서 론
백두대간은 지표상의 분수계를 중심으로 산계의 흐름을 파악할 수 있는 민족 고유의 지리인식체계로서의 의미를 지니며 남북으로 길게 뻗은 한반도의 지형축으로 능선을 중심으로 펼쳐진 자연환경은 다양한 동·식물의 서식처로 상호 연결된 생태계 축을 형성하여 자연환경 및 자원의 보존이라는 생태적 가치 측면에서 중요한 의미를 갖는다(Kim and Choo, 2003). 백두대간의 인식은 위와 같은 지형학적인 고찰 이전에 우리나라 사람들의 풍수지리적인 산지인식체계, 즉 최고 고산지에 대한 경외와 백두산부터 지리산까지의 산줄기를 국토지리적으로 일체화하려는 오랜 신념적, 관념적 자연관이 함축되어온 체계라 할 수 있다(Lee, 1991; Cho, 1994).
백두대간은 백두산 병사봉에서 시작해 지리산 천왕봉에 이르기까지 1,400 km에 이르며(Shin, 2004), 남한의 백두대간은 설악산권역(향로봉-구룡령), 태백산권역(구룡령-깃대배기봉), 속리산권역(깃대배기봉-청화산), 덕유산권역(청화산-남덕유산), 지리산권역(남덕유산-천왕봉)으로 나눌 수 있으며(Korea Forest Service, 2016), 본 연구대상지는 육십령-천왕봉 구간의 마루금으로 지리산권역 전체에 해당한다. 지리산은 한반도 남부에서 가장 높은 해발고도를 지닌 ‘한반도 남쪽의 지붕’으로 불리우는 산지로 경상남도, 전라남도, 전라북도 3도의 경계부에 솟아 있으며 그 크기와 면적에 걸맞게 한국인의 다양한 정신문화를 담고 있어 크기와 깊이를 가늠하기 어려운 그릇이 되어 왔다. 또한 산지기후임에도 해양성 기후와 대륙성 기후를 분리시키는 중간적인 위치를 점하고 있어 고도에 따른 산악 특성이 잘 반영된 기후가 나타나 그에 따른 식생을 포함한 다양한 자연생태계 자원의 총체라 할 수 있다(Beom, 2008).
식생은 기후, 지형, 토양 등에 의하여 결정되지만 생물의 영향과 사람의 활동에 의하여 그 양상이 변화하여 현존식생을 이루게 된다. 이러한 식생은 어느 특정지역에 다양한 식물 종으로 구성되어 있고 이 종들은 공간과 환경적 특성 등에 의해 분포한다. 따라서 일정한 환경조건 하의 서식지에는 우점하는 몇몇 식물군락들로 구성되고 이들 각각의 식생은 군집 내 우점하는 식물의 특성과 형태에 따라 특정된다(Barbour et al., 1987). 한반도 남부의 식생학적 성격은 중남부 또는 남부 내륙형의 냉온대 남부·저산지형 및 중부·산지형의 낙엽활엽수림이 혼재해 있으며, 한반도 식물사회학적 분류체계로는 신갈나무-생강나무아군단과 졸참나무-작살나무아군단에 속한다(Kim and Lee, 2006). 백두대간 남부권역 주요 산지의 산림은 소나무림형, 신갈나무림형, 굴참나무-졸참나무림형, 중생혼합림형, 서어나무림형, 구상나무림형, 기타활엽수림형 등으로 구분된다고 알려져 있지만(Lee et al., 2014) 백두대간 마루금 전체를 대상으로 한 산림식생구조를 파악할 필요가 있다.
1990년대부터 백두대간에 대한 관심이 부각되기 시작하면서 다양한 연구들이 선행되었으며 그 가운데 백두대간 내 산림식생에 대한 연구도 활발히 수행되어 왔다(Hwang et al., 2012). 백두대간 지리산권역의 훼손지 식생 및 식물상에 관한 연구로는 지리산권역 내 존재하는 훼손지 유형별 실태조사(Lee et al., 2020), 백두대간의 식물상, 식생, 이용 및 환경훼손의 기존 학술자료들을 정리, 분석한 연구(Oh and Lee, 2003), 덕유산 향적봉부터 지리산 천왕봉까지의 식물상을 정리한 연구(Lim, 2003)가 있었으며, 식생구조 및 산림유형분류를 실시한 연구로는 백두대간 노고단-고리봉 구간의 식생구조를 살펴본 연구(Kim and Choo, 2003), 백두대간 남부권역 천연림의 산림 분류에 관한 연구(Lee et al., 2014)가 이루어져 왔지만 통계패키지를 활용한 군집분석이 주를 이루고 있었고, 군락유형을 인식하고 분류할 때에 식물사회학적(floristic-sociological)인 측면, 즉 식물사회의 종조성을 강조하는 Z-M학파의 전통군락분류법(Whittaker, 1965)에 의한 식생유형분류는 거의 이루어지고 있지 않았다.
