Article
낙엽송과 리기다소나무 벌채지에 조성된 낙엽송 임분의 11년간 토양 물리·화학적 특성 변화
노남진1
,
한승현2,
이상태2,
조민석3,*
Changes in Soil Physiochemcial Properties Over 11 Years in Larix kaempferi Stands Planted in Larix kaempferi and Pinus rigida Clear-Cut Sites
Nam Jin Noh1,
Seung-hyun Han2,
Sang-tae Lee2,
Min Seok Cho3,*
Author Information & Copyright ▼
1Division of Forest Science, Kangwon National University, Chuncheon 24341, Korea
2Forest Technology and Management Research Center, National Institute of Forest Science, Pocheon 11187, Korea
3Research Planning and Coordination Division, National Institute of Forest Science, Seoul 02455, Korea
© Copyright 2023 Korean Society of Forest Science. This is an Open-Access article distributed under the terms of the
Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits
unrestricted non-commercial use, distribution, and reproduction in any
medium, provided the original work is properly cited.
Received: Nov 09, 2023; Revised: Nov 24, 2023; Accepted: Nov 27, 2023
Published Online: Dec 31, 2023
요 약
본 연구는 낙엽송과 리기다소나무 벌채지에 조성된 낙엽송 조림지의 토양 물리·화학적 특성 및 조림목 생장의 장기적인 변화를 이해하고자 수행되었다. 낙엽송 전생임분(춘천, 김천)과 리기다소나무 전생임분(원주, 가평)에 낙엽송 노지묘(1-1)를 3,000본 ha−1 밀도로 2009-2010년에 식재하였다. 조림 당해연도와 식재 후 3, 7, 11년에 토양 시료(0-20 cm)를 채취하여 물리·화학적 특성을 분석하였으며, 동시에 주기적으로 수고 및 근원경을 측정하였다. 연구 결과, 식재 초기 미사와점토 함량, 총탄소와 전질소, 유효태인산 농도, 양이온치환용량 등의 토양 특성은 전생임분에 따른 차이를 보였으나, 조림목 생장은 전생임분에 따른 차이를 보이지 않았다. 토양의 화학적 특성은 전반적으로 김천지역이 가장 양호한 반면, 조림목 초기 생장은 가평지역에서 가장 높게 나타났다. 조림 11년 경과 후에는 어린나무가꾸기 작업에 따라 임분밀도가 크게 감소한 원주(1,028본 ha−1)와 춘천(1,359본 ha−1)에서 흉고직경이 더 크게 나타났다. 한편 조림 초기 전생임분과 조림지별 토양 특성의 차이는 낙엽송 조림 11년 경과 후 유사하게 변화하였다. 특히 벌채·조림 후 높게 나타났던 모래함량과 유효태인산 농도는 잠재적으로 유기물 유입과 양분 흡수에 의해 크게 감소하였다. 본 연구는 벌채 후 조기 재조림이 양분 용탈을제한하고 토양 안정화에 기여할 수 있음을 제안하고, 낙엽송 재조림지의 양분 관리에 유용한 정보를 제공한다.
Abstract
This study was conducted to understand the long-term changes in soil physiochemical properties and seedling growth in Larix kaempferi (larch) stands planted in clear-cut larch and Pinus rigida (pine) forest soils over an 11-year period after reforestation. Two-year-old bare-root larch seedlings were planted in 2009-2010 at a density of 3,000 seedlings ha−1 in clear-cut areas that harvested larch (Chuncheon and Gimcheon) and pine (Wonju and Gapyeong) stands. We analyzed the physiochemical properties of the mineral soils sampled at 0-20 cm soil depths in the planting year, and the 3rd, 7th and 11th years after planting, and we measured seedling height and root collar diameter in those years. We found significant differences in soil silt and clay content, total carbon and nitrogen concentration, available phosphorus, and cation exchangeable capacity between the two stands; however, seedling growth did not differ. The mineral soil was more fertile in Gimcheon than in the other plantations, while early seedling growth was greatest in Gapyeong. The seedling height and diameter at 11 years after planting were largest in Wonju (1,028 tree ha−1) and Chuncheon (1,359 tree ha−1) due to decreases in stand density after tending the young trees. The soil properties in all plantations were similar 11 years after larch planting. In particular, the high sand content and high available phosphorus levels (caused by soil disturbance during clear-cutting and planting) showed marked decreases, potentially due to soil organic matter input and nutrient uptake, respectively. Thus, early reforestation after clear-cutting could limit nutrient leaching and contribute to soil stabilization. These results provide useful information for nutrient management of larch plantations.
Keywords: field performance; harvesting; reforestation; soil nutrient; soil texture
서 론
임목수확기에 도래한 산림을 적정한 시기에 벌채하고 적합한 수종으로 재조림하여 산림의 생산성을 향상시키는 것은 지속가능한 산림관리에 필수적이며, 산림 탄소흡수 기능 증진을 통한 산림부문 탄소중립 이행에도 기여할 수 있다(Domke et al., 2020; Picchio et al., 2020; Lefebvre et al., 2021). 그러나 동시에 집약적인 목재 수확 및 재조림 과정 동안 발생할 수 있는 교란을 최소화하고 생태계 구성요소들의 안정성을 유지해야 한다(Schweier et al., 2019).
개벌(모두베기)은 수확의 효율성을 달성하기 위한 일반적인 관리 방법이지만 벌채 후 재조림 단계 동안 생태계에 생물적·비생물적 교란을 초래할 수 있다(Povilaitienè et al., 2022). 특히 토양 물리·화학적 특성은 모두베기에 영향을 받을 수 있다(Siebers and Kruse, 2019; Picchio et al., 2020). 산림 바이오매스를 제거하면 토양 유기물과 탄소 축적량이 감소하며(Smith et al., 2022), 강도 높은 바이오매스 제거는 토양 양분 손실로 이어질 수 있다(Piirainen et al., 2007; Caldeira et al., 2023). 이는 잠재적으로 재조림지의 생산성을 감소시킬 수 있다. 최근 Povilaitienè et al. (2022)은 북유럽 구주소나무(Pinus sylvestris) 임분을 대상으로 모두베기와 재조림 후 4년간 토양의 생물·화학적 특성 변화에 대해 보고하였고, Farahnak et al. (2022)은 일본 편백(Chamaecyparis obtusa) 임분을 대상으로 모두베기에 따른 토양 화학성 변화에 대해 보고한 바 있다.
우리나라에서도 점차 벌채 및 재조림 면적이 확대됨에 따라 산림 시업에 따른 토양 특성 변화에 대한 관심이 높아지고 있다. 그러나 벌채 및 재조림이 산림 토양에 미치는 영향에 대한 연구 결과는 매우 부족한 실정이다. 목재 수확 후 재조림에 따른 토양 물리·화학적 특성의 시간적 변화는 조림지의 양분관리 측면에서 조림 성과 및 목재 생산성 향상을 위해 중요한 정보를 제공할 수 있다. 최근 국내에서도 목재수확 과정에서 기계화작업이 토양 답압에 미치는 영향(Han et al., 2019; Baek et al., 2022) 및 재조림 초기 토양의 물리·화학적 특성 변화(Yang et al., 2017, 2018) 등에 대한 연구가 수행된 바 있다. 그러나 재조림 이후 토양 특성 변화를 장기적으로 분석한 연구 결과는 보고된 바 없다.
낙엽송(Larix kaempferi (Lambert) Carrière)은 소나무, 편백 등과 함께 우리나라 주요 조림 수종 중 하나로 2021년 기준 총 4,721 ha의 면적에 조림되었으며, 목재 수요 증가 및 벌기령 도달에 따라 벌채·조림 면적 및 묘목 생산이 지속적으로 확대되고 있다(Korea Forest Service, 2022). 한편 리기다소나무(Pinus rigida Mill.)도 1960년대 우리나라 주요 조림 수종 중 하나로 낙엽송(약 26.0만ha) 다음으로 많은 면적(약 23.4만ha)을 차지하며, 특히 벌기령에 도달한 불량임분의 경우에는 수종갱신의 필요성이 제기되고 있다(Korea Forest Service, 2022). 낙엽침엽수인 낙엽송은 상록침엽수 리기다소나무에 비해 양분요구도가 높고 양분순환율이 빠른 수종으로, 다른 두 수종이 우점하는 성숙림은 임상유기물 및 토양 특성이 다를 수 있다(Son and Gower, 1991; Kim et al., 2010). 즉 벌채 이전, 수종이 다른 임분 간에는 토양 특성 차이를 보일 수 있으며(Binkley and Menyailo, 2005; Russell et al., 2007), 이러한 전생임분 차이에 기인한 토양 특성은 조림목 초기 생장에도 영향을 미칠 수 있다(Yang et al., 2018). 따라서 본 연구는 낙엽송과 리기다소나무 전생임분을 벌채하고 낙엽송을 재조림한 임분을 대상으로 토양 물리·화학적 특성과 조림목 생장의 장기적 변화를 분석하고자 수행되었다.
재료 및 방법
1. 연구 대상지 및 조림목 식재
본 연구는 2009-2010년 국유림에 조성된 낙엽송 조림지 4곳을 대상으로 수행되었다. 강원특별자치도 춘천과 경상북도 김천 지역은 낙엽송 임분을 벌채 후 낙엽송을 재조림하였으며, 강원특별자치도 원주와 경기도 가평 지역은 리기다소나무 임분을 벌채하고 낙엽송을 재조림하였다. 김천 조사지는 2009년에, 춘천, 원주, 가평 조사지는 2010년에 낙엽송 노지묘(1-1)를 3,000본 ha−1의 동일한 밀도로 식재하였다. 각 조림지별 연평균 기온 및 강수량은 인접 지역의 기상청 관측자료(2009-2020)를 활용하였으며, 해발고가 높은 김천과 춘천 조림지의 연평균 기온은 산림청 산악기상관측시스템으로부터 조사지와 가장 인접한 지점의 관측자료(2019-2020)를 활용하였다. 각 임분 특성은 Table 1과 같다.
