사되었다.
방형구 내 희귀식물(Korea National Arboretum, 2022)은 CR(멸종위기종: Critically Endangered)의 산마늘, 산작약, 이삭단엽란 3분류군, EN(위기종: Endangered Species)의 가는잎개별꽃, 개회향, 눈향나무, 두메닥나무, 땃두릅나무, 만주송이풀 6분류군, VU(취약종: Vulnerble)의 금강초롱꽃, 나도옥잠화, 눈측백, 미역고사리, 백작약, 주목, 6분류군, LC(약관심종: Least Concern)은 게박쥐나물, 과남풀, 귀박쥐나물, 금강애기나리, 금마타리, 꽃개회나무, 덩굴꽃마리, 도깨비부채, 만병초, 말나리, 세잎종덩굴, 연영초, 태백제비꽃 13분류군, 21과 27속 25종 3변종으로 총 28분류군이 확인되었다.
방형구 내 특산식물(Korea National Arboretum, 2022)은 금강초롱꽃, 금마타리, 꽃황새냉이, 병꽃나무, 병조희풀, 분취, 삼도하수오, 숲개별꽃, 요강나물, 은꿩의다리, 지리대사초, 진범, 참개별꽃, 처녀치마, 청괴불나무, 한라사초, 할미밀망으로 10과 14속 15종 2변종으로 총 17분류군이 확인되었다.
종면적곡선은 종수 및 적정조사구 수를 추정하는 방법으로(Park and Seo, 2002) 본 연구대상지의 종면적곡선을 분석한 결과는 다음과 같다(Figure 2). 총 140개소의 조사지에 대해 수관층식생(교목층, 아교목층)과 하층식생(관목층, 초본층)으로 나누어 살펴보았을 때 기울기가 0에 수렴하는 것으로 나타나 데이터 분석에 적절한 조사구 수로 판단된다.
본 연구는 분비나무림에서 미역줄나무의 출현에 따른 식생구조 특성을 파악하고자 하였다. 미역줄나무의 출현 유무와 미역줄나무가 출현하는 층위에 따라 미역줄나무가 교목·아교목층에 출현하는 유형(TP), 미역줄나무가 관목층에 출현하는 유형(SP), 미역줄나무가 출현하지 않는 유형(A)으로 구분하였다. 유형 구분의 기준에 있어 층위별로 구분한 이유는 군집 내 경쟁에서 우위에 있는 상층부와 중층부의 식물종과 환경 조건에 따라 생육이 달라질 수 있는 하층 식물종은 임분 내 생태적 지위가 상이하기에(Sim et al., 2022) 미역줄나무와 분비나무의 관계를 파악하기에 더 적절할 것으로 판단되었다. 유형별 조사지 개수는 TP유형 11개소, SP유형 64개소, A유형 65개소로 분류되었다.
유형별 흉고단면적과 줄기밀도를 살펴본 결과 흉고단면적은 유의한 차이를 나타냈지만, 줄기밀도는 유의하지 않았다(Table 2). 분비나무의 흉고단면적은 TP유형이 17.2±13.7 m2/ha로 가장 높았고, SP유형이 10.8±10.2m2/ha이 가장 낮은 것으로 나타났다. 각 유형별 흉고단면적에는 통계적으로 유의한 차이를 보였다(p<0.001). 임분 내 총 흉고단면적은 TP유형이 13.76±19.03 m2/ha이 가장 높았고, SP유형이 9.29±11.67 m2/ha로 가장 낮은 것으로 분석되었다. 분비나무의 흉고단면적이 각 유형별 차이가 있는 것으로 나타났다(p<0.001). 임목밀도의 경우 유형별 유의한 차이는 나타나지 않았다(p<0.05).
