Article
친환경 벌채(retention harvesting)에 따른 식물 및 토양, 곤충, 조류·포유류 군집의 초기 변화: 국내 두 가지 사례 연구
이연우1,
손지혜1,
최지원1,
이혁재1,
황수경1,
전수민1,
심형석2,3,
김정남1,
배양섭1,
엄태원2,
이종구1,*
Early Ecological Responses of Vegetation, Soil, Insect, Bird, and Mammal Communities to Retention Harvesting: Two Case Studies in South Korea
Yeon Woo Lee1,
Jihye Son1,
Jiwon Choi1,
Hyeok Jae Lee1,
Soo Kyeong Hwang1,
Sumin Jeon1,
Hyung Seok Sim2,3,
Jeong–Nam Kim1,
Yang-Seop Bae1,
Tae Won Um2,
Jong koo Lee1,*
Author Information & Copyright ▼
1Division of Life Sciences, Incheon National University, Incheon 22012, Korea
2Forest Ecosystem Restoration, Institute of Hutan, Seoul 06336, Korea
3Department of Forest Resources, Kookmin University, Seoul 02707, Korea
© Copyright 2025 Korean Society of Forest Science. This is an Open-Access article distributed under the terms of the
Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits
unrestricted non-commercial use, distribution, and reproduction in any
medium, provided the original work is properly cited.
Received: Sep 23, 2025; Revised: Nov 21, 2025; Accepted: Nov 24, 2025
Published Online: Dec 31, 2025
요 약
수종 갱신 및 수확을 위한 벌채 산림 시업은 산림생태계 구성요소인 곤충, 식물, 조류 및 포유류 군집의 변화를 발생시키기 때문에, 이 변화의 과정을 파악하는 것은 산림생태계 균형 유지 및 생태계 안정성 확보를 위해 매우 중요하다. 그러나 국내에 산림 벌채 시업의 전과 후의 생물 군집의 직접적인 변화를 비교한 사례는 매우 부족한 실정이다. 본 연구는 홍천과 충주에서 친환경 벌채에 따른 생태계 변화를 모니터링하였다. 2022년 벌채가 진행되기 전 조사지의 토양, 식생, 곤충, 포유류, 조류 군집에 대한 사전 조사를 실시했으며, 벌채가 진행된 후, 1년과 2년 경과 시점에 사후조사를 수행하였다. 또한, 연구지역내 벌채가 진행된 벌채구와 일부 산림을 남긴 잔존구, 벌채를 진행하지 않은 비벌채 대조구의 변화상을 비교하였다. 그 결과, 벌채 시업으로 인해 일시적으로 토양의 구성성분이 변화하였고, 식생, 곤충, 포유류, 조류 군집의 종다양도 지수가 감소하였으나, 시간이 지나면서 증가하는 경향을 보였다. 식생의 경우 벌채구, 잔존구 모두 시간에 따라 증가하였다. 잔존구에서 특히 크게 증가하였다. 곤충의 경우도 마찬가지로 잔존구에서 크게 증가하였으며, 벌채구 또한 큰 증가폭이 나타났다. 조류와 포유류의 경우 벌채에 따라 종마다 다른 경향을 보이는 것으로 판단된다. 본 연구는 친환경 벌채가 산림 생물군집에 미치는 직접적 영향과 초기 회복 과정을 보여주는 중요한 사례를 제시한다. 그러나 산림 시업의 효과는 장기적으로 산림생태계에 영향을 미칠 수 있기 때문에, 정확한 회복 과정을 파악하기 위해서는 지속적인 모니터링이 필요하다.
Abstract
Forest harvesting for stand regeneration and timber production inevitably alters insect, plant, avian, and mammalian communities—key components of forest ecosystems. Therefore, understanding these successional changes is crucial for maintaining ecological balance and ensuring ecosystem stability. Nevertheless, there is a scarcity of direct comparative studies evaluating preharvest and postharvest biotic changes in Korea. Hence, this study was conducted to investigate the ecological responses to environmentally sustainable retention harvesting practices in Hongcheon and Chungju. Baseline surveys of soil, vegetation, insect, mammalian, and avian communities were performed before harvesting in 2022, followed by postharvest surveys 1 and 2 years after logging. Ecological changes were compared across the following three plot types: harvested plots, residual plots with partial tree retention, and unharvested control plots. Harvesting temporarily altered the soil composition and caused short-term decreases in species diversity across multiple taxa; however, diversity indices generally increased over time. Vegetation diversity increased in both harvested and residual plots, with particularly remarkable gains in residual plots. Insect diversity also increased considerably, especially where partial retention was applied. For avian and mammalian communities, the impact of harvesting appeared to vary according to the specific species. These findings provide a direct case example of the immediate effects of eco-friendly logging on forest ecosystems. Nonetheless, considering that forest management practices can exert long-term effects on these ecosystems, continuous monitoring is necessary to accurately characterize the recovery process.
Keywords: ecosystem-based management; green-tree retention system; aggregated retention; biodiversity; adaptive forest management
서 론
1970년대 이후 추진된 전국적인 산림녹화 및 조림사업의 성공으로 지난 50여 년간 국내 산림의 임목은 크게 축적되었다(Tak et al., 2007; Ko et al., 2015). 이후로, 국내 산림은 목재 섬유자원의 활용(Jang and Youn, 2021), 산림 연령 분포의 균형 개선(Kim et al., 2021), 미래 수요 대응 및 기후변화 적응을 위한 새로운 수종 생산(Bang, 2017), 순환형 바이오경제 촉진(KFS, 2017) 등 다양한 목적을 위해 관리되고 있다. 또한, 적절한 산림 관리는 탄소 흡수 기능 증진(An et al., 2019), 산불 위험 감소(Lee, 2005), 야생동물 서식지 개선, 생물다양성 증진(Schwenk et al., 2012; Carpentier et al., 2017) 등 다양한 생태계서비스 향상에 기여할 수 있다.
산림관리에서 벌채는 여전히 중요한 논쟁의 대상이며, 이에 대한 반대도 적지 않다(Bliss, 2000). 벌채는 경관에 부정적인 영향을 미치며, 산림생태계에 바람직하지 않은 생태적 결과를 초래할 수 있다. 특히, 천이 과정을 교란하고 산림식생의 구조 및 산림 곤충과 동물의 서식지를 변화시킨다(Franklin and Donato, 2020). 이러한 우려와 더불어 벌채 이후 2차림 형성에 긴 시간이 소요된다는 점을 고려하여, 벌채량을 최소화하는 친환경 벌채 방안을 촉진하기 위한 정책이 시행되고 있다(Kim et al., 2016).
벌채의 부정적 환경 영향을 최소화하기 위한 방안으로 “친환경 벌채”가 제안되었다(Wang and Kim, 2022). 다양한 친환경 벌채 방식 중 ‘잔존벌채(retention harvesting)’는 전 세계적으로 널리 적용된다(Franklin et al., 1997). 이러한 벌채 방식은 산림생태계의 지속가능성을 증진시키고, 산림을 친환경적으로 관리할 수 있도록 한다(Tak et al., 2007; Park, 1995). 국내에서 5 ha 이상의 벌채지에서 잔존벌채 방식이 적용되며, 전체 면적의 최소 10% 또는 ha당 50본 이상의 수목을 남겨둠으로써 탄소격리, 수원 보호, 토양 침식 방비, 야생동물 서식지 보전 등 다양한 생태계 기능을 유지하도록 하고 있다(Kim et al., 2024). 전면벌채와 달리, 잔존벌채는 급격한 환경 변화를 유발하지 않으며, 효율적인 조림을 가능하게 한다(Wang and Kim, 2022). 전 세계적으로 해당 분야의 적용 및 연구가 활발히 진행되고 있음에도 불구하고(e.g. Vanha-Majamaa and Jalonen 2001; MacDonald and Fenniak 2007; Franklin and Donato 2020; Gustafsson et al., 2020; Akashi 2023), 국내에서는 산림 벌채가 산림 생물 다양성에 미치는 영향에 대한 연구는 여전히 부족하다(Kim et al., 2020a; Kim et al., 2020b; Kim et al., 2024).