Table 1.
Site characteristics of four Larix kaempferi plantations.
| Study site |
Chuncheon (CC) |
Gimcheon (GC) |
Gapyeong (GP) |
Wonju (WJ) |
| Location (N, E) |
37°58′23.81″, |
35°50’43.16″, |
37°43’37.78″, |
37°23’37.21″, |
| 127°50’25.31″ |
128°03’05.39″ |
127°33’27.28″ |
127°48’06.71″ |
| Elevation (m a.s.l.) |
420-438 |
712-872 |
128 |
200 |
| Slope (°) |
21-25 |
25-28 |
27 |
31 |
| Aspect (°) |
N, SE |
S, NW |
NW |
NE |
| Mean annual temperature (°C) |
10.0† |
11.0† |
11.7 |
12.5 |
| Mean annual precipitation (mm yr−1) |
1101 |
1310 |
1357 |
1228 |
| Management records |
| Planting year |
2010 |
2009 |
2010 |
2010 |
| Planting density (trees ha−1) |
3,000 |
| Seedling type |
1-1 bare root seedling |
| Year of forest tending work |
2017 |
2016 |
none |
2017 |
| Stand density (trees ha−1) in 2021‡ |
1,359±175 |
1,873±104 |
2,770±246 |
1,028±207 |
| Stand properties before harvesting |
| Species |
Larix kaempferi
|
Pinus rigida
|
| Age class |
VIII |
VIII |
V |
VI |
| Mean DBH (cm) |
18 |
21 |
16 |
10 |
| Mean height (m) |
28 |
32 |
17 |
16 |
Download Excel Table
2. 생장 조사
조림지 조성 후, 각 조사지에 20 × 20 m 조사구를 3개씩 설치하였으며, 조림목의 생장 특성을 조사하기 위하여 각 조사구 내에서 30본씩 총 90본의 조림목을 2010년부터 2014년까지 매년 9월 말 캘리퍼와 절척을 이용하며 근원경(cm)과 수고(m)를 측정하였다. 2015년 이후부터는 2~3년 주기로 근원경과 수고를 측정하였으며, 2021년에는 모든 조사지에서 흉고직경을 측정하였다. 한편 김천지역 조림지의 경우 다른 지역과 조림지 조성 시기의 차이가 있어, 동일한 수령을 기준으로 비교·분석하였다.
3. 토양 조사
토양 시료는 조림 당해연도인 2010년과 2013년, 2017년, 2021년 4회에 걸쳐 채취하였다. 토양 시료는 직경 5.5 cm 토양시료 채취기를 이용하여 조사구 내 3~5 지점에서 토심 0~20 cm 깊이에서 채취하였다(n=9~15). 채취한 토양 시료는 48시간 이상 충분히 음건 후 2 mm체(US standard no.10)로 쳐서 토양의 물리성 및 화학성 분석에 이용하였다. 토성은 비중계법을, 토양 산도(pH)는 1:5 증류수로 희석하여 pH 미터로 측정하였다. 전질소와 총탄소 농도는 건식산화연소법으로 원소분석기(Vario Max cube, Elementar Analyzer, Germany)를 이용하여 측정하였다. 유효태인산농도는 Lancaster법, 치환성양이온(K+, Na+, Mg2+, Ca2+) 농도는 Brown No.1법을 이용하여 1N 초산암모늄으로 추출한 추출액을 이온분석기(ICP-OES, Optima 8300, PerkinElmer, Singapore)를 이용하여 측정하였다.
4. 통계 분석
조림지 내 토양 특성에 조사지와 조림 경과시기 및 전생 임분과 조림 경과시기가 미치는 영향을 검정하기 위해 2-way ANOVA 분석을 실시하였고, 처리 간 유의적 차이는 HSD-test를 수행하였다(p<0.05). 토양 특성 인자들 간 주요인분석(Principle Component Analysis)을 수행하였으며 ‘ggbiplot’ 패키지를 이용하였다. 주요한 토양 인자들 간의 상관관계는 Pearson 상관분석을 실시하였으며, 모든 통계분석은 R 소프트웨어를 사용하였다(R software, 2017).
결과 및 고찰
1. 토양 물리·화학적 특성 변화
1) 토성
2010년 조림 초기 김천 조사지의 토성은 모래의 함량이 80±5%(평균±표준오차)인 양질사토로 다른 지역의 58±3~ 64±1%의 범위에 비해 유의적으로 높게 나타났다[Table 2, Figure 1(a)]. 그러나 조림 10년 경과 후 2021년 김천 조사지의 토성은 모래 함량이 47±3%로 감소하였고, 미사의 함량이 43±1%로 증가하여 양토로 나타났으며, 다른 지역(44±10~53±1)과 유사한 수준을 보였다(Table S2). 김천, 춘천, 가평, 원주 조사지 모두 2010년부터 2021년까지 모래 함량은 지속적으로 감소하였고, 미사와 점토 함량은 증가하는 경향을 보였다[Figure 1(b)]. 재조림을 위한 모두베기 과정에서 토양 답압 및 입단 구조 파괴 등의 교란이 발생할 수 있으며(Siebers and Kruse, 2019; Nazari et al., 2023), 토양 층위 발달이 미흡한 우리나라 산림의 경우, 목재 수확 과정에서 토양 교란으로 무기물표층과 조립질 모재층이 혼합 또는 역전될 때 모래 함량이 일시적으로 높게 나타날 수 있다. 그러나 시간이 경과함에 따라 낙엽의 유입과 분해가 토양 구조를 빠르게 안정화시키고, 토성 변화에 영향을 미칠 수 있다(Lajtha et al., 2014).
Table 2.
The results of two-way ANOVA of the year, site, former stand and their interaction effects on the variables of growth and soil property.
| Variables |
Year (Y) |
Site (S) |
Former stand (F) |
Y × S |
Y × F |
| Growth |
| Root collar diameter |
<0.001 |
<0.001 |
n.s. |
<0.001 |
0.014 |
| Height |
<0.001 |
<0.001 |
n.s. |
<0.001 |
0.006 |
| Soil |
| Sand |
<0.001 |
0.016 |
n.s. |
0.019 |
0.106 |
| Silt |
<0.001 |
0.007 |
0.027 |
n.s. |
n.s. |
| Clay |
0.008 |
<0.001 |
<0.001 |
0.010 |
0.007 |
| pH |
n.s. |
<0.001 |
n.s. |
n.s. |
n.s. |
| Total carbon |
<0.001 |
<0.001 |
<0.001 |
<0.001 |
n.s. |
| Total nitrogen |
0.083 |
<0.001 |
<0.001 |
n.s. |
<0.001 |
| P2O5− |
<0.001 |
0.006 |
0.042 |
0.006 |
<0.001 |
| C.E.C. |
0.002 |
<0.001 |
<0.001 |
n.s. |
n.s. |
| K+ |
<0.001 |
<0.001 |
0.004 |
0.002 |
n.s. |
| Na+ |
n.s. |
n.s. |
n.s. |
0.006 |
<0.001 |
| Ca2+ |
n.s. |
<0.001 |
n.s. |
<0.001 |
0.010 |
| Mg2+ |
<0.001 |
<0.001 |
0.006 |
<0.001 |
<0.001 |
Download Excel Table
Figure 1.
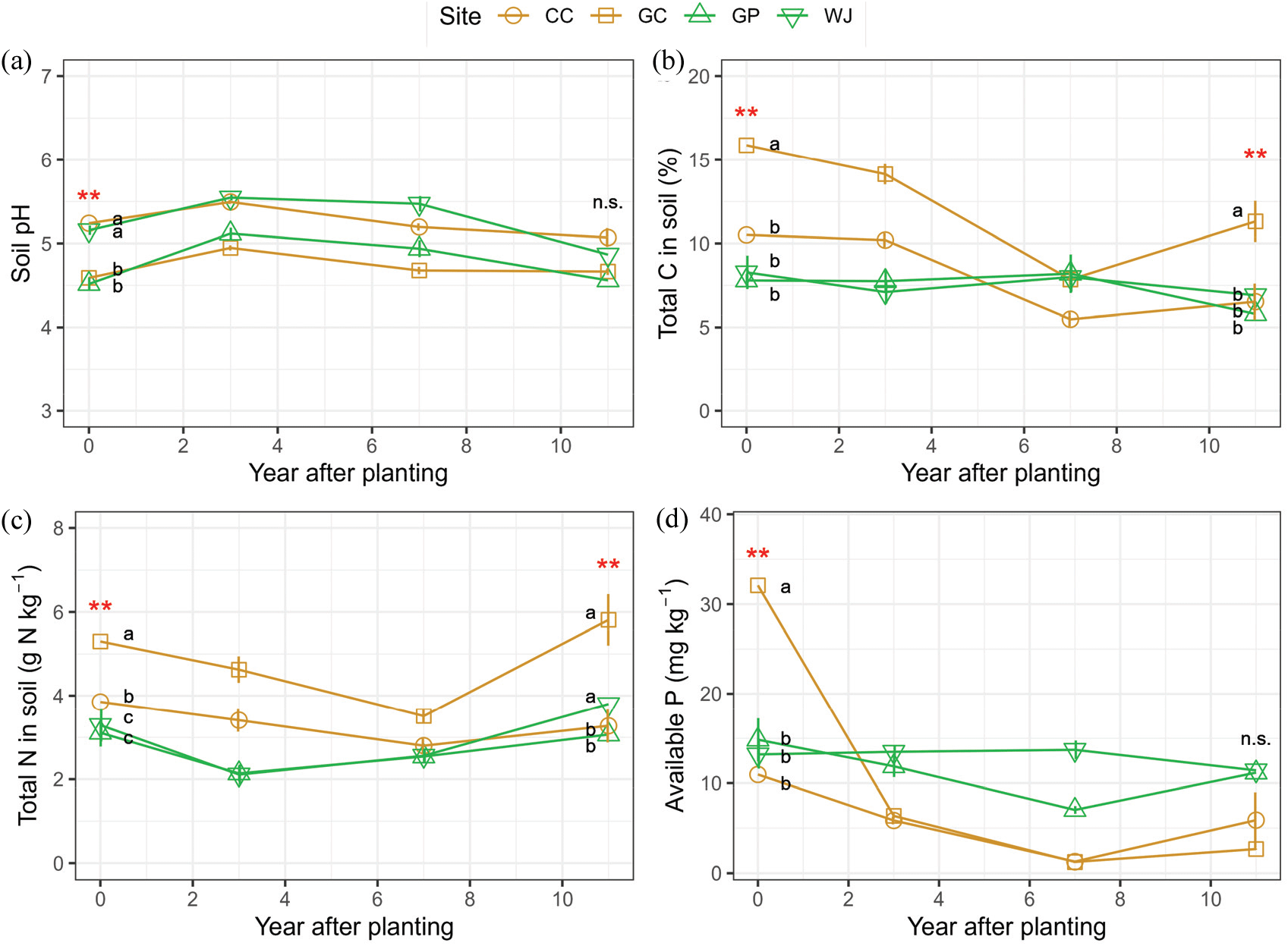
Temporal changes in sand content (a), silt plus clay contents (b) over 11 years after planting Larix kaempferi trees in harvested L. kaempferi (yellow) and Pinus rigida (green) plantations. CC: Chuncheon, GC: Gimcheon, GP: Gapyeong, WJ: Wonju. Vertical bars indicate standard deviation of the mean (n=9~15). Asterisks indicate significant differences in the variables between the study sites by ANOVA (p < 0.05).