Scheffe’s post hoc, *: p < 0.05
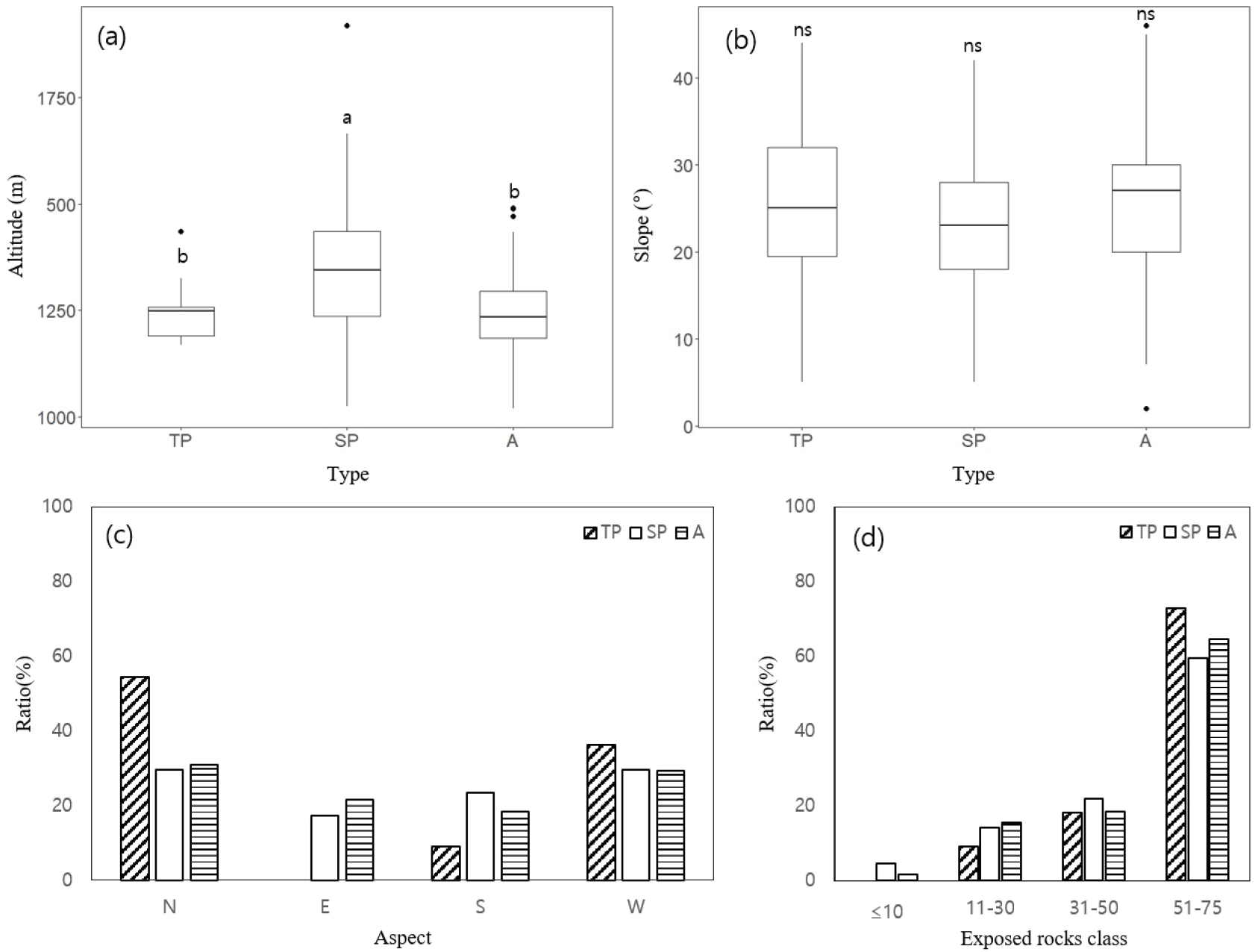
조사지의 임분구조를 살펴본 결과는 다음과 같다(Figure 3). 임분구조 중 사면방위는 북사면이 54.5-29.7%, 동사면이 21.5-0%, 남사면이 23.4-9.1%, 서사면이 36.4-29.2%의 분포로 대부분 북사면에 위치하였다. 암석노출도는 분비나무 생육 특성상 3유형 모두 50% 이상의 높은 암석노출도를 나타났다. 해발고도의 경우 SP유형이 1347.0±157.6 m로 나타나 다른 유형에 비하여 높은 해발고도를 보였고 각 유형 간 유의한 차이가 있는 것으로 나타났다(p<0.001). 경사는 A유형이 25.6±8.3°로 가장 급하였으나, 유의한 차이는 나타나지 않았다(p<0.05).
미역줄나무 출현에 따른 분비나무림의 식생 구조 특성을 파악하기 위하여 유형 간 층위별 구성종의 평균상대중요치와 종다양도를 비교 분석하였다.