본 연구는 친환경 벌채의 한 종류인 잔존벌채에 따른 산림생태계 구성 요소의 초기 반응을 규명하고자 하였다. 이를 통해 벌채가 산림생태계에 미치는 영향을 평가하고, 이러한 변화가 야기하는 환경변화와 더불어 먹이사슬 및 포식-피식 관계와 같은 상호작용에 어떠한 영향을 미치는가를 이해하는 데 기여하고자 한다. 본 연구의 결과는 벌채로 인한 식생, 토양, 곤충, 조류 및 포유류 간의 상호작용을 이해하기 위한 기초 자료를 제공할 것으로 기대된다.
재료 및 방법
1. 연구지역
1) 홍천
홍천 조사지는 강원도 홍천군(37°50’21” N, 127°57’50” E)에 위치한 친환경 벌채 대상지이며, 고도 393-672 m, 7-42°의 남서사면이다. 본 조사지는 잣나무(Pinus koraiensis)와 일본잎갈나무(Larix kaempferi) 조림지로 구성되어 있다. 상층 식생은 주로 잣나무가 우점하며, 신갈나무(Quercus mongolica), 굴참나무(Quercus variablilis), 물푸레나무(Fraxinus rhynchophylla) 등이 혼재한다. 평균 수고는 16-24 m에 달하며, 중간식생은 희소하였으나, 최근 실시된 간벌로 광환경이 개선되면서 산초나무(Zanthoxylum schinifolium)가 활발히 발달하였다. 지난 10년간 연평균기온은 11.8°C이며, 최저 기온과 최고 기온은 각각 6.4°C, 18.4°C, 연평균 강수량은 1,136.0 mm, 평균풍속 1.1 m/s 평균 상대습도는 67.3%로 나타났다.
2) 충주
충주 조사지는 충청북도 충주시(37°6’34”N, 127°56’48”E)에 위치한 친환경 벌채 대상지이며, 고도 151-220 m, 5-21°의 완만한 남서향 경사면이다. 상층식생은 상수리나무(Quercus acutissima), 굴참나무(Quercus. variabilis), 밤나무(Castanea crenata)가 우점하며, 중층 및 하층식생은 수관층에 의한 수광량 부족으로 인해 생장이 제한되었다. 지난 10년간 연평균기온은 12.4°C이며, 최저 기온과 최고 기온은 각각 6.9°C, 18.5°C, 연평균 강수량은 1,170.4 mm, 평균풍속 1.4 m/s, 평균 상대습도는 63.6%로 나타났다.
3) 친환경 벌채
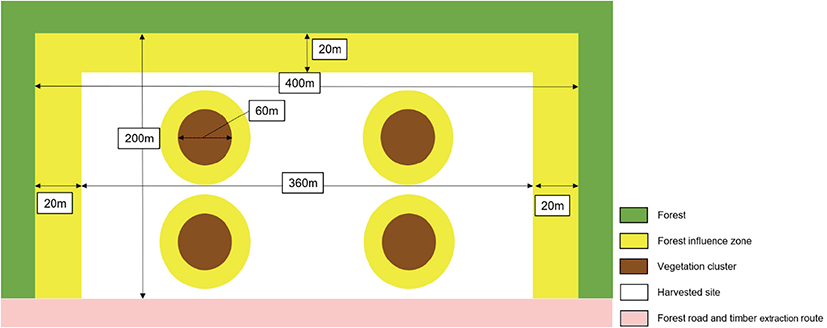
친환경 벌채는 잔존벌채로 진행되었으며, 홍천, 충주 두 조사지에서 2022년에 실시하였다. 친환경 벌채 이후 조사지는 벌채가 된 벌채구와 벌채가 되지 않은 잔존구가 형성되었다. 벌채 면적은 홍천 26.0 ha, 충주 14.5 ha였다(Figure 1). 각 조사구에서 약 8 ha의 임분을 벌채하였으며, 잔존구는 최소 폭 60 m 이상을 확보하였다(Figure 2). 잔존지는 평균 상층목 수고가 20 m가 되도록 유지하였다.
2. 시료 채취 및 분석
1) 식생 조사
두 조사지에서 식생 조사를 위해 20 m×20 m의 정방형 영구 조사구를 설치하였으며, 모든 조사구는 최소 50 m 이상의 간격을 두고 배치하였다. 홍천 조사지에서는 벌채 전 식생을 조사하기 위해 13개 조사구를 설치하였고, 벌채 영향을 분석하기 위해 총 10개 조사구를 추가로 설치하였다. 충주 조사지에서는 벌채 전후 식생 평가를 위하여 9개 조사구를 설치하였다. 모든 조사구는 벌채구와 잔존구에 고루 분포할 수 있게 설치하였다. 식생 조사는 흉고직경(DBH, Diameter at Breast Height) 2 cm 이상의 모든 수목을 대상으로 수종, 수고 및 흉고직경을 측정하였다. 흉고직경 2 cm 미만의 관목과 치수 등 하층식생은 Braun-Blanquet 방법(Braun-Blanquet, 1964)을 이용하여 조사하였다. 식생 조사는 2022년부터 2024년까지 매년 2회(5-6월, 9-10월)에 걸쳐 수행하였다.
2) 토양 조사
식생 조사를 수행할 때 토양의 화학적 특성 변화를 모니터링하기 위해 매년 1회(5-6월) 토양 시료를 채취하였다. 홍천 조사지에서는 13점, 충주 조사지에서는 9점의 시료를 잔존구에서 벌채구까지 분포하도록 채취하였다. 토양 채취 시 유기물층(O층)을 제거한 후, 지정된 토양 조사구에서 약 200 g의 토양을 소형 흙삽으로 채취하였다. 채취된 시료는 지퍼백에 밀봉하여 실험실로 운반하였으며, 총 유기물 함량, 총 질소(N) 함량, 유효인산(유효P2O5), 양이온치환용량(CEC, Cation Exchange Capacity), 토양 pH를 분석하였다.
3) 곤충 조사
곤충 조사는 2022년부터 2024년까지 진행하였다. 벌채구, 잔존구 이외에도 홍천 내 벌채가 진행되지 않은 산림을 임의로 지정하여 비벌채 대조구로 설정하였다. 각 조사지에는 10 m×10 m 크기의 고정 조사구를 벌채구, 잔존구, 대조구에 각각 설치하였다. 정량조사는 쓸어잡기법(sweeping)과 버킷라이트트랩(bucket light trap)을 이용하였다. 쓸어잡기법은 조사구당 30회씩 3반복으로 수행하였으며, 버킷라이트트랩은 조사구당 1개씩 설치하여 일몰 후 4시간동안 가동한 뒤 익일 아침에 회수하였다. 정성조사는 임의채집법(random collecting)과 털어잡기법(beating method)을 병행하였다. 임의채집법은 1 km 선조사 경로를 따라 수행하였으며, 털어잡기법은 선조사 경로 상의 수목과 관목을 흔들어 곤충을 채집하는 방식으로 수행하였다. 정성 채집은 조사자 2인이 각각 2시간 이상 실시하였다. 모든 채집은 5월에 1차 조사, 7월 9월, 10월 중 2차 조사를 진행하여 총 연 2회 수행하였다. 채집된 곤충들은 실험실로 운반하여 처리, 분류 및 동정을 거친 후 분석에 활용하였다. 분류, 동정과 국명 및 학명 체계는 NIBR(2022)를 참고하였다.