Download Original Figure
2) 토양 산도(pH)
2010년 토양 pH는 김천(4.59±0.05)과 가평(4.52±0.08) 조사지가 춘천(5.24±0.09)과 원주(5.16±0.06) 조사지에 비해 낮게 나타났다[p<0.001, Figure 2(a), Table S2]. 조림 초기 토양 pH는 전생임분(낙엽송, 소나무)의 영향을 받지 않았으며, 조림 경과 시간에 따른 유의미한 차이는 나타나지 않았다(p>0.05, Table 2). 그러나 모든 조사구에서 조림 3년 후 2013년에 일시적으로 토양 pH가 증가하는 경향을 보였고, 점차 감소하여 조림 10년 경과 후 2021년의 토양 pH는 조림 초기와 유사하게 나타났으며, 이때 토양 pH는 조사지 간 통계적인 차이를 보이지 않았다[Figure 2(a)]. Povilaitiené et al.(2022)에 의한 북유럽 P. sylvestris 조림지에서의 연구는 모두베기 및 재조림 이후 0-20 cm 토양 산도(pHCaCl2)는 미벌채지 성숙림 4.0 이하에 비해 높았고, 조림 3년차에 4.4±0.2까지 지속적으로 증가하였으며, 조림 4년차에는 다시 감소하는 경향으로 나타나 본 연구와 유사한경향성을 보였다. 또한 일본 C. obtusa 조림지에서의 연구는 모두베기가 표층토(0-5 cm)에서의 토양 pH를 유의적으로 증가시키는 결과를 나타냈다(Farahnak et al., 2022). 이러한 증가는 모두베기 이후 수목으로부터 공급되는 용해성 수소이온(H+)의 감소와 무기토양으로부터 칼슘(Ca2+), 마그네슘(Mg2+) 등의 2가 이온의 증가에 기인한다고 보고하였다(Farahnak et al., 2022). 한편 조림목과 하층식생에 의한 지하부 뿌리발달 및 토양안정화, 유기물의 유입 등으로 인해 점차 감소할 수 있다. 그러나 스웨덴 P. sylvestris와 Picea abies가 우점하는 침엽수림에서 모두베기 이후 8년 동안 자연적인 식생 회복에 따라 유기물층과 무기물층 모두 pH가 지속적으로 증가하는 경향을 나타내기도 하였다(Grønflaten et al., 2008).
Figure 2.
Temporal changes in soil pH (a), total carbon (b), total nitrogen (c), and available phosphorous (P2O5-) over 11 years after planting Larix kaempferi trees in harvested L. kaempferi (yellow) and Pinus rigida (green) plantations. CC: Chuncheon, GC: Gimcheon, GP: Gapyeong, WJ: Wonju. Vertical bars indicate standard deviation of the mean (n=9~15). Asterisks indicate significant differences in the variables between the study sites by ANOVA (p < 0.05).
Download Original Figure
3) 총탄소 농도
2010년 평균 총탄소 농도(%)는 낙엽송 전생임분인 김천(15.9±0.8) 조사지가 낙엽송 전생임분인 춘천(10.5±0.6), 리기다소나무 전생임분인 원주(8.3±1.0), 가평(7.8±0.4) 조사지에 비해 높게 나타나 조사지 간 큰 변이를 보였다[p<0.001, Figure 2(b), Table S2]. 탄소 농도는 모두베기 수확 후 조림 초기에 가장 높았고 2013년, 2017년까지 급격하게 감소하는 경향을 보였으며, 2021년에는 이전 수준을 유지 또는 증가하는 경향을 보였다(p<0.001). 일반적으로 모두베기 또는 간벌 과정에서 발생하는 낙엽·낙지 등 임상 잔존물과 지하부 고사근의 일시적 증가는 토양 유기탄소의 농도 증가에 영향을 미칠 수 있다(Kim et al., 2009; Siebers and Kruse, 2019). 선행 연구에서 P. sylvestris 벌채 후 재조림지에서의 토양 탄소 농도는 조림 후 경과 시간에 따라 유의적인 차이를 보이지 않았으나, 조림 3년차까지 감소하는 경향을 보였고, 4년차에 다소 증가하는 경향을 나타내 본 연구 결과와 유사한 경향성을 보였다(Povilaitiené et al., 2022). 한편 본 연구에서 조림 초기 조사지 간 총탄소 농도 차이는 전생임분 수종 차이에 기인하는 것으로 보이며, 이는 리기다소나무 침엽에 비해 낙엽송 침엽의 빠른 분해율 때문으로 판단된다(Kim et al., 2010). 이러한 수종 간 분해율의 차이는 낙엽송 전생임분인 김천과 춘천 조림지에서 모두베기 후 재조림 시간 경과에 따라 탄소 농도가 급격하게 감소하는 것에서도 확인할 수 있다.
4) 전질소 농도
2010년 조사지별 평균 전질소 농도(g kg−1)는 탄소 농도와 유사한 경향을 보였으며[Figure 3(a)], 낙엽송 전생임분인 김천(5.30±0.21) 조사지가 낙엽송 전생임분인 춘천(3.85 ±0.28), 리기다소나무 전생임분인 가평(3.29±0.39), 원주(3.11 ±0.33) 조사지에 비해 높게 나타났다[p<0.001, Figure 2(c), Table 2]. 이후 2013년, 2017년까지 전질소 농도는 감소하는 경향을 보였으며, 2021년에는 전질소 농도가 다시 증가하여 조림 초기와 유사한 수준으로 나타났으나, 연변화에 따른 통계적 차이는 나타나지 않았다(p=0.083). P. elliotti 벌채 수확 후 잔존물 존치구에서는 제거 처리한 곳에 비해 토양 전질소 농도가 증가한다고 보고된 바 있으나(Chen and Xu, 2005), 본 연구의 경우 벌채 이전 토양시료 분석의 부재로 벌채 후 임상유기물 증가로 인한 토양 전질소 농도의 유의미한 증가는 확인할 수 없었다. 그러나 모두베기 및 재조림으로 인한 조림지 초기 토양 온도와 수분의 증가는 임상유기물의 분해율 증가 및 질산화과정을 촉진시킬 수 있고, 강우 유입시 질산태질소의 용탈이 크게 증가할 수 있다(Jussy et al., 2004; Hedwall et al., 2013; Povilaitiené et al., 2022). 그러나 양분유효도의 증가로 인해 임분 발달 초기 지피 식생의 빠른 회복에 기여할 수도 있다(Cesonienè et al., 2019). 한편 조림 10년 경과 후 2021년의 전질소 농도가 다소 증가한 것은 낙엽송 조림목의 생산성 증가로 인한 임상유기물의 증가에 기인한 것으로 보인다.
Figure 3.
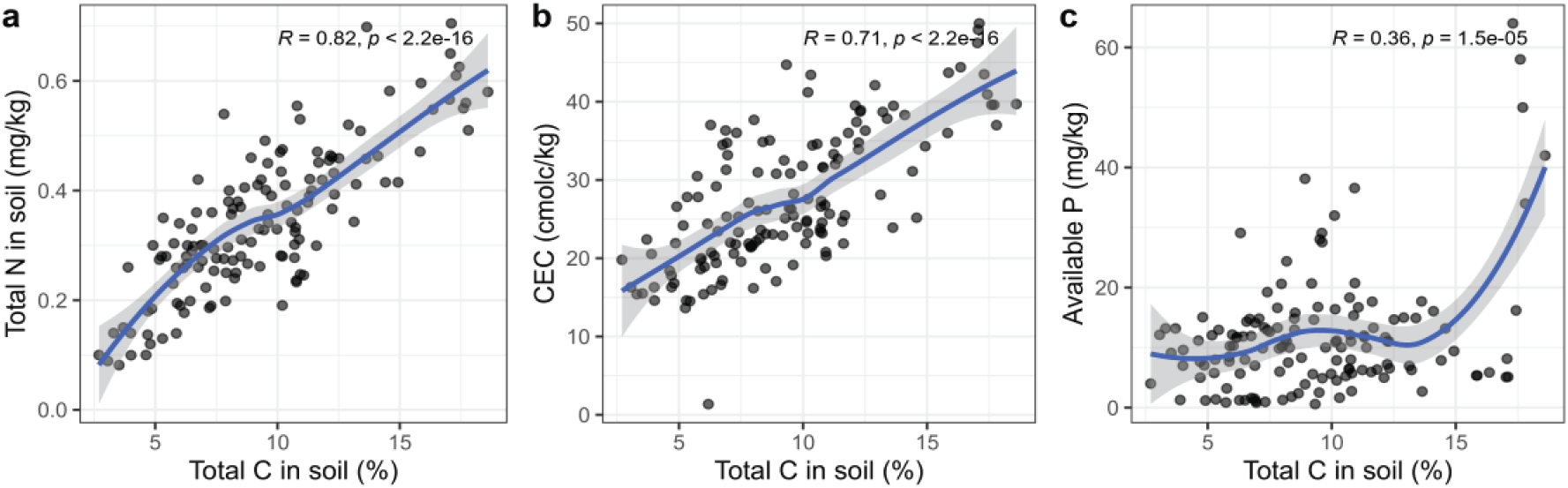
Relationships between total soil carbon and total nitrogen (a), cation exchangeable capacity (b), and available phosphorus (mg P2O5− kg−1) across the study sites over 11 years after planting. The correlation coefficients (R) and p-values are shown in the figures. The non-linear lines with 95% confidence intervals were fitted to the data.
Download Original Figure
5) 유효태인산
2010년 평균 유효태인산 농도(mg kg−1)는 낙엽송 전생임분 김천(32.1±6.4) 조사지가 가장 높고, 춘천(11.0±0.6), 원주(13.2±1.6), 가평(14.8±2.4)은 유사한 수준을 보였다[p<0.006, Table 2, Figure 2(d)]. 유효태인산 농도는 시간 변화에 따라 감소하는 경향을 보였으며, 특히 낙엽송 전생임분이었던 김천과 춘천의 경우 조림 경과 10년 후 각각 2.7±0.1 mg kg−1로 92%, 5.9±3.1 mg kg−1로 46% 감소하였다[Figure 2(d)]. 본 연구에서 유효태인산 농도는 조사지 간 변이가 크게 나타났으나 2020년 우리나라 낙엽송 재조림지 120개 토양의 유효태인산 농도 범위(1.0~69.5 mg kg−1) 내에 있는 것으로 나타났다(Cho, unpublished). 인의 주요 공원급은 무기광물인 인회석의 풍화와 식생으로부터 유입되는 유기물이다. 느리게 진행되는 무기광물의 풍화에 비해, 모두베기와 재조림 과정에서의 일시적으로 증가한 유기물 유입과 분해가 조림 초기의 유효태인산 농도에 영향을 미칠 수 있다. 본 연구에서 총탄소 농도와 유효태인산 농도는 낮은 수준에서 정의 상관 관계를 보였다[Figure 3(c), r=0.36, p<0.001]. Povilaitiené et al.(2022)의 연구에 따르면 유효태인산의 농도가 P. sylvestris 조림 1년차에 169.8 mg kg−1로 가장 높았으며 조림 4년차에 107.9로 약 36% 감소하였다. Yang et al.(2017)에 의한 선행 연구에서 조림 초기 유효태인산 농도의 감소의 잠재적 원인을 조림목에 의한 흡수와 강우에 의한 용탈이나 침전으로 추정한 바 있다. 우리나라를 포함하는 온대 산림에서 임목의 인요구도는 높은 편이며(Rodionov et al., 2020), 특히 산성 토양 내 인의 가용성이 낮기 때문에 유실량이 크지 않을 수 있고, 임목에 의해 흡수된 인화합물은 체내 이동성이 크기 때문에 조림목 내부에 상당량 축적될 가능성도 있다. 모두베기가 토양 유효태인산 농도에 영향을 미치지 않았다는 연구 결과도 보고된 바 있으나(Pennock and van Kessel, 1997), P. sylvestris 벌채지와 1년차 재조림지 0-20 cm 무기토양의 유효태인산 농도가 성숙림에 비해 1.5배, 2.1배 높게 보고된 바 있다(Povilaitiené et al., 2022). 따라서 토양 내 인순환에 대한 이해를 높이기 위해서는 벌채 이전 성숙림 토양과 임목의 부위별 인 농도 분석이 중요하다(Seidel et al., 2022). 일반적으로 산림 수목의 높은 인요구도와 수목생육 적정 기준(60~200 mg kg−1)을 고려할 때, 본 연구에서 낙엽송 재조림지의 인 가용성은 조림 초기부터 지속적으로 낮은 수준을 보였다. 따라서 본 연구에서의 조림지는 인이 생장 제한인자로 작용할 수 있어 토양 인의 관리를 통해 조림목 생장을 향상시킬 수 있을 것으로 보인다.
6) 양이온치환용량(CEC)
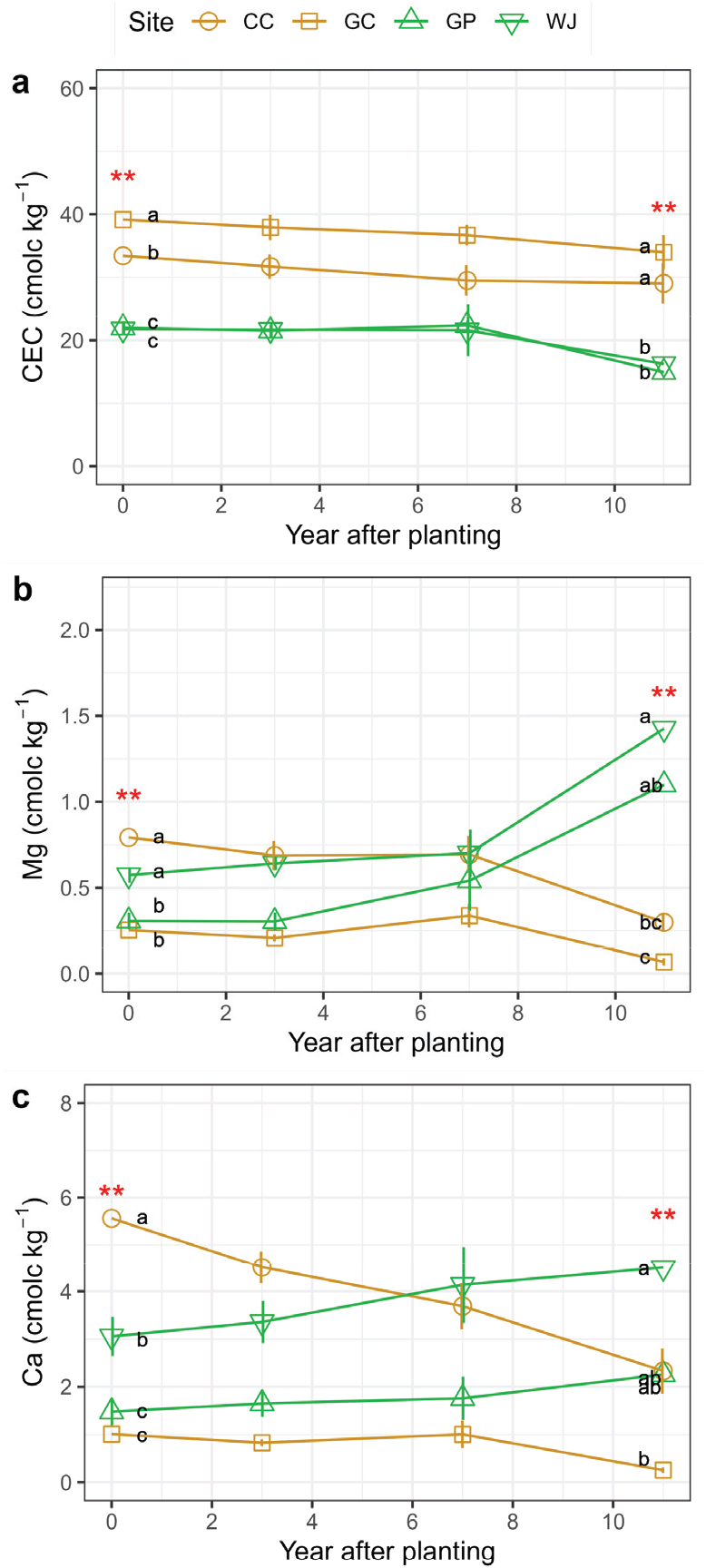
2010년 평균 양이온치환용량(cmolc kg−1)은 낙엽송 전생임분 김천(39.1±0.7)과 춘천(33.4±1.5) 조사지가 리기다소나무 전생임분 원주(21.8±0.9), 가평(22.0±1.0) 조사지에 비해 높게 나타났다[p<0.001, Figure 4(a)]. Yang et al.(2017)에 의한 선행연구에서는 CEC가 조림 후 3년차에도 식재 당시와 유사한 수준을 보였다고 보고한 바 있으나, 본 연구에서 11년간 모니터링 결과 조림 후 경과 시간에 따라 CEC는 지속적으로 감소하는 경향을 보였으며[p=0.002, Figure 4(a), Table 2], 조림 10년 경과 후 2021년 조사지 간 차이는 조림 초기와 유사하게 나타났다. 본 연구에서 조사지 간 CEC의 차이는 토양 총탄소, 즉 토양 유기물 함량과 유사한 경향을 보였으며[Figure 3(b)], 이러한 경향은 선행 연구들에서의 두 인자 간 상관성을 뒷받침한다(Johnson et al., 1997; Ranger et al., 2008). 한편 재조림 초기부터 CEC의 지속적인 감소는 조림 초기 토양으로 강우 유입 증가에 의한 침출에 기인할 수 있으며(Chen and Xu, 2005), 무기표층에서 감소된 CEC는 집적층(B층)에서의 증가로 나타날 수 있다(Johnson et al., 1997).