유형별 식생구조 분석 결과(Table 3), 분비나무의 층위별 상대중요치는 미역줄나무가 교목·아교목층에서 출현하는 TP유형이 교목을 제외한 모든 층위에서 가장 낮게 나타났다. 미역줄나무의 중요치 변화는 초본부터 관목층까지 상대적으로 영양 경쟁을 증가시킬 수 있으며, 서식지에서 동시 출현하는 종조성에 영향을 끼칠 수 있다(Kim et al., 2020). 미역줄나무는 덩굴성 식물로 산림식생의 구조를 단순화시키고 빠른 성장으로 영양분과 빛에 우위를 차지하여 타 수종의 생장에 부정적인 영향을 끼친다(Kim et al., 2018). 특히, TP유형은 미역줄나무의 세력이 강한 유형으로 다른 유형에 비하여 분비나무의 생태적 지위가 낮은 것으로 판단된다.
T: Tree, ST: Subtree, S: Shrub H: Herb, MIP; Mean impotance value
분비나무림의 종다양성을 유형에 따라 층위별로 구분하여 종다양도(H’), 최대종다양도(H’max), 균등도(J’), 우점도(D)의 4가지 지수로 분석하였다(Table 4). 종다양도는 새로운 종이 유입되고 중간 정도의 교란과 생산성을 가지거나 종간 경쟁이 심한 식생 유형일수록 높게 나타난다(Schnitzer et al., 2001; Vasilevich, 2009). 균등도는 다양한 종이 특정 지역에 고르게 분포하면 증가하고 반대의 경우 감소한다(Norman et al., 2005).
H’ : Diversity, H’max : Max Diversity, J’ : Evenness, D : Dominance
층위별 종다양도 분석 결과(Table 5), 초본층을 제외한 모든 층위에서 TP유형이 다른 유형에 비해 상대적으로 종다양도(H’)가 높게 나타났으며, 종합적으로 TP유형의 종다양도가 가장 높게 나타났다. 미역줄나무의 출현은 임분 내 생산성을 감소시키고, 외부 유입종에 대한 정착기회를 증가시킨다(Norman et al., 2005; Kim et al., 2020). 이는 미역줄나무의 세력이 강한 TP유형에서 외부 유입종에 대한 지위경쟁력이 낮아 다른 유형에 비해 종다양도가 높게 나타난 것으로 판단된다.
| T | A | p | |
|---|---|---|---|
| TP vs SP | −0.662 | 0.006 | 0.230 |
| TP vs A | −3.608 | 0.034 | 0.004 |
| SP vs A | −14.402 | 0.074 | 0.000 |
MRPP 및 indicator species 분석을 이용하여 분비나무림에서 미역줄나무 출현에 따른 종조성의 이질성 및 지표종을 파악하고자 하였다((Table 5 및 Table 6). MRPP-test는 유형의 군집 간 이질성을 파악할 수 있으며 이를 종조성 유사도 A(Chance-corrected within-group agreement)값으로 나타낼 수 있고, 그 값은 0에서 1의 값을 가지게 되고 1에 가까울수록 유사한 종조성을 이룬다(Mielke and Berry, 1982; Cheon et al., 2014; Park et al., 2020).
IV max : maximum indicator value
MRPP-test 결과 대체적으로 미역줄나무의 출현을 층위별로 비교한 그룹에서는 유의한 차이가 없었지만, 미역줄나무의 출현 유무와 층위별 출현을 비교한 그룹에서는 유의한 차이가 있는 것으로 나타났다(Table 5). p값의 경우, 미역줄나무의 출현 유무와 층위로 구분하였을 때 TP유형와 SP유형(p=0.23)은 유의하지 않았고, TP유형와 A유형(p=0.004), SP유형과 A유형(p=0.000)은 유의한 것으로 나타났다. A값의 경우, TP유형과 SP유형은 0.006로 다른 유형에 비하여 더 이질적인 군집으로 나타났으나 그 유의성은 인정되지 않았다. TP유형과 A유형은 0.034, SP유형과 A유형은 0.074로 미역줄나무가 교목·아교목층에 나타난 TP유형이 관목층에서 나타난 SP유형보다 더 이질적인 종조성을 가진 것으로 분석되었다.
지표종(indicator species)은 특정 환경 조건이나 생태계의 특정 상태를 나타내는 종으로(Dufrene and Legendre, 1997) 이러한 종들은 환경 변화나 생태계의 변화를 지표로 사용할 수 있다. 유형에 따른 지표종을 층위별로 살펴보기 위한 지표종 분석 결과는 다음과 같다(Table 6).