4) 포유류 및 조류 조사
포유류 조사와 조류 조사의 경우 비벌채 대조구를 추가로 설정하여, 벌채가 진행된 조사지와 비교하였다. 무인센서카메라의 경우 세분화하여 벌채구, 잔존구, 대조구 3곳을 비교하였다. 포유류 조사는 흔적조사와 무인센서카메라 조사를 병행하여 수행하였다. 흔적조사는 연구지역 내 1 km 조사 경로를 따라 이동하며 발자국, 배설물, 은신처, 먹이 흔적 등 포유류의 흔적을 관찰 및 동정하였다. 조사는 벌채 조사지와 대조구 각 계절 당 2회 실시하였다. 무인센서카메라는 동작 센서를 통해 움직임을 감지하여 사진 2장과 10초 영상 1개를 촬영하였으며, 24시간 모니터링 하였다. 동일 지점에서 30분 내 동일 종이 탐지된 경우 독립적이지 않은 것으로 가정하고 동일한 사건으로 판단하였다. 연구 지역마다 총 20대의 카메라를 설치하였으며, 벌채 조사지에 10대, 대조구에 10대를 설치하였다. 홍천 조사지의 경우 잔존구 6대, 벌채구 4대로 나눠 설치하였으며, 충주 조사지의 경우 잔존구, 벌채구 각 5대씩 설치하였다.
조류 조사는 조사지와 대조구 각 조사경로를 따라 이동하며 조사를 진행하는 선조사법을 이용하였다. 약 2 km/h의 속도로 1 km의 조사 경로를 걸었으며, 조사는 일출부터 일출 후 2시간 이내에 조사를 수행하였다. 쌍안경을 이용한 시각적 관찰과 울음소리를 통해 조류를 동정 및 기록하였다. 현장에서 동정이 되지 않는 경우 사진 촬영 또는 녹음 후 실험실에서 추가 동정하였다. 철새와 텃새 간의 잠재적 차이로 인한 계절적 변동을 파악하기 위해 계절별로 2회 조사를 실시하였다.
3. 데이터 분석
식생 조사에서 개체군별 중요치(IV)는 각 조사구에서 상대밀도(RD, 전체 개체수 중 해당 수종의 비율)와 상대피도(RC, 전체 피복 면적 대비 해당 수종의 점유 면적 비율)의 평균값으로 산출하였다. 종다양도의 경우 Shannon-Weaver(1949)의 지수(H’)를 산출하였다.
Ni는 i번째 종의 개체수, N은 전체 개체수, S는 종풍부도를 나타낸다.
종다양도지수의 변화량을 파악하기 위해 변동계수 CV(%)를 계산하였다(Abdi, 2010).
S는 표준편차, M은 평균을 나타낸다.
무인센서카메라 자료 분석에는 상대풍부도지수(Relative Abundance Index, RAI; O’Connell et al., 2011)를 사용하였다. RAI는 조사구별 총 관측 개체수(Number of independent detections)를 총 관측일(Toral camera trap days)로 나눈 후 100을 곱하여 산출하였으며, 100일 기준의 상대적 빈도를 나타낸다(Lamelas-López et al., 2020).
결 과
1. 산림식생
홍천 조사지의 식생은 주로 잣나무(Pinus koraiensis)림으로 구성되어 있으며, 대부분의 조사구에서 중요치(importance values)가 50% 이상을 차지하였다(Table 1). 신갈나무(Quercus mongolica)가 우점하는 조사구는 굴참나무(Quercus varialilis)와 혼생하였으며, 점차 침엽수림으로 확산되는 양상을 보였다. 벌채 이후 침엽수종의 중요치는 감소하였으나, 점차 침엽수림이 확산되는 양상을 보였다. 그러나 잔존구에서는 여전히 우점성을 유지하였다.
Table 1.
Temporal changes in vegetation composition for major species observed in the study sites before (2022) and after harvesting. The importance value (IV) was calculated as the averaging of relative density (RD) and relative coverage (RC).
| Site |
Species |
IV |
RD |
RC |
| 2022 |
2023 |
2024 |
2022 |
2023 |
2024 |
2022 |
2023 |
2024 |
| Hongcheon |
Pinus koraiensis
|
80.6 |
68.5 |
67.9 |
74.7 |
48.2 |
47.3 |
86.5 |
88.7 |
88.5 |
|
Larix kaempferi
|
24.2 |
41.6 |
37.1 |
15.8 |
19.8 |
14.7 |
32.6 |
63.4 |
59.4 |
|
Quercus mongolica
|
23.3 |
100.0 |
74.4 |
24.3 |
100.0 |
50.0 |
22.2 |
100.0 |
2.7 |
|
Lindera obtusiloba
|
- |
25.1 |
26.2 |
- |
48.1 |
49.7 |
- |
2.1 |
98.7 |
|
Quercus variabilis
|
17.3 |
41.7 |
16.4 |
17.2 |
23.1 |
18.6 |
17.4 |
60.4 |
22.0 |
| Chungju |
Quercus variabilis
|
42.6 |
45.7 |
56.5 |
38.1 |
30.7 |
43.0 |
47.2 |
60.7 |
70.1 |
|
Quercus aliena
|
17.5 |
27.6 |
24.2 |
17.9 |
37.0 |
32.3 |
17.1 |
18.2 |
16.1 |
|
Quercus acutissima
|
24.5 |
10.3 |
25.2 |
21.5 |
8.0 |
15.6 |
27.5 |
12.6 |
34.8 |
|
Castanea creanta
|
13.4 |
11.3 |
7.8 |
10.5 |
8.0 |
6.7 |
16.3 |
14.7 |
8.8 |
|
Zelkova serrata
|
7.0 |
14.1 |
15.6 |
11.9 |
25.0 |
26.7 |
1.9 |
3.2 |
4.6 |
Download Excel Table
충주 조사지에서는 참나무류(Quercus spp.)가 우점하였으며, 산벚나무(Prunus sargentii), 아까시나무(Robinia pseudoacacia), 말채나무(Cornus walteri), 느티나무(Zelkova serrata), 신나무(Acer tataricum) 등이 함께 출현하였다. 벌채 이후에도 참나무류는 우점성을 유지하였으며, 일부는 그 우점도가 증가하였다.