Figure 4.
Temporal changes in soil pH (a), total carbon (b), total nitrogen (c), and available phosphorous (P2O5-) over 11 years after planting Larix kaempferi trees in harvested L. kaempferi (yellow) and Pinus rigida (green) plantations. CC: Chuncheon, GC: Gimcheon, GP: Gapyeong, WJ: Wonju. Vertical bars indicate standard deviation of the mean (n=9~15). Asterisks indicate significant differences in the variables between the study sites by ANOVA (p < 0.05).
Download Original Figure
7) 치환성양이온
2010년 치환성양이온 중 Ca2+과 Mg2+ 평균 농도(cmolc kg−1)는 춘천(5.56±0.49, 0.79±0.10) 조사지에서 가장 높게 나타났으며, 원주(3.06±0.41, 0.57±0.04), 가평(1.48±0.29, 0.31±0.05), 김천(1.01±0.13, 0.25±0.04) 순으로 나타났다[Figure 4(b), 4(c), p<0.001]. K+과 Na+ 농도는 조사지 간 차이를 보이는 경우도 있었으나, 비교적 낮은 농도 범위 내에서 뚜렷한 경향성을 보이지는 않았다(Table 2, Table S2). 또한 전생임분과 시간 경과가 치환성양이온에 미치는 영향도 조사지별로 다르게 나타나 전반적인 경향성을 확인하기는 어려웠다. 특히 춘천과 김천은 조림 초기에 비해 2021년 Ca2+와 Mg2+ 농도가 50% 이하로 감소하는 경향을 보였으나, 원주와 가평에서는 2021년에 Ca2+ 농도가 약 50% 증가, Mg2+는 약 250% 이상 증가하는 등 반대의 경향을 보였다[Figure 4(b)]. Farahnak et al.(2022)은 C. obtusa 조림지에서 모두베기가 수간류로부터 기인하는 수소(H+)이온 유입을 감소시켰고, 이로 인해 수소이온과 양이온치환을 통한 2가 양이온의 손실이 줄었기 때문에 토양 내 Ca2+, Mg2+ 농도가 증가했음을 보고한 바 있다. 한편 Povilaitie et al. (2022)은 P. sylvestris 벌채 후 재조림지에서 조림 3년차에 Ca2+가 가장 높게 나타났으나, Mg2+는 벌채 및 조림 경과년수에 따라 유의미한 차이를 보이지 않았다.
2. 근원경 및 수고 생장
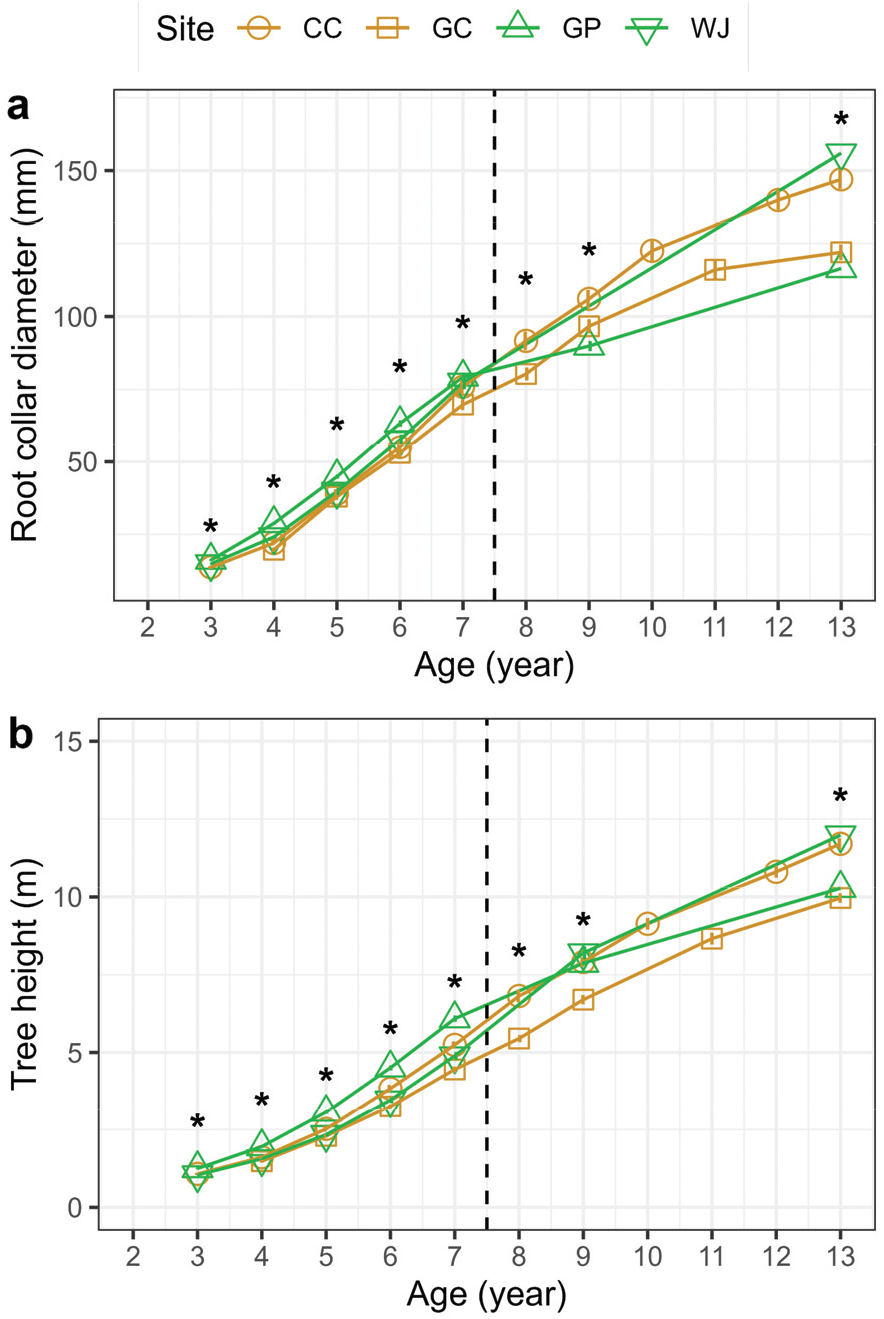
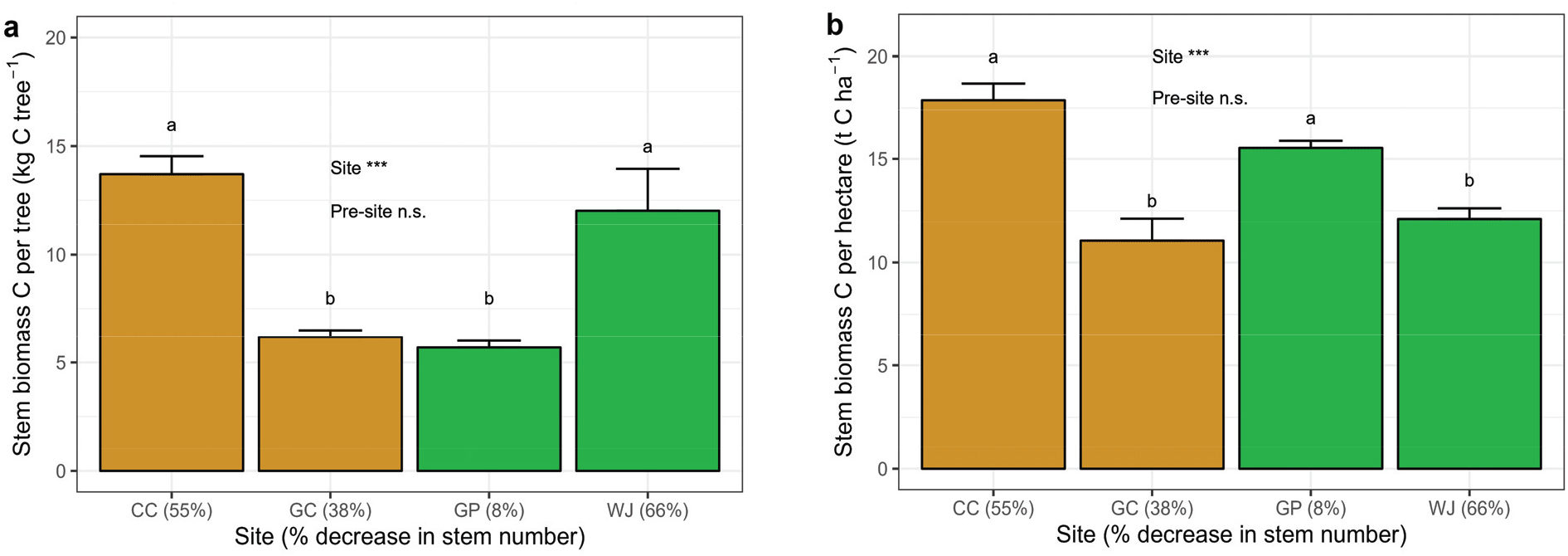
춘천, 김천, 원주, 가평 조사지 내 낙엽송 조림목의 근원경과 수고는 조사지 간 통계적 차이를 보였으나(p<0.001), 전생임분의 영향은 나타나지 않았다(Table 2, Figure 5).7년생 낙엽송 조림목의 근원경과 수고는 가평 조림지(79.2± 1.5 mm, 6.09±1.0 m)가 다른 조림지에 비해 높게 나타났다(Table S1). 한편 13년생 조림목의 근원경과 수고는 원주(156.0±3.5, 11.98±0.20)와 김천(147.0±3.3, 11.70±0.17) 조림지에서 가장 높게 나타나(p<0.001), 조림 초기와는 다른 생장 차이를 나타났다. 이는 조림목 7년생 이후 어린나무가꾸기 작업으로 조절된 조사지별 임분밀도, 즉 생육공간의 차이에 의한 영향을 크게 받았기 때문이다(Figure 5). 특히 임분밀도가 크게 감소한 원주(1,028본 ha-1)와 김천(1,359본 ha-1) 조림지에서 근원경 및 수고 생장이 크게 증가하였다. 조사지별 임분밀도 차이로 인해 토양 물리·화학적 특성이 생장에 미칠 수 있는 영향을 구명하기 어려웠으며, 향후 자연고사 및 어린나무가꾸기로 인한 고사유기물 발생에 따른 토양환경 변화와 생장 변화에 대한 추가적인 연구가 필요할 것으로 보인다(Kim et al., 2012; Liu et al., 2017).