교목층의 경우 TP유형에서는 당단풍나무가 지표종으로 나타났다. 아교목층의 경우 TP유형에서는 미역줄나무, 함박꽃나무, 다래, 부게꽃나무, 쥐다래, SP유형에서는 사스래나무가 나타났다. 관목층의 경우 TP유형에서는 다래, 부게꽃나무, 까치밥나무, 주목, SP유형에서는 미역줄나무로 나타났다. 초본층의 경우 TP유형에서는 십자고사리, 오리방풀, 개회나무, 나도옥잠화, 단풍취, 바늘사초, 주목, SP유형에서는 미역줄나무로 분석되었다. A유형은 모든 층위에서 뚜렷한 지표종이 나타나지 않았다.
미역줄나무를 대상으로 한 연구목적에 맞춰 TP, SP유형에서 미역줄나무가 지표종으로 나타나 구분한 유형이 알맞게 나눠진 것으로 나타났다. 또한 미역줄나무가 교목·아교목층에 출현한 TP유형에서 다래와 쥐다래가 지표종으로 분석되었는데, 미역줄나무는 국내 아고산 식생에서 숲 틈(forest gap) 발생 후 다수의 개체로 발생하는 종이며(Byeon et al., 2020; Cheon et al., 2022), 다래속 식물은 생장력이 왕성하고 생육속도가 빠른 특징이 있다(Nam et al., 2008). 덩굴성 식물은 교란과 함께 선구종으로 이입되는 종으로(Park et al., 2019) 해당 TP유형의 군집에서 교란의 영향이 있는지 지속적인 모니터링이 필요한 것으로 판단된다.
어린나무의 생육에는 충실한 종자의 공급, 종자 발아 및 활착에 적합한 지표면의 환경, 경쟁수종의 유무, 미기상 환경 등 다양한 요인이 영향을 미치며(Park et al., 2022), 나무의 생활사에 있어 성목에 비해 환경적 요인에 더욱 민감하게 반응한다(Kullman, 1993; Germino et al., 2002).