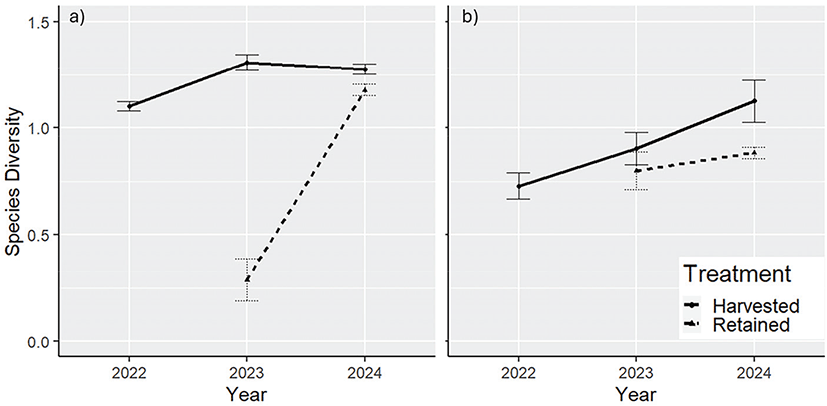
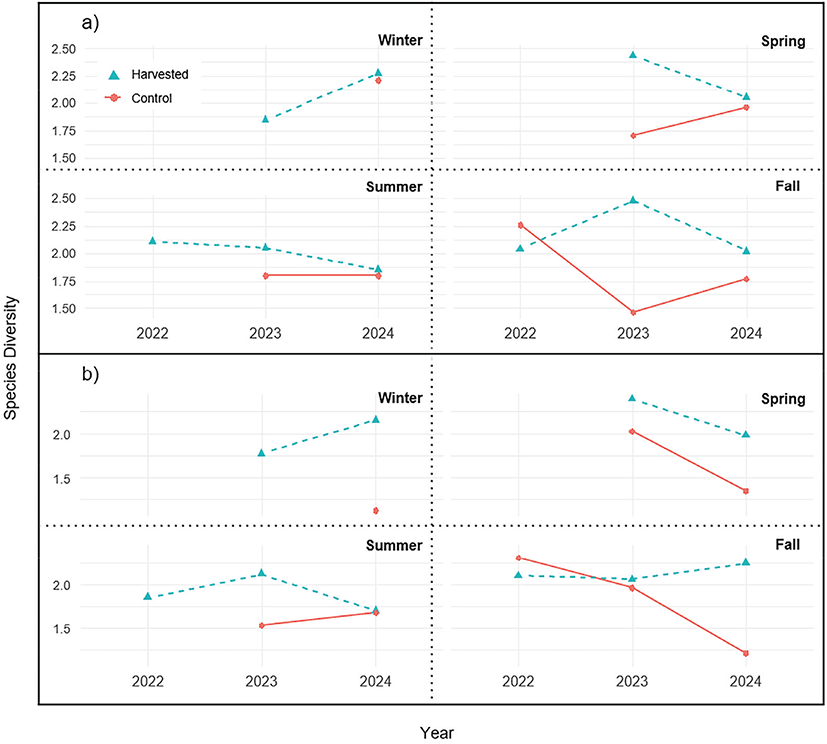
벌채 후 홍천과 충주 두 조사지 모두에서 종다양도가 증가하는 경향을 보인다(Figure 3). 홍천 벌채구의 초기 종다양도 지수는 낮게 나타났으나(2022년: 1.10±0.08), 이후 조사에서 증가하였다(2023년: 1.31±0.13, 2024년: 1.28±0.09). 홍천 벌채구의 종다양도의 변동계수 CV(%)는 9.07%로 나타났다. 잔존구의 경우 2023년 종다양도는 낮게 나타났으나(2023년: 0.29±0.31), 1년 후 종다양도는 증가하였다(2024년: 1.18±0.08). 홍천 잔존구의 종다양도의 변동계수 CV(%)는 86.01%로 나타났다. 충주 벌채구의 초기 종다양도 지수는 낮게 나타났으나(2022년: 0.73±0.19), 이후 조사에서 증가하였다(2023년: 0.90±0.23, 2024년 1.05±0.16). 충주 벌채구의 종다양도의 변동계수 CV(%)는 17.86%로 나타났다. 충주 잔존구의 경우 2023년 종다양도는 낮게 나타났으나(2023년: 0.79±0.26), 2년 후 종다양도는 증가하였다(2024년: 0.88±0.08). 충주 잔존구의 경우 CV(%)는 7.15%로 나타났다. 두 조사구 모두에서 종다양도가 지속적으로 증가하는 경향이 관찰되었다([Figure 3(b)]). 그러나 잔존구와 벌채 영향 구역의 종다양도 변화는 벌채구와 홍천 조사지에 비해 상대적으로 낮게 나타났다.
Figure 3.
Species diversity (Shannon-Weaver Index) changes in vegetation after retention harvesting for the a) Hongcheon and b) Chungju site.
Download Original Figure
2. 토양
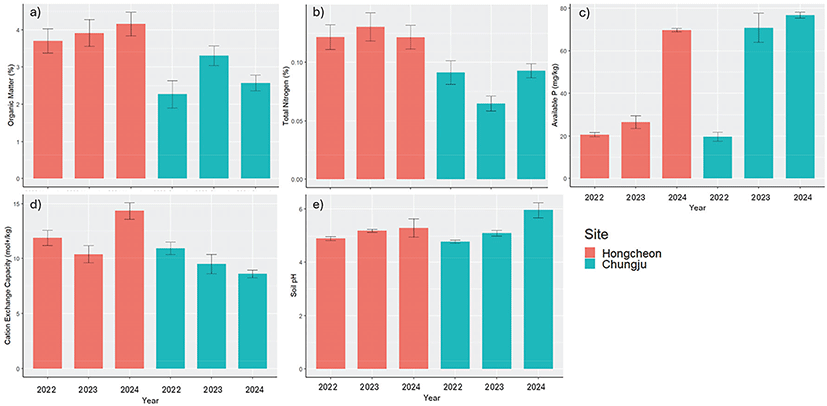
홍천 조사지에서는 토양 유기물 함량, 유효인산, 토양 pH가 증가하는 경향을 보였다(Figure 4). 반면 충주 조사지에서는 질소 함량이 벌채 2년 후 회복하여 벌채 이전 수준과 유사한 수준에 도달하는 것으로 나타났다. 두 조사지 모두에서 CEC는 감소하는 경향을 보였으며, 토양 pH는 벌채 이후 점진적인 증가하는 경향을 나타냈다.
Figure 4.
Temporal changes in soil chemical properties of study sites, including a) organic matter contents (%), b) total N (%), c) available phosphorus (P2O5), d) cation exchange capacity (CEC; mol+/kg), and e) soil pH.
Download Original Figure
3. 곤충
홍천 조사지에서는 2022년 10목 56과 93종, 2023년 12목 67과 157종, 2024년 11목 71과 190종이 관찰되었다. 친환경 벌채 후 2024년 기준 2022년도에 비해 1목 15과 97종, 2023년도에 비해 4과 33종이 증가하였다(Table 2).
Table 2.
Number of insect samples collected on the study sites.
| Site |
Order |
2022 |
2023 |
2024 |
Site |
Order |
2022 |
2023 |
2024 |
| Family |
Species |
Family |
Species |
Family |
Species |
Family |
Species |
Family |
Species |
Family |
Species |
| Hongcheon |
Lepidoptera |
13 |
34 |
22 |
83 |
16 |
92 |
Chungju |
Lepidoptera |
12 |
50 |
18 |
64 |
21 |
163 |
| Phasmida |
0 |
0 |
0 |
0 |
1 |
1 |
Coleoptera |
8 |
17 |
11 |
19 |
20 |
45 |
| Coleoptera |
13 |
19 |
15 |
32 |
14 |
40 |
Diptera |
4 |
4 |
3 |
3 |
4 |
8 |
| Hymenoptera |
4 |
6 |
4 |
10 |
5 |
8 |
Hymenoptera |
3 |
9 |
3 |
7 |
3 |
7 |
| Hemiptera |
11 |
17 |
10 |
13 |
12 |
20 |
Hemiptera |
7 |
9 |
6 |
7 |
12 |
18 |
| Diptera |
8 |
9 |
6 |
9 |
4 |
7 |
Neuroptera |
1 |
1 |
1 |
1 |
0 |
0 |
| Trichoptera |
1 |
1 |
4 |
4 |
7 |
8 |
Orthoptera |
4 |
6 |
0 |
0 |
3 |
3 |
| Mantodea |
0 |
0 |
1 |
1 |
0 |
0 |
Odonata |
1 |
1 |
0 |
0 |
2 |
2 |
| Orthoptera |
3 |
4 |
1 |
1 |
4 |
4 |
Phasmida |
1 |
1 |
0 |
0 |
0 |
0 |
| Mecoptera |
1 |
1 |
1 |
1 |
1 |
1 |
Blattodea |
1 |
1 |
0 |
0 |
0 |
0 |
| Dermaptera |
1 |
1 |
1 |
1 |
0 |
0 |
Dermaptera |
1 |
1 |
0 |
0 |
1 |
1 |
| Neuroptera |
0 |
0 |
1 |
1 |
0 |
0 |
Ephemeroptera |
0 |
0 |
0 |
0 |
1 |
1 |
| Ephemeroptera |
0 |
0 |
1 |
1 |
4 |
4 |
|
|
|
|
|
|
|
| Odonata |
1 |
1 |
0 |
0 |
3 |
5 |
|
|
|
|
|
|
|
Download Excel Table
충주 조사지에서는 2022년 11목 43과 100종, 2023년 6목 42과 101종, 2024년 9목 70과 249종이 관찰되었다. 친환경 벌채 후 2024년 기준 2022년도에 비해 27과 149종, 2023년도에 비해 28과 148종이 증가하였다(Table 2).
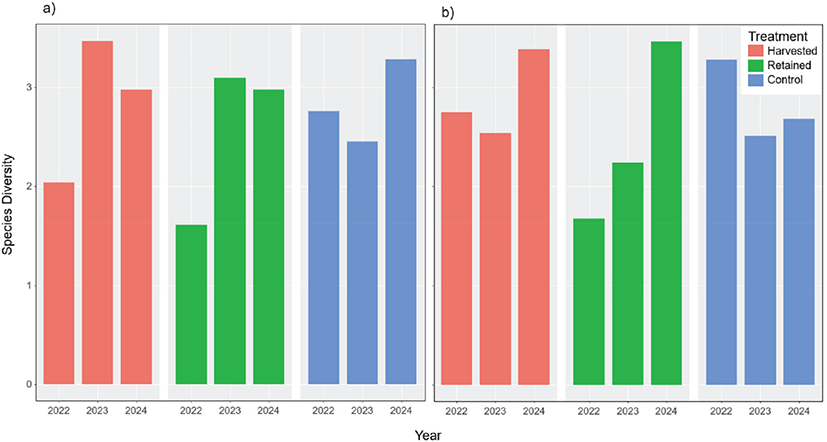
홍천과 충주 두 조사지 모두에서 2024년에 곤충 종다양도가 벌채 이전 및 전년도에 비해 증가하는 경향을 보였다(Figure 5). 홍천 벌채구의 초기 종다양도 지수는 낮게 나타났으나(2022년: 2.04), 이후 조사에서 증가하였다(2023년: 3.47, 2024년 2.98). 홍천 벌채구의 변동계수 CV(%)는 25.61로 나타났다. 잔존구의 경우 2022년 종다양도는 낮게 나타났으나(2022년: 1.61), 벌채 후 종다양도는 증가하였다(2023년: 3.10, 2024년: 2.98). 홍천 잔존구의 변동계수 CV(%)는 32.30로 나타났다. 홍천 대조구의 경우 종다양도 지수는 2022년에 비해 2023년이 소폭 감소하였으며(2022년: 2.76, 2023년: 2.45), 2024년에 증가하였다(2024년: 3.28). 홍천 잔존구의 변동계수 CV(%)는 14.87로 나타났다. 충주 벌채구의 초기 종다양도 지수는 낮게 나타났으나(2022년: 2.75), 이후 조사에서 증가하였다(2023년: 2.54, 2024년: 3.38). 충주 벌채구의 변동계수 CV(%)는 15.21로 나타났다. 잔존구의 경우 2022년 종다양도는 낮게 나타났으나(2022년: 1.67), 벌채 후 증가하였다(2023년: 2.24, 2024년: 3.47). 충주 잔존구의 변동계수 CV(%)는 37.24로 나타났다. 충주 대조구의 경우 2022년 이후 2년간 감소하였다(2022년: 3.28, 2023년: 2.51, 2024년: 2.68). 충주 대조구의 변동계수 CV(%)는 14.31로 나타났다. 조사구 모두에서 종다양도가 지속적으로 증가하는 경향이 관찰되었다([Figure 3(b)]).
Figure 5.
Species diversity (Shannon-Weaver Index) changes in forest insect species after retention harvesting for the a) Hongcheon and b) Chungju site.
Download Original Figure
4. 포유류 및 조류
홍천 조사지에서 6과 4목 8종이 관찰되었으며, 대조구에서는 6과 4목 7종의 흔적이 발견되었다. 대조구는 조사지에 비해 종의 흔적이 적게 나타났다. 충주 조사지에서는 조사지에서 6과 3목 7종이 관찰되었고, 대조구에서는 4과 2목 6종의 흔적이 확인되었다. 특히 두 대상지 모두에서 멸종위기 야생동물 II급인 삵(Prionailurus bengalensis)과 담비(Martes flavigula)가 발견되었다.
홍천 조사지에서 무인센서카메라에 의해 기록된 종은 벌채구에서 4목 7과 8종, 잔존구에서는 4목 8과 12종, 대조구에서는 5목 10과 14종이었다(Table 3). 다람쥐(Eutamias sibiricus), 등줄쥐(Apodemus agrarius), 청설모(Sciurus vulgaris), 오소리(Meles leucurus), 너구리(Nyctereutes procyonoides), 삵은 잔존구에서 벌채구와 대조구보다 더 자주 관찰되었다. 충주 조사지에서 무인센서카메라에 의해 기록된 종은 벌채구에서 4목 6과 7종, 잔존구에서는 3목 7과 9종, 대조구에서는 4목 9과 13종이었다(Table 3). 충주 조사지에서는 무인센서카메라 자료 분석 결과, 관찰된 종수와 개체수가 시간에 지남에 따라 증가하는 경향을 보였으며, 벌채구와 잔존구보다 대조구에서 더 많은 종과 개체가 기록되었다.
Table 3.