Figure 5.
The root collar diameter (mm) (a), height (cm) (b) of Larix kaempferi trees planted in harvested L. kaempferi (yellow) and Pinus rigida (green) plantations over 11 years after planting. CC: Chuncheon, GC: Gimcheon, GP: Gapyeong, WJ: Wonju. Vertical bars indicate standard deviation of the mean (n=90). Asterisks indicate significant differences in the variables between the treatments by ANOVA (p < 0.001). Vertical line indicates the year of forest tending work.
Download Original Figure
3. 전생임분과 토양 물리·화학적 특성 변화
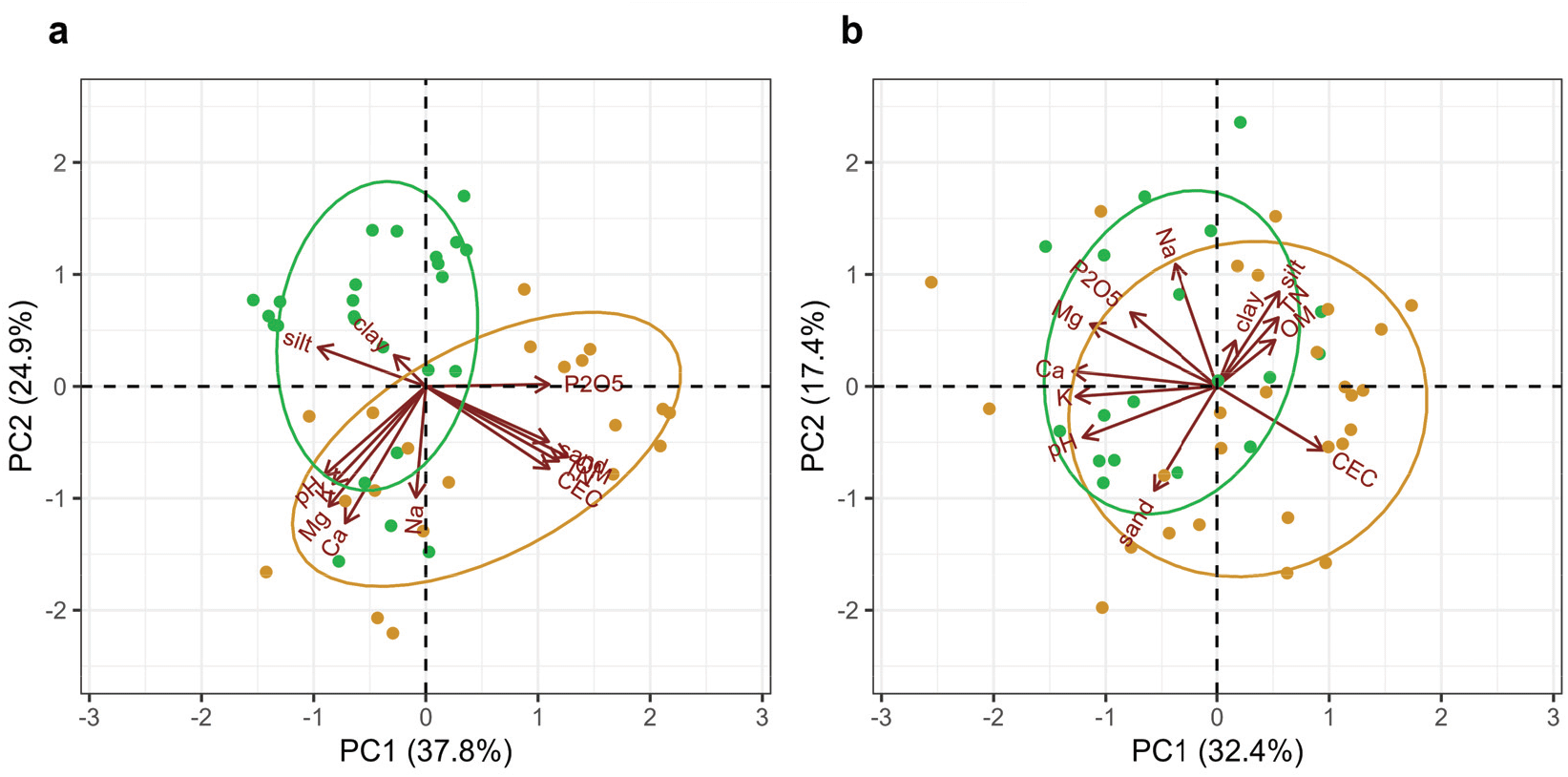
2010년 조림 초기 토양의 물리 · 화학적 특성들은 낙엽송 벌채지와 리기다소나무 벌채지 간에 전반적인 토양 특성의 차이를 보였다[Figure 6(a)]. 특히 모래함량, 총탄소 및 전질소 농도, 유효태인산 농도 등은 전생임분에 따른 차이를 보였다(Table 2). 그러나 본 연구는 각 지역별로 두 전생임분을 포함하지 않기 때문에, 지역과 전생임분이 토양 특성에 미치는 영향을 분리하는 데 한계가 있다. 한편 전생임분 따른 조림목의 생장 차이는 나타나지 않았다(Table 2). Yang et al.(2018)은 낙엽송 벌채지 내 식재된 낙엽송 조림목의 초기 생장에 유효태인산이 주요 영향인자 중 하나임을 보고한 바 있으나, 본 연구에서는 유효태인산과 전질소농도가 상대적으로 낮은 리기다소나무 전생임분(가평)에 조림된 낙엽송의 수고 및 근원경이 가장 크게 나타났다(Table S1). 이러한 결과는 향후 리기다소나무 벌채지에서 낙엽송으로의 수종갱신을 통한 재조림 성과도 기대할 수 있음을 시사한다. 단 어린나무가꾸기를 통해 임분밀도의 적절한 조절이 필요할 것으로 보인다(Figure S2). 한편 낙엽송 조림 후 10년이 경과한 2021년에는 조사지 간 토양 특성의 차이가 총탄소, 전질소 농도 등을 제외하고 유사하게 나타났으며[Figure 6(b)], 이러한 결과는 수종이 토양 특성 변화에 잠재적으로 영향을 미칠 수 있음을 시사한다(Binkley and Menyailo, 2005). 특히 수종 간 낙엽과 세근의 양적(생산량), 질적(질소농도, 탄질율 등) 차이는 토양미생물, 유기물 분해속도, 양분순환 등 토양의 생물·물리·화학적 특성 변화에 영향을 미칠 수 있다(Ayres et al., 2009). 수종이 토양 특성에 미치는 영향은 장기간에 걸쳐 나타나지만(Augusto et al., 2002), 속성수 조림지의 경우 표층에서부터 수종에 의한 토양 특성 변화를 기대할 수 있다(Russell et al., 2007). 낙엽송의 경우 리기다소나무에 비해 낙엽 내 질소 농도가 높고 분해율도 빠르게 나타나(Kim et al., 2005; 2010), 상록침엽수 조림수종에 비해 토양 특성 변화가 빠르게 나타날 수도 있을 것으로 보인다. 그러나 전생임분과 재조림 수종이 토양 특성에 미치는 영향을 보다 명확하게 구명하기 위해서는 유사한 입지환경에 조성된 다양한 수종의 조림지에서 장기적인 토양 특성 모니터링 연구가 필요하다.
Figure 6.
Principle component analysis (PCA) biplots of the soil variables for the planting year (2010) (a) and 11 years after planting larch seedlings (2021) (b) in the harvested larch (yellow) and pine (green) stands.
Download Original Figure
결 론
본 연구에서 낙엽송 재조림지의 인 가용성은 조림 초기부터 지속적으로 낮은 수준으로 나타나, 이는 조림목의 초기 생장 및 조림 성과 향상을 위해 산림 토양 양분 중 인 관리의 중요성을 시사한다. 또한 낙엽송 조림 후 10년이 경과한 2021년에는 조림 초기 지역 간 토양 특성의 차이가 낙엽송 조림으로 인해 점차 유사한 토양 특성으로 변했음을 보여준다. 특히 재조림 이후 주요하게 변화한 토양 인자는 모래함량, 총탄소 및 전질소 농도, 유효태인산 농도 등이다. 또한 조림 초기 유기물 및 양분 농도의 감소를 고려할 때, 모두베기 이후 토양 양분 용탈을 최소화하기 위해 조기 재조림을 통해 토양이 노출되는 기간을 최소화하고 조림목에 의한 가용성 양분의 흡수율을 높이는 것이 필요하다. 본 연구에서 벌채 후 재조림지의 장기적인 토양 물리·화학적 특성 변화는 낙엽송 재조림지의 양분 관리를 위해 유용한 정보를 제공한다.
감사의 글
본 연구는 2021년도 강원대학교 대학회계 학술연구조성비 및 국립산림과학원(SC0500-2020-01), 한국임업진흥원(2022460A00-2324-0201) 연구 지원으로 수행되었음.