수고를 기준으로 분비나무 어린나무를 ha당 평균 개체수로 나타낸 결과는 다음과 같다(Figure 4). 분석 결과, 분비나무는 TP유형에서 10cm 미만의 등급에서는 개체수가 가장 많았으나 10 cm 이상의 등급에서는 다른 유형에 비하여 가장 적은 개체수로 나타났다. 특히 미역줄나무의 세력이 강한 TP유형에서 성목으로 성장할 가능성이 높은 50 cm 이상의 등급의 분비나무의 개체수가 상대적으로 낮게 나타났다.
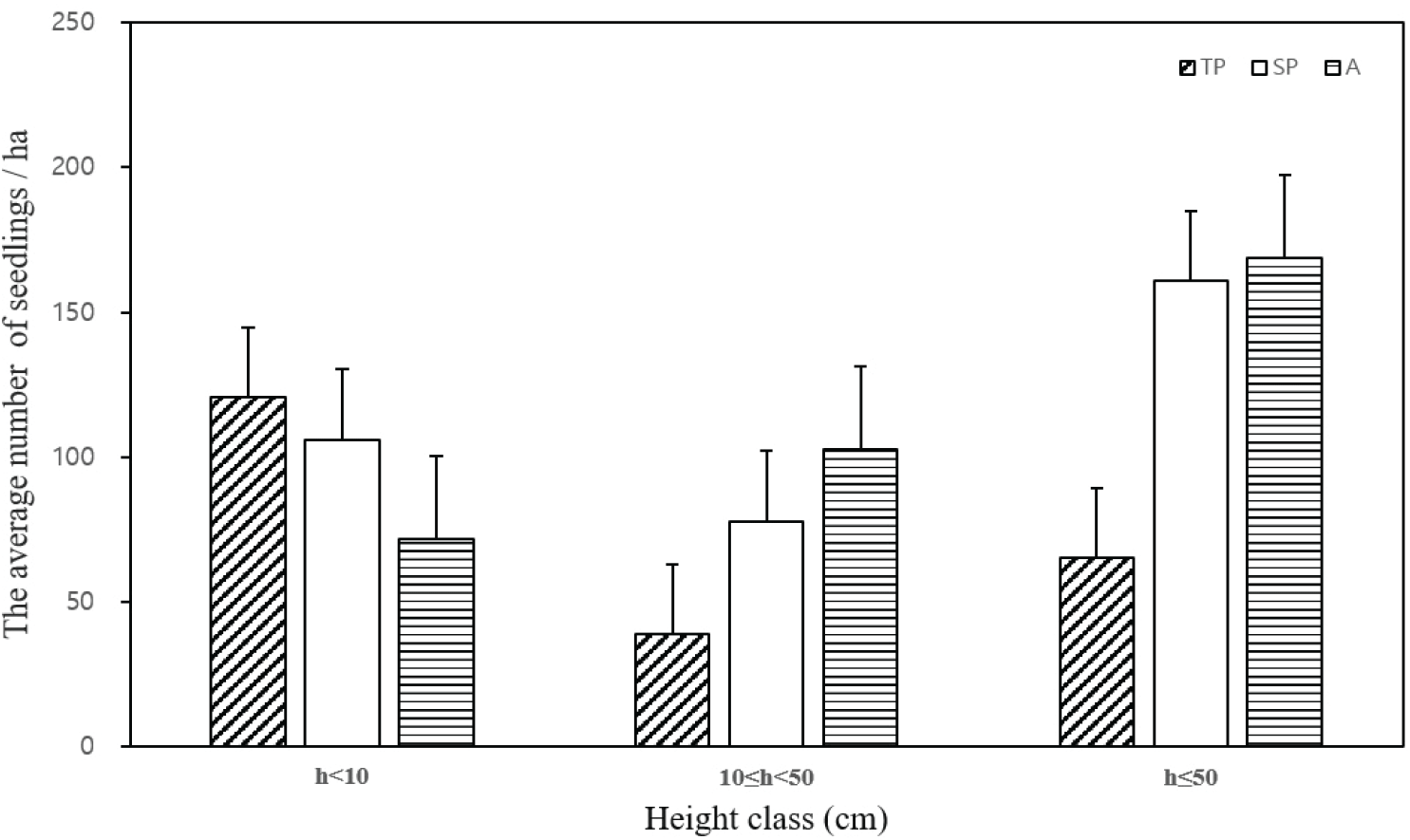
이는 설악산, 지리산 등 상록침엽수림의 서식지에서 미역줄나무의 중요치가 높을수록 분비나무, 구상나무 등 어린나무에 부정적인 영향을 끼쳐 타 수종의 생장을 저해한다는 연구결과와 유사하다(Kim et al., 2018; Cheon et al., 2022).
매목조사를 실시하여 얻은 자료를 통해 유형별 분비나무의 흉고직경급 분포를 비교하였다(Figure 5). 산림식생에서 흉고직경급 분포는 개체군의 지속성, 유지 가능성 여부를 추론하는 중요한 정보를 제공한다(Harcombe and Marks, 1978).
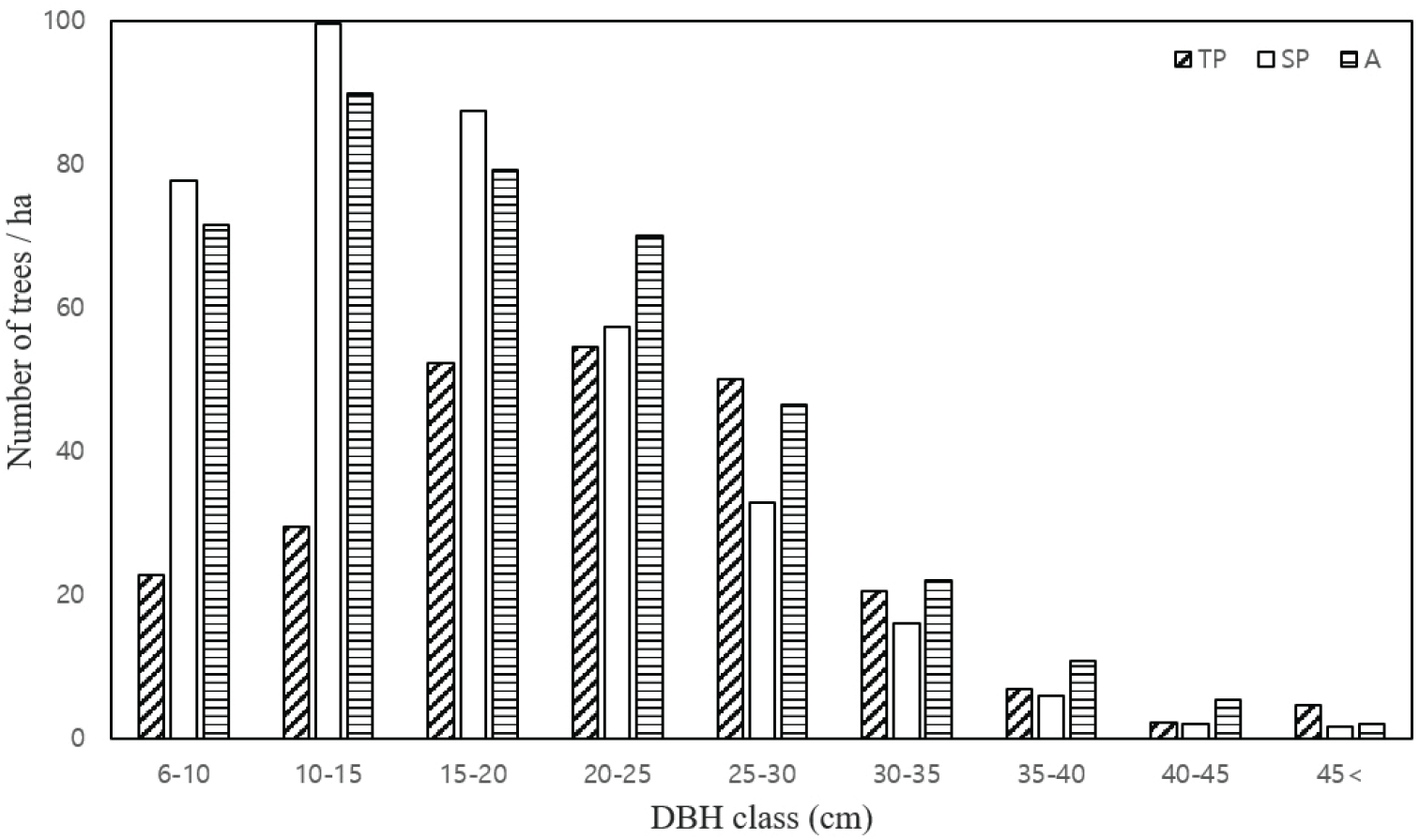
분비나무의 경우 20 cm 이하의 직경급에서 개체수의 차이가 비교적 크게 나타났으며, 총 개체수는 A유형 398개체/ha, SP유형 389개체/ha, TP유형 243개체/ha 순으로 나타났다. 6-25 cm 직경급에서 TP유형의 개체수가 가장 적었으나, 25 cm 이상의 직경급에서는 유형별 큰 차이는 나타나지 않았다. 층위별 중요치 분석결과와 함께 살펴보면, 모든층위에서 TP유형이 SP, A유형에 비하여 상대중요치 및 흉고직경급 분포에서 모두 낮은 수치를 나타내었다. 미역줄나무의 세력이 강한 TP유형에서 다른 유형에 비하여 분비나무의 지위가 부진한 것으로 나타났다.