Relative Abundance Index of mammals by treatments.
| Hongcheon |
Chungju |
| Order |
Harvested |
Retained |
Control |
Order |
Harvested |
Retained |
Control |
|
Eutamias sibiricus
|
|
9.88 |
0.85 |
Eutamias sibiricus
|
|
0.06 |
0.76 |
|
Apodemus agrarius
|
|
0.55 |
0.03 |
Micromys minutus
|
0.39 |
0.06 |
|
|
Sciurus vulgaris
|
0.14 |
3.87 |
2.85 |
Apodemus agrarius
|
|
|
0.07 |
|
Lepus coreanus
|
0.05 |
0.21 |
0.31 |
Myodes rufocanus
|
|
|
0.03 |
|
Naemorhedus caudatus
|
|
|
0.47 |
Sciurus vulgaris
|
|
0.17 |
0.24 |
|
Hydropotes inermis
|
2.72 |
3.11 |
3.53 |
Lepus coreanus
|
|
|
0.07 |
|
Capreolus pygargus
|
10.68 |
12.71 |
15.76 |
Hydropotes inermis
|
0.06 |
20.17 |
29.41 |
|
Sus scrofa
|
1.38 |
3.94 |
8.14 |
Capreolus pygargus
|
20.06 |
4.71 |
16.26 |
|
Mustela sibirica
|
|
0.21 |
0.07 |
Sus scrofa
|
8.86 |
3.10 |
10.87 |
|
Meles leucurus
|
1.20 |
10.36 |
8.81 |
Mustela sibirica
|
1.27 |
|
0.31 |
|
Martes flavigula
|
|
0.21 |
1.22 |
Meles leucurus
|
|
1.22 |
14.60 |
|
Nyctereutes procyonoides
|
0.37 |
1.86 |
0.47 |
Martes flavigula
|
1.44 |
|
0.24 |
|
Prionailurus bengalensis
|
0.14 |
1.10 |
0.31 |
Nyctereutes procyonoides
|
|
1.88 |
0.35 |
|
Erinaceus amurensis
|
|
|
0.68 |
Prionailurus bengalensis
|
0.39 |
0.06 |
0.42 |
Download Excel Table
홍천 2022-2024년 조류 조사 결과 조사지에서 총 7목 24과 40속 52종 573개체, 대조구에서 총 7목 24과 40속 52종 220개체가 기록되었다. 충주 2022-2024년 조류 조사 결과 조사지에서 총 8목 24과 32속 39종 275개체, 대조구에서 총 8목 24과 32속 39종 241개체가 관찰되었다. 조사 결과, 두 조사지 모두 박새과(Paridae)를 포함하여 붉은머리오목눈이(Paradoxornis webbianus), 노랑턱멧새(Emberiza elegans) 등이 주요 조류 종으로 확인되었다. 충주 연구지에서는 멸종위기 야생동물 II급인 독수리(Aegypius monachus), 새매(Accipiter nisus)가 관찰되었으며, 홍천 연구지에서도 새매가 기록되었다. 벌채 이후 연구 대상지에서는 출현 종수와 개체수가 모두 증가하였다(Figure 6). 대조구에서는 출현 개체수는 증가한 반면, 출현 종수는 감소하는 경향을 보였다.
Figure 6.
Species diversity (Shannon-Weaver Index) changes in bird species after retention harvesting for the a) Hongcheon and b) Chungju site.
Download Original Figure
고 찰
1. 산림식생
산림 식생의 경우 친환경 벌채 이후 벌채구의 종다양도는 증가하는 경향을 보였으며, 이는 이질적이고 복잡한 환경이나 국지적 교란이 종다양도를 증가시킬 수 있다는 선행연구와 일치하였다(Barbour et al., 1980; Krebs, 1985; Kim et al., 2021). 벌채 후 수관량 증가로 토양 내 종자 발아가 촉진되어 종다양도가 증가한 것으로 나타났다(Lishawa et al., 2015). 또한, 하층부 광환경 개선은 초본 및 관목류의 번성을 촉진하여 종다양도 증가에 기여하였다(Zhang et al., 2023). 더불어, 벌채 후 조림 행동으로 인해 새로운 식재목 수가 일시적으로 증가하면서 종다양도가 높아졌다. 그러나 벌채 직후 며느리배꼽(Persicaria perfoliata), 환삼덩굴(Humulus japonicus)과 같은 덩굴식물 및 미국자리공(Phytolacca americana)과 같은 침입외래종의 점유 또한 종다양도 증가에 기여한 것으로 나타났다.
잔존구의 종다양도 증가는 가장자리 효과(edge effect)에 기인한 것으로 잔존구 주변부의 광환경이 개선되면서 하층식생의 번성이 촉진된 결과로 판단된다(Li et al., 2018). 그러나 잔존구의 변화 폭은 벌채구에 비해 상대적으로 낮았다. 이는 잔존구의 임분 구조가 그대로 유지되어 환경 변화가 제한적이었기 때문으로 해석된다(Nelson and Halpern, 2005). 보존된 임분 구조는 잔존구 내에서 천이 초기종의 확산을 저지하는 저항성 유지의 주요 요인으로 작용하는 것으로 보인다(Nelson and Halpern, 2005; Zenner et al., 2006).
2. 토양
산림 벌채 후 토양은 일시적인 유기물 손실을 겪으며, 이는 단기적으로 토양 환경을 변화시키고 장기적으로 토양 생산성에 영향을 미친다고 알려져 있다(Alban et al., 1978; Mann et al., 1988; Johnson et al., 1997). 특히 벌채로 인한 질소, 칼륨(K), 칼슘(Ca)과 같은 원소에서 영양분 손실이 크게 나타난 것으로 보고되었다. 따라서 본 연구에서 관찰된 결과는 산림 관리자가 벌채 시 신중을 기해야 함을 시사한다.
CEC의 감소는 벌채 후 강우가 토양으로 유입되면서 용탈이 증가한 데 기인한 것으로 판단된다(Chen and Xu, 2005). 이러한 용탈 증가는 토양 pH의 증가를 유발할 수 있는데, Park(1995)은 벌채 후 토양 상부 50 cm 층에서 토양 pH가 상승하였으며, 이는 양이온 손실에 따른 결과라고 보고하였다. 그러나 Park(1995)은 벌채 2년 후에는 토양의 화학적 특성에서 유의한 변화가 없었으며, 6년 후에는 질소-질산태 농도가 벌채 이전 수준으로 회복되었다고 하였다. Lebo and Herrmann(1998)도 유사한 결과를 제시하였으며, 질소 용탈은 벌채 후 2-3년 내에 벌채 이전 수준으로 회복되었다고 보고하였다.
토양 pH의 경우, 벌채 이후 증가하는 것으로 나타났다. 이는 벌채 이후 수목에 의해 공급되는 수소이온이 감소하고, 무기토양으로부터 칼슘(Ca), 마그네슘(Mg)과 같은 2가 양이온의 증가에 기인하는 것으로 해석된다(Farahnak et al., 2022). 또한, 유효인산은 벌채 후 증가하는 것으로 나타났으며, 이는 벌채로 인한 유기물 분해의 일시적 증가에 기인하는 것으로 해석된다(Noh et al., 2023).
본 연구 결과는 소규모 벌채 및 친환경 벌채와 같은 경우 토양에 미치는 영향이 제한적임을 시사한다. 그러나 벌목 기계의 운행에 따른 경운이나 압밀과 같은 토양 교란은 여전히 발생할 수 있다. 토양은 시간이 경과함에 따라 안정화될 것으로 예상되지만, 추가적인 변화가 나타날 가능성도 있으며, 침엽수 조림지와 활엽수 조림지 간의 토양 발달 양상은 차이를 보일 수 있으므로 지속적인 모니터링이 필요하다.
3. 곤충
산림 벌채는 식생 구조, 국지적 일조 조건, 미세기후를 변화시켜 곤충 군집의 구성과 다양성에 큰 영향을 미칠 수 있다(Summerville and Crist, 2002). 벌채 이후 산림 하층의 광환경 개선으로 인한 초본 및 천이 초기종의 번성은 다양한 곤충 종을 유입시켜 단기적으로 종다양도를 증가시킬 수 있다(Swanson et al., 2011). 특히 본 연구에서 나방류의 채집 개체수가 증가한 것은 비행 장애 요소의 제거에 기인한 것으로 해석된다. 또한, 벌채로 인해 형성된 서식지 자원(예: 초지의 확장, 그루터기 및 벌채 잔재물의 증가)은 딱정벌레목(Coleoptera)과 노린재목(Hemiptera)의 종수를 증가시키는 요인으로 작용하였다.
일반적인 개벌과 달리, 본 연구에서 적용된 친환경 벌채는 성숙 임분 구조를 보전하여 성숙 산림에 의존하는 종의 감소 우려를 완화하는 효과가 있었다. 본 연구 결과는 조사구의 곤충 군집이 다양한 종으로 구성되어 있으며, 벌채 이후 회복 과정을 거치고 있음을 시사한다. 그러나 친환경 벌채가 생태계 과정을 모방하려는 목적에도 불구하고, 비벌채 임분과 비교할 때 수관 지배 양상이 변화하는 것으로 보고된 바 있다(Swanson et al., 2011). 비록 본 연구에서는 다루지 않았지만, 벌목 기계로 인한 토양 교란은 지표성 곤충의 서식지를 저하시키며 부정적인 영향을 미칠 수 있다(Menta and Remelli, 2020). 따라서 산림 관리자는 회복 과정이 산림 유형, 기후 변화, 관리 이력 및 체계 등 다양한 요인에 따라 달라질 수 있음을 고려해야 한다. 부정적 영향을 완화하고, 벌채 이전 상태로의 회복을 보장하며, 생물다양성을 유지하기 위해서는 장기적인 모니터링이 필수적이다.
4. 포유류 및 조류
무인센서카메라 조사 결과 전반적으로 대조구에서의 포유류 RAI가 조사지보다 높게 나타났는데, 이는 벌채 1년 후(2023년)까지 서식지가 완전히 회복되지 않았음을 시사한다. 대조구의 서식지는 상대적으로 안정적이며 은폐 구조가 풍부하여 대부분의 포유류 종에서 높은 출현 빈도를 보인 것으로 판단된다. 그러나 충주 지역 노루의 경우는 벌채구에서 잔존구와 대조구에 비해 RAI가 높게 나타났다. 이는 벌채 이후 새롭게 형성된 초본류와 같은 먹이 자원 증가와 개방된 환경이 일부 종에게 더 적합한 서식 조건을 제공하여, 벌채구에서 활동 빈도가 높아진 결과로 해석된다(Augustine and McNaughton, 1998).
조사지에서 관찰된 조류 종수의 증가는 벌채 후 서식지의 회복이 진행되면서 다양한 조류 종의 서식지 이용이 점차 확대된 데 기인한 것으로 판단된다. 그러나 벌채 작업이 진행 중이던 2022년 여름과 가을에는 조류 활동이 일시적으로 감소했을 가능성도 있다. 일부 조류가 활동을 줄이거나 대체 서식지로 이동한 경우, 벌채지에서의 관찰 개체수는 일시적으로 감소할 수 있다. 반면, 본 연구 결과에서 대조구의 서식지가 비교적 안정적임에도 불구하고 조류 다양성이 감소하는 경향을 보였다. 이는 뚜렷한 환경 변화가 없는 상태에서 일부 특정 종의 개체수 증가에 기인한 종다양도 감소로 해석된다. 또한 벌채 작업 과정에서 일부 조류 종이 대조구로 이동함에 따라, 대조구에서 상대적으로 더 많은 종이 관찰되었을 가능성도 있다.
결 론
본 연구는 친환경 벌채가 산림생태계의 식생 및 동물상, 토양 특성에서 조성, 구조, 종다양도 등의 초기 변화를 유발할 수 있음을 시사한다. 벌채 이후 햇빛 노출과 천이 초기종의 번성은 생태계 회복을 촉진하고 식생의 종다양도를 크게 증진시키는 요인으로 작용하였다 [홍천: 1.10±0.08(’22) → 1.28±0.09(’24), CV(%) = 9.07 / 충주: 0.73±0.19(’22) → 1.05±0.16(’24), CV(%) = 17.86]. 두 지역 모두 식생의 종다양도는 크게 변화하였다. 그러나 용탈 증가 및 양이온치환용량(CEC) 변화 등 토양 특성의 변화는 장기적인 토양 생산성 저하를 방지하기 위한 신중한 관리의 필요성을 강조한다. 곤충 군집은 벌채 이후 회복 양상을 보였으며, 특히 새로운 식재목과 다양한 관목의 출현은 곤충 생존에 유리한 조건을 제공하였다. 이로 인해 꽃가루매개곤충의 다양도가 증가하였고, 결과적으로 곤충 종다양도가 크게 증진되었다 [홍천: 2.04(’22) → 2.98(’24), CV(%) = 25.61 / 충주: 2.75(’22) → 3.38(’24), CV(%) = 15.21]. 곤충의 종다양도의 변동성은 잔존구, 벌채구, 대조구 순서로 나타났다. 곤충의 종다양도 증가는 서식지 구조와 먹이 자원의 변화에 기인한 것으로 판단된다. 포유류와 조류는 종에 따라 서로 다른 반응을 보였으며, 일부 종은 먹이 자원 증가로 인해 교란지에서 선호도가 높아진 반면, 일부 종은 일시적으로 개체수가 감소하였다. 본 연구 결과는 벌채 후 1~2년 초기 모니터링 결과로서, 식생, 곤충의 종다양도가 빠른 회복세를 보였다. 친환경 벌채 이후 회복세의 장기 모니터링은 산림관리를 위해 필수적일 것이라고 판단한다. 본 연구 결과는 친환경 벌채 이후 생물다양성 회복과 생태계 안정성을 확보하기 위해 지속적인 모니터링과 친환경적 산림경영의 중요성을 강조한다.
감사의 글
본 연구는 산림청의 “친환경 벌채 대상지 모니터링” 과제의 지원에 의하여 이루어진 것입니다.
References
Abdi, H. 2010. Coefficient of variation. Encyclopedia of Research Design 1(5): 169-171.

Akashi, N., 2023. Responses of understory vascular plant communities up to 6 years after retention harvesting in planted
Abies sachalinensis forests. Forest Ecology and Management 538: 120991.

Alban, D.H., Perala, D.A. and Schlaegel, B.E., 1978. Biomass and nutrient distribution in aspen, pine, and spruce stands on the same soil type in Minnesota. Canadian Journal of Forest Research. 8(3): 290-299.
An, H., Seok, H.D., Lee, S.M. and Choi, J., 2019. Forest management practice for enhancing carbon sequestration in national forests of Korea. Forest Science and Technology 15(2): 80-91.
Augustine, D.J. and McNaughton, S.J., 1998. Ungulate effects on the functional species composition of plant communities: herbivore selectivity and plant tolerance. The Journal of Wildlife Management. 62(4): 1165-1183.
Bang, S. 2017. Case study of the conversion of forest management mountain areas into the development sites and elicitation of forest management improvement. Korea Environment Institute. Research Report
Barbour, M.G., Burk, J.H. and Pitts, W.D., 1980. Terrestrial plant ecology. California, U.S.A. pp. 604.
Bliss, J.C., 2000. Public perceptions of clearcutting. Journal of Forestry 98(12): 4-9.
Braun-Blanquet, J., 1964. Pflanzensoziologie: Grundzuege der Vegetationskunde; dritte, neubearbeitete und wesentlich vermehrte Auflage. Springer Verlag.
Carpentier, S., Filotas, E., Handa, I.T. and Messier, C., 2017. Trade-offs between timber production, carbon stocking and habitat quality when managing woodlots for multiple ecosystem services. Environmental Conservation. 44(1): 14-23.
Chen, C.R. and Xu, Z.H., 2005. Soil carbon and nitrogen pools and microbial properties in a 6-year-old slash pine plantation of subtropical Australia: impacts of harvest residue management. Forest Ecology and Management 206(1-3): 237-247.
Farahnak, M., Mitsuyasu, K., Ide, J., Chiwa, M., Enoki, T., JEong, S., Otsuki, K., Shimizu, K. and Kume, A. 2022. Soil pH and divalent cations after clear-cutting on a Japanese cypress plantation. Journal of Forest Research 27(5): 363-370.
Franklin, J.F. and Donato, D.C., 2020. Variable retention harvesting in the Douglas-fir region. Ecological Processes. 9(1): 8.
Franklin, J.F., Berg, D.R., Thornburgh, D.A. and Tappeiner, J.C., 1997. Alternative silvicultural approaches to timber harvesting: variable retention harvest systems. Creating a forestry for the 21st century: the science of ecosystem management. Island Press. Washington, DC, U.S.A. pp.111-139.
Jang, E.K. and Youn, Y.C., 2021. Effects of wood product utilization on climate change mitigation in South Korea. Sustainability 13(12): 6737.
Johnson, C.E., Romanowicz, R.B. and Siccama, T.G., 1997. Conservation of exchangeable cations after clear-cutting of a northern hardwood forest. Canadian Journal of Forest Research 27(6): 859-868.
KFS (Korea Forest Service). 2017. The 6th Forest Basic Plan (2018-2037); Korea Forest Service: Seoul, Republic of Korea.
Kim, D.S. et al. 2020a. Investigation of vegetation and insect in Green Tree Retention, Mokgye-ri, Wangsan-myeon, Gangneung, Korea. p. 49. In: Proceedings of the Korean Society of Environment and Ecology Conference. 30: 49.
Kim, D.S., Park, S.W., Roh, S.J., Jeon, J.H. and Yoo, T.H. 2016. Responses of bark beetle (Coleoptera: Curculionidae: Scolytinae) community structure to green-tree retention in pine tree forest from Korea. Journal of Asia-Pacific Biodiversity 9(4): 443-447.
Kim, H., Lee, J., Kim, H. and Yun, C., 2024. Forest community structure of aggregated retention harvest for
Larix kaempferi. Korean Journal of Environmental Biology 42(2): 176-186.
Kim, M., Kraxner, F., Forsell, N., Song, C. and Lee, W.K., 2021. Enhancing the provisioning of ecosystem services in South Korea under climate change: The benefits and pitfalls of current forest management strategies. Regional Environmental Change. 21(1): 6.
Kim, M.S., Kim, J.S., Kim, H.S., Park, C.W. and Bae, K.H., 2020b. Changes in community structure of understory vegetation by silvicultural treatments in a
Larix kaempferi plantation forest. Journal of Agriculture & Life Science 54: 23-33.
Ko, D.W., Sung, J.H., Lee, Y.G. and Park, C.R., 2015. The current status and challenges of forest landscape models. Journal of Korean Forest Society 104(1): 1-13.
Krebs, C.J. 1985. Ecology (3rd Ed.) Haber and Row Publishing Company, New York, U.S.A.
Lamelas-López, L., Fontaine, R., Borges, P.A. and Gonçalves, D., 2020. Impact of introduced nest predators on insular endemic birds: the case of the Azores Woodpigeon (
Columba palumbus azorica). Biological Invasions 22(12): 3593-3608.
Lebo, M.E. and Herrmann, R.B., 1998. Harvest impacts on forest outflow in coastal North Carolina. Journal of Environmental Quality 27(6): 1382-1395.
Lee, C.W. 2005. Silvicultural Treatment for Reduction of Forest Fire Hazard. In Proceedings of the Korea Institute of Fire Science and Engineering Conference. Korean Institute of Fire Science and Engineering. pp. 278-292.
Li, J., Zhao, C., Peng, Y., Hu, Y. and Yuan, X., 2018. Edge effects on tree growth and species diversity in forests of different types and ages. Polish Journal of Ecology. 66(3): 239-249.
Lishawa, S.C., Lawrence, B.A., Albert, D.A. and Tuchman, N.C., 2015. Biomass harvest of invasive Typha promotes plant diversity in a Great Lakes coastal wetland. Restoration Ecology 23(3): 228-237.
Macdonald, S.E. and Fenniak, T.E., 2007. Understory plant communities of boreal mixedwood forests in western Canada: natural patterns and response to variable-retention harvesting. Forest Ecology and Management 242(1): 34-48.
Mann, L.K., et al., 1988. Effects of whole-tree and stem-only clearcutting on postharvest hydrologic losses, nutrient capital, and regrowth. Forest Science 34(2): 412-428.
Nelson, C.R. and Halpern, C.B., 2005. Edge-related responses of understory plants to aggregated retention harvest in the Pacific Northwest. Ecological Applications. 15(1): 196-209.
NIBR (National Institute of Biological Resources). 2022. National list of Korea. National institute of Biological Resources. Incheon, Korea, Accessed 24 Aug 2025,
https://kbr.go.kr/
Noh, N.J., Han, S.H., Lee, S.T. and Cho, M.S. 2023. Changes in Soil Physiochemcial Properties Over 11 Years in
Larix kaempferi Stands Planted in
Larix kaempferi and
Pinus rigida Clear-Cut Sites. Journal of Korean Society of Forest Science 112(4): 502-514.
O’Connell, A.F., J.D. Nichols and Karanth, K.U. (eds.). 2011. Camera traps in animal ecology: methods and analyses. Springer. New York, U.S.A.
Park, J.H., 1995. Environmental changes after timber harvesting in (Mt.) Paekunsan. Journal of Korean Society of Forest Science 84(4): 465-478.
Shannon, C. E. and W. Weaver. 1949. The mathematical theory of communication. Urban. Univ. Illinois Press. Illinois, U.S.A. pp. 125.
Summerville, K.S. and Crist, T.O., 2002. Effects of timber harvest on forest Lepidoptera: community, guild, and species responses. Ecological Applications 12(3): 820-835.
Swanson, M.E., Franklin, J.F., Beschta, R.L., Crisafulli, C.M., DellaSala, D.A., Hutto, R.L., Lindenmayer, D.B. and Swanson, F.J., 2011. The forgotten stage of forest succession: early-successional ecosystems on forest sites. Frontiers in Ecology and the Environment 9(2): 117-125.
Tak, K., Chun, Y., and Wood, P. 2007. The South Korean forest dilemma. International Forestry Review. 9: 548-557.
Vanha-Majamaa, I. and Jalonen, J., 2001. Green tree retention in Fennoscandian forestry. Scandinavian Journal of Forest Research. 16: 79-90.
Wang, R. J. and Kim, D. Y., 2022. Early effect of environment-friendly harvesting on the dynamics of organic matter in a Japanese Larch (
Larix leptolepis) Forest in Central Korea. Journal of Korean Society of Forest Science 111(4): 473-481.
Zenner, E.K., Kabrick, J.M., Jensen, R.G., Peck, J.E. and Grabner, J.K., 2006. Responses of ground flora to a gradient of harvest intensity in the Missouri Ozarks. Forest Ecology and Management 222(1-3): 326-334.
Zhang, Y., Zhao, X., Wang, L., Wang, Z., Shuai, H., Wang, Y., Jin, B. and Chen, C., 2023. Effects of clearcutting on species composition and community renewal of Rhododendron shrubs in northwest Guizhou Province, China. Frontiers in Ecology and Evolution. 11: 1225466.