References
Augusto, L., Ranger, J. and Binkley, D. 2002. Impact of several common tree species of European temperate forests on soil fertility. Annals of Forest Science 59: 233-253.


Ayres, E., Steltzer, H., Berg, S., Wallenstein, M.D., Simmons, B.L. and Wall, D.H. 2009. Tree species traits influence soil physical, chemical, and biological properties in high elevation forests. PLoS ONE 4(6): e5964.


Baek, K., Lee, E.J., Choi, H., Cho, M., Choi, Y. and Han, S. 2022. Impact on soil physical properties related to a high mechanization level in the row thinning of a Korean pine stand. Land 11, 329.
Binkley, D. and Menyailo, O. (Eds.) 2005. Tree species effects on soils: Implications for Global Change: Proceedings of the NATO Advanced Research Workshop on Trees and Soil Interactions, Implications to Global Climate Change, August 2004, Krasnoyarsk, Russia (Vol. 55). Springer Science & Business Media.
Caldeira, A., Krushe, A.V., Mareschal, L., da Silva, P., Nouvellon, Y., Campoe, O., Stape, J.L., Montebelo, A., Formaglio, G., le Maire, G., Guillemot, J., Ranger, J. and Laclau, J.P. 2023. Low nutrient losses by deep leaching after clearcutting and replanting Eucalyptus plantations in Brazil. Forest Ecology and Management 534: 120866.
Cesonienè, L., Daubaras, R., Tamutis, V., Kaskonienè, V., Kaskonas, P., Stakènas, R. and Zych, M. 2019. Effect of clear-cutting on the understory vegetation, soil and diversity of litter beetles in Scots pine-dominated forest. Journal of Sustainable Forestry 38: 791-808.
Chen, C.R. and Xu, Z.H. 2005. Soil carbon and nitrogen pools and microbial properties in a 6-year-old slash pine plantation of subtropical Australia: impacts of harvest residue management. Forest Ecology and Management 206(1-3): 237-247.
Farahnak, M., Mitsuyasu, K., Ide, J., Chiwa, M., Enoki, T., Jeong, S., Otsuki, K., Shimizu, K. and Kume, A. 2022. Soil pH and divalent cations after clear-cutting on a Japanese cypress plantation. Journal of Forest Research 27(5): 363-370.
Grønflaten, L.K., Steinnes, E. and Örlander, G. 2008. Effect of conventional and whole-tree clear-cutting on concentrations of some micronutrients in coniferous forest soil and plants. Forestry Studies 48: 5-16.
Han, S.K., Lee, K.C., Oh, J.H., Mun, H.S., Chio, Y.S. and Choi, B.K. 2019. Characteristics of soil disturbance caused by passages of harvester and forwarder in cut-to-length harvesting operations. Journal of Korean Society of Forest Science 108(1): 67-76. (in Korean with English abstract)
Hedwall, P.O., Grip, H., Linder, S., Lövdahl, L., Nilsson, U. and Bergh, J. 2013. Effects of clear-cutting and slash removal on soil water chemistry and forest floor vegetation in a nutrient optimised Norway spruce stand. Silva Fennica 47: 16.
Johnson, C.E., Romanowicz, R.B. and Siccama, T.G. 1997. Conservation of exchangeable cations after clear-cutting of a northern hardwood forest. Canadian Journal of Forest Research 27: 859-868.
Jussy, J.H., Ranger, J., Bienaimè, S. and Dambrine, E. 2004. Effects of a clear-cut on the in situ nitrogen mineralisation and the nitrogen cycle in a 67-year old Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) plantation. Annals of Forest Science 61: 397-408.
Kim, C., Koo, K.-S. and Byun, J.-K. 2005. Litterfall and nutrient dynamics in pine (
Pinus rigida) and larch (Larix leptolepis) plantations. Journal of Korean Forest Society 94(5): 302-306.
Kim, C., Son, Y., Lee, W.K., Jeong, J. and Noh, N.J. 2009. Influence of forest tending works on carbon distribution and cycling in a Pinus densiflora S. et Z. stand in Korea. Forest Ecology and Management 257(5): 1420-1426.
Kim, C., Son, Y., Lee, W.K., Jeong, J., Noh, N.J. and Kim, S.R. 2012. Litter decomposition and nutrient dynamics following forest tending (Soopkakkugi) works in a Pinus densiflora stand. Forest Science and Technology 8(2): 99-104.
Korea Forest Service. 2022. Statistical Yearbook of Forestry. Korea Forest Service, Korea. pp. 455. (in Korean)
Lajtha, K., Bowden, R.D. and Nadelhoffer, K. 2014. Litter and root manipulations provide insights into soil organic matter dynamics and stability. Soil Science Society of America Journal 78: S261-S269.
Lefebvre, D., Williams, A.G., Kirk, G.J.D., Burgess, P.J., Meersmans, J., Silman, M.R., Román-Daňobeytia, F., Farfan, J. and Smith, P. 2021. Assessing the carbon capture potential of a reforestation project. Scientific Report 11: 19907.
Liu, Y., Ding, F, Cui, Y., Xie, T., Ma, H. and Zhao, W. 2017. Effects of tending on carbon storage in the ecosystems of young Chinese fir plantations at the middle region of Guizhou Province, southwetern China. Journal of Beijing Forestry University 39(1): 27-33. (In Chinese with English abstract)
Nazari, M., Arthur, E., Lamandé, M., Keller, T., Bilyera, N. and Bickel, S. 2023. A meta-analysis of soil susceptibility to machinery-induced compaction in forest ecosystems across global climatic zones. Current Forestry Reports 9(5): 370-381.
Pennock, D.J. and van Kessel, C. 1997. Clear-cut forest harvest impacts on soil quality indicators in the mixedwood forest of Saskatchewan, Canada. Geoderma 75(1-2): 13-32.
Picchio, R., Mederski, P.S. and Tavankar, F. 2020. How and how much, do harvesting activites affect forest soil, regeneration and stands? Current Forestry Reports 6(2): 115-128.
Piirainen, S., Finér, L., Mannerkoski, H. and Starr, M. 2007. Carbon, nitrogen and phosphorus leaching after site preparation at a boreal forest clear-cut area. Forest Ecology and Management 243(1): 10-18.
Povilaitiené, A., Gedminas, A., Varnagiryté-Kabašinskiené, I., Marčiulyniené, D., Marčiulynas, A., Lynikienè, J., Mishcherikova, V. and Menkis, A. 2022. Changes in chemical properties and fungal communities of mineral soil after clear-cutting and reforestation of Scots pine (Pinus sylvestris L.) sites. Forests 13(11): 1780.
Ranger, J., Bonnaud, P., Bouriaud, O., Gelhaye, D. and Picard, J-F. 2008. Effects of the clear-cutting of a Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) plantation on chemical soil fertilty. Annals of Forest Science 65: 303.
Rodionov, A., Bauke, S.L., von Sperber, G., Hoeschen, C., Kandeler, E., Kruse, J., Lewandowski, H., Marhan, S., Mueller, C.W., Simon, M., Tamburini, F., Uhlig, D., von Blanckenburg, F., Lang, F. and Amelung, W. 2020. Biogeochemical cycling of phosphorus in subsoils of temperate forest ecosystems. Biogeochemistry 150(3): 313-328.
Russell, A.E., Raich, J.W., Valverde-Barrantes, O.J. and Fisher, R.F. 2007. Tree species effects on soil properties in experimental plantations in tropical moist forest. Soil Science Society of America Journal 71(4): 1389-1397.
Schweier, J., Magagnotti, N., Labelle, E.R. and Athanassiadis, D. 2019. Sustainability impact assessment of forest operations: a review. Current Forestry Reports 5(3): 101-113.
Seidel, F., Lopez, C.M.L., Bonifacio, E., Kurokawa, H., Yamanaka, T. and Celi, L. 2022. Seasonal phosphorus and nitrogen cycling in four Japanese cool-temperate forest species. Plant and Soil 472(1-2): 391-406.
Smith, C.T., Driggs, R.D., Stupak, I., Preece, C., Rezai-Stevens, A., Barusco, B., Roth, B.E., Fernandez, I.J. and Simpson, M.J. 2022. Effects of whole-tree and stem-only clearcutting on forest floor and soil carbon and nutrients in a balsam fir (Abies balsamea (L.) Mill.) and red spruce (Picea rubens Sarg.) dominated ecosystem. Forest Ecosystem and Management 519: 120325.
Son, Y. and Gower, S.T. 1991. Aboveground nitrogen and phosphorus use by five plantation-grown trees with different leaf longevities. Biogeochemistry 14(3): 167-191.
Yang, A-R. and Cho, M.S. 2019. The growth performances and soil properties of planted Zelkova serrata trees according to fertilization in harvested
Pinus rigida plantation over 6 years after planting. Journal of Korean Society of Forest Science 108(1): 29-39 (In Korean with English abstract).
Yang, A.-R., Jeong, J.Y. and Cho, M.S. 2017. The early growth characteristics of
Larix kaempferi trees planted in harvested
Larix kaempferi plantations. Journal of Korean Forest Society 106(1): 10-18 (In Korean with English abstract).
Yang. A.-R., Jeong, J.Y. and Cho, M.S. 2018. Site and soil factors affecting early growth of
Larix kaempferi trees planted in harvested
Larix kaempferi plantations. Journal of Korean Forest Science 107(1): 35-42 (In Korean with English abstract).
Supplementary Tables and Figures
Table S1.
The root collar diameter (RCD), height, diameter at breast height (DBH), basal area, stem volume and biomass, and total carbon stock of Larix kaempferi trees planted in harvested L. kaempferi and Pinus rigida plantations.
| Year |
Pre-stand |
Site |
RCD (mm) |
Height (m) |
DBH (em) |
Basal area (m2 ha−1) |
Stem volume (cm3 tree−1) |
Stem biomass (kg tree−1) |
Stem biomass (kg ha−1) |
Total C in tree (t C ha−1) |
2015
(7-year-old) |
Larix kaempferi
|
CC |
76.0 (3.3)ab |
5.25 (0.16)b |
- |
- |
17,813 (l,681)a |
8.1 (0.8)a |
- |
- |
| GC |
69.8 (1.9)b |
4.45 (0.09)c |
- |
- |
12,305 (822)a |
5.6 (0.4)a |
- |
- |
|
Pinus rigida
|
WJ |
77.4 (2.3)a |
4.89 (0.13)b |
- |
- |
17,165 (l,310)a |
7.8 (0.6)a |
- |
- |
| GP |
79.2 (1.5)a |
6.09 (0.10)a |
- |
- |
20,929 (l,038)a |
9.5 (0.5)a |
- |
- |
2021
(13-year-old) |
Larix kaempferi
|
CC |
147.0 (3.3)a |
11.70 (0.17)a |
12.7 (2.6)a |
17.9(0.6)ab |
- |
27.4 (1.7)a |
35.8 (1.6)a |
71.9 (1.0)a |
| GC |
122.0 (2.7)b |
9.97 (0.15)b |
9.6 (0.2)b |
14.1(0.8)bc |
- |
12.4 (0.6)b |
22.1 (2.1)b |
45.4 (3.2)c |
|
Pinus rigida
|
WJ |
156.0 (3.5)a |
11.98 (0.20)a |
12.2 (0.2)a |
13.8(1.4)c |
- |
24.0 (3.9)a |
24.2 (1.0)b |
46.0 (1.7)c |
| GP |
116.5 (2.1)b |
10.28 (0.1 l)b |
9.2 (0.2)b |
19.6(0.4)a |
- |
11.4 (0.6)b |
31.1 (0.7)a |
60.9 (1.2)b |
Download Excel Table
Table S2.
Soil physio-chemical properties of the Larix kaempferi stands planted in harvested L. kaempferi and Pinus rigida plantations.
| Year |
Pre-stand |
Site |
Sand (%) |
Silt (%) |
Clay (%) |
pH |
TN (g kg−1) |
TC (%) |
P2O5 (mg kg−1) |
CEC (cmol+ kg−1) |
K+ (cmol+ kg−1) |
Ca2+ (cmol+ kg−1) |
Na+ (cmol+ kg−1) |
Mg2+ (cmol+ kg−1) |
1st planation
(2010) |
Larix kaempferi
|
CC |
63.8(1.3)b |
28.7(2.0)a |
7.4(1.0)bc |
5.24(0.09)a |
3.85(0.28)b |
10.5(0.6)b |
11.0(0.6)b |
33.4(1.51)b |
0.21(0.02)a |
5.56(0.49)a |
0.058(0.01l)a |
0.79(0.10)a |
| GC |
80.3(4.8)a |
14.8(5.3)b |
4.9(1.0)c |
4.59(0.05)b |
5.30(0.21)a |
15.9(0.8)a |
32.1(6.4)a |
39.1(0.7)a |
0.13(0.01)b |
1.01(0.13)c |
0.034(0.005)a |
0.25(0.04)b |
|
Pinus rigida
|
WJ |
60.1(1.7)b |
24.5(2.7)ab |
15.3(2.0)a |
5.16(0.06)a |
3.11(0.33)b |
8.3(1.0)b |
13.2(1.6)b |
21.8(0.9)c |
0.21(0.01)a |
3.06(0.4 l)b |
0.045(0.008)a |
0.57(0.04)a |
| GP |
58.3(2.7)b |
28.6(2.7)a |
13.1(1.7)ab |
4.52(0.08)b |
3.29(0,39)b |
7.8(0.4)b |
14.8(2.4)b |
22.0(1.0)c |
0.13(0.01)b |
1.48(0.29)c |
0.031(0.003)a |
0.31(0.05)b |
11st plantation
(2021) |
Larix kaempferi
|
CC |
44.1(10.1)a |
41.0(6.8)a |
14.9(3.4)a |
5.07(0.1l)a |
3,28(0.40)b |
6.5(1.l)b |
5.9(3.1)a |
29.0(3.2)a |
0.32(0.04)a |
2,33(0.47)ab |
0.034(0.002)a |
0.30(0.06)bc |
| GC |
46.9(2.6)a |
42.7(1.3)a |
10.4(1.3)a |
4.67(0.04)a |
5.81(0.61)a |
11.3(1.2)a |
2.7(0.l)a |
34.0(2.8)a |
0.08(0.0 l)b |
0.25(0.06)b |
0.058(0.010)a |
0.07(0.02)c |
|
Pinus rigida
|
WJ |
52.7(1.0)a |
35.3(1.5)a |
12.0(1.0)a |
4.87(0.16)a |
3.07(0.27)b |
7.0(0.6)b |
11.4(0.6)a |
16.2(0.9)b |
0.28(0.05)a |
4.51(1.30)a |
0.153(0.009)b |
1.43(0.34)a |
| GP |
52.7(1.0)a |
47.4(2.0)a |
16.6(1.4)a |
4.56(0.19)a |
3.80(0.23)b |
5.8(0.4)b |
11.2(2.7)a |
14.9(0.9)b |
0.20(0.02)ab |
2,26(0.33)ab |
0.153(0.003)b |
1.10(0.26)ab |
Download Excel Table
Figure S1.
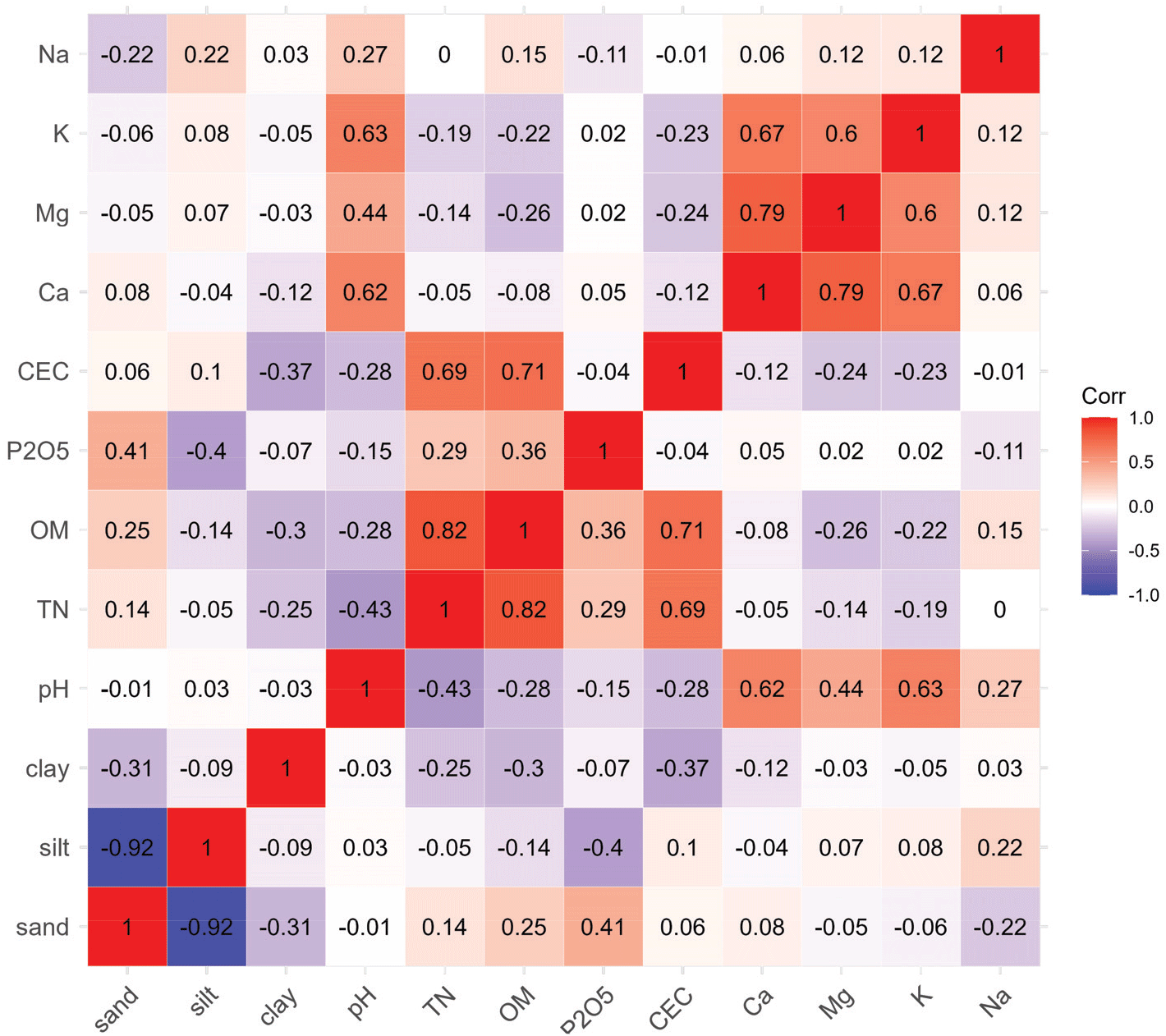
Heat map for the correlations between the soil properties. The correlation coefficients between the variables were shown in each cell. The map was produced by using ‘ggcorrplot’ package in R.
Download Original Figure
Figure S2.
Stem biomass carbon per tree (a) and stem biomass carbon per hectare (b) estimated in 2021 for Larix kaempferi plantations. CC: Chuncheon, GC: Gimcheon, GP: Gapyeong, WJ: Wonju. The number in the parenthesis indicates percentage decrease in stem number by forest tending works. Vertical bars indicate standard deviation of the mean (n=3). Different letter indicates significant differences in the variables between the treatments by ANOVA (p < 0.05).
Download Original Figure