결 론
본 연구는 미역줄나무 출현에 따른 분비나무림의 식생구조 특성을 파악하여 분비나무림의 보전 및 관리에 기여하고자 하였다. 평균상대중요치 분석, 종다양도 분석, MRPP-test, 지표종 분석, 어린나무 분석, 흉고직경급 분석을 통해 연구를 진행하였다. 그 결과 구분한 유형별 군집의 종조성은 이질적이며 미역줄나무의 출현이 어린나무의 생육과 분비나무림의 식생구조에 부정적인 영향을 미치는 것으로 나타났다. 미역줄나무가 교목·아교목층에 출현한 TP유형은 다른 유형에 비해 분비나무의 어린나무 직경급의 개체수와 치수 발생량이 현저히 낮게 나타나 분비나무가 미역줄나무와의 지위경쟁에 어려움이 있는 것으로 판단된다.
아고산 침엽수종이 당면한 문제는 급격한 기후변화로 인한 쇠퇴와 집단 고사뿐만 아니라 후대로 성장할 어린나무의 생육이 부진한 점이다(Park et al., 2015; Kim et al., 2018; Park et al., 2022). 미역줄나무와 분비나무의 관계를 더욱 명확히 파악하기 위해서는 추후 미역줄나무의 인위적인 제거 연구와 치수 발생량에 대한 관찰이 필요하다.
본 연구는 우리나라 분비나무를 대상으로 미역줄나무 출현에 따른 분비나무의 지속성을 파악하고자 하였다. 기후변화로 인해 지구 온도가 상승함에 따라 덩굴의 성장 속도가 더욱 증가할 것으로 예측되는 가운데(Phillips et al., 2002), 본 연구가 분비나무뿐만 아니라 멸종위기 고산 침엽수종의 보전 및 관리에 기여하고 기후변화로 급격한 변화를 겪고 있는 생태계를 이해하는데 보탬이 될 것으로 기대한다.