따라서, 본 연구는 백두대간 지리산권역(육십령-천왕봉 구간)의 산림식생을 대상으로 식물사회학적 유형분류를 통해 종조성을 파악하여 식생단위를 구분하고 이를 정량적으로 검증하고 구분된 식생단위별 식생구조를 파악하기 위하여 층위별 중요치, 군락유사도, 종다양성, 환경인자와의 상관관계를 분석하여 연구대상지의 산림식생구조를 밝히는데 목적을 두었다.
재료 및 방법
본 연구는 남한의 백두대간 구간 내, 산림청에서 지정한 5개의 권역(Korea Forest Service, 2016) 중 지리산권역에 해당하는 육십령(734 m)에서부터 백운산(1,279 m), 복성이재(550 m), 고리봉(1,304 m), 성삼재(1,102 m), 노고단(1,507 m), 벽소령(1,350 m), 제석봉(1,806 m)을 거쳐 지리산 천왕봉(1,915 m)까지 총 101.0 km 구간을 연구대상지로 선정하였으며(Figure 1) 조사대상지의 해발고도의 분포는 최저 492 m에서 최고 1,867 m이다. 조사지의 식물상 지역은 대부분 상량 온대 식물상 지역에 속하지만 온난 온대 식물상 지역에 해당하는 지역도 분포하고 있다(Cho et al., 2020). 지리산 권역 내 지질구조를 살펴보면 육십령 부근은 중생대 쥬라기 복운모화강암이 넓게 분포하고 있고, 백운산 부근은 선캄브리아기 반상 변정질 편마암, 복성이재 부근은 중생대 트라이아스기 중립질화강암, 고리봉에서 성삼재 구간은 화강암질편마암, 노고단 부근은 미그마타이트질 편마암, 벽소령에서 천왕봉 구간은 혼성페그마타이트질편마암으로 주로 구성되어 다양한 지질이 나타나는 것을 알 수 있었다(Korea Institute of Geoscience and Mineral Resources, 2021).
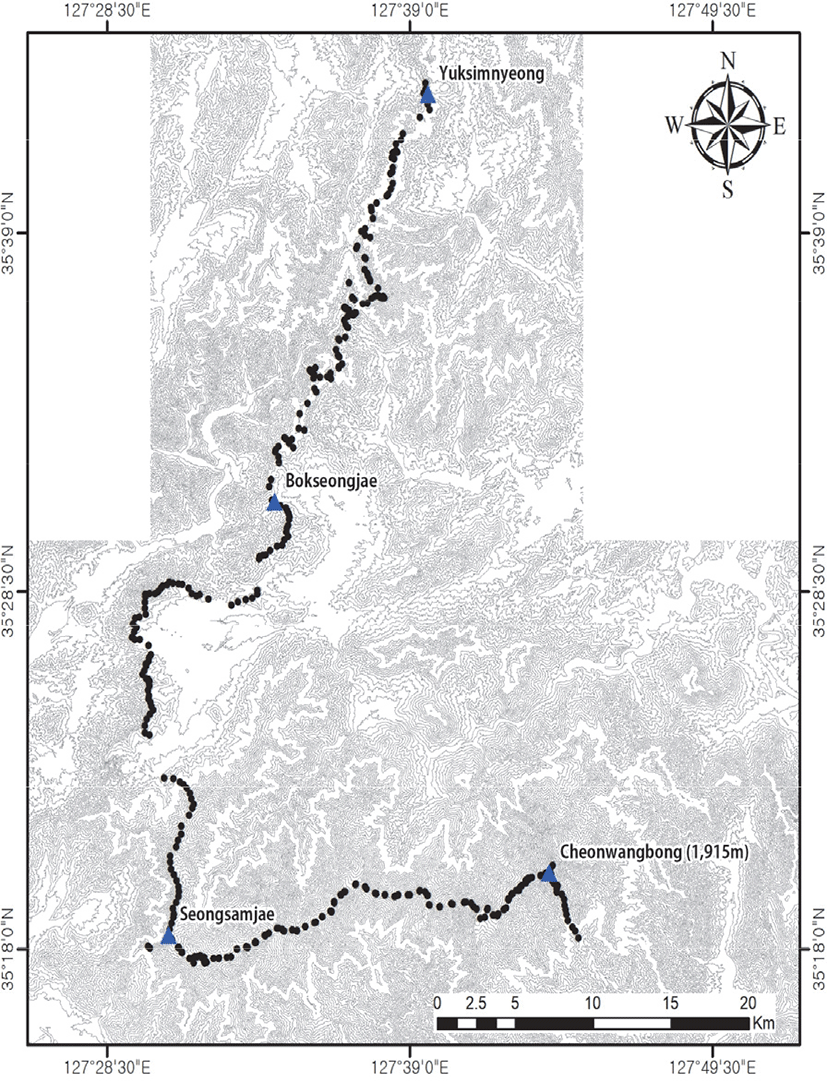
조사기간은 2020년 4월부터 동년 10월까지 백두대간 육십령(m)에서 지리산 천왕봉(m) 구간의 마루금 식생을 대상으로 실시하였다. 조사지점 선택의 임의성을 낮추고자 조사지점 간 거리는 약 500 m 간격으로 설정하였으며 마루금을 기준하여 좌우 양안으로 50 m 이상 떨어진 지점에 20 m × 20 m 크기의 총 373개소의 방형구를 선정하였다. 식물사회학적 방법(Ellenberg, 1956; Braun-Blanquet, 1964)에 따라 조사구내에 출현하는 각 종의 피도(coverage)와 개체수를 조합시킨 우점도(dominance) 계급을 층위별로 구분하여 판정 기록하였고 생육상태는 종 개체의 집합 혹은 이산의 정도에 따른 군도(sociality) 계급 등을 측정하였다. 환경인자의 측정은 해발고도, 사면경사각, 사면방위, 암석노출도, 낙엽층깊이 등을 측정 및 기록하였고 층위별 평균식피율, 층위별 평균수고, 층위별 평균흉고직경 등과 같은 식생의 구조를 파악하였다. 군락유형 분류 후, 연속계수가 아닌 정성적 범주화 계수인 지형의 경우, 산정(T) 10, 능선부(R) 9, 사면상부(US) 7, 사면중부(MS) 5, 사면하부(LS) 3, 계곡부(V) 1로 정량적 값으로 치환하여 기술통계적 분석을 실시하였다. 구성종의 식물분류와 동정은 원색식물도감(Lee, 2003), 원색한국수목도감(Hong et al., 1987), 나무생태도감(Yun, 2016), 한국의 나무(Kim and Kim, 2011), 한국의 양치식물(Lee and Lee, 2015), 벼과·사초과 생태도감(Cho et al., 2016)을 기준으로 하였으며 국가표준식물목록(Korea National Arboretum, 2020a)과 국가생물종지식정보시스템(Korea National Arboretum, 2020b)을 기준으로 학명과 국명을 작성하였다.
373개소 방형구의 식생자료를 바탕으로 Ellenberg(1956)의 표조작법과 Hill(1979a)의 TWINSPAN을 이용하여 소표(raw table)로부터 여러 단계의 표조작과정을 거쳐 최종적으로 상재도를 나타낸 식별표를 작성하여 식생유형분류를 실시하였다. 군락의 분포특성을 파악하고자 DECORANA (Detrended correspondence analysis)를 이용하여 요인분석을 실시하였으며 DECORANA를 수행한 결과를 토대로 이차원 공간에 방형구들을 배열하여 방형구들간의 유연관계를 분석하였다(Hill, 1979b). 식생유형분류를 통해 산출된 각각의 식생단위(vegetation unit)별 층위별 점유율을 파악하기 위해 조사지내 수종을 대상으로 Curtis and McIntosh(1951)의 방법을 이용하여 상대빈도(RF), 상대피도(RC)를 합산하여 평균한 중요치((상대빈도+상대피도)/2)를 분석하였다. 일정 면적 내의 종수, 또는 군집 내에서 일정수의 개체를 구성하는 종수를 의미하는 종풍부도(species richness)와 종의 양적 척도에 대한 균질한 정도를 나타내는 균등도(species evenness)를 동시에 나타내는 척도인 종다양도지수(H′=−∑(pi*logpi)), 종풍부도(S=(s-1)/logN)), 종균재도(J′=H′/logS), 종우점도(D=1-∑ni(ni-1)/N(N-1))를 분석하였다(Shannon, 1949; Herlbert, 1971; Brower and Zar, 1977). 환경인자(해발고도, 사면경사각, 지형, 암석노출도, 낙엽층깊이)와 식생단위와의 상관관계를 정량적이고 객관적으로 파악하기 위해 NMS(Non-Metric Multidimensional Scaling) 분석을 실시하였고 서열척도(ordinal scale)로 기재하여 활용하였다. 환경인자 중 지형은 사면하부(LS)는 1, 사면중부(MS)는 3, 사면상부(US)는 5, 능선(R)은 7, 정상(T)은 9로 환산하여 정량화하였으며 수리통계처리를 위하여 통계처리 프로그램인 PC-ORD v5.17을 이용하여 Biplot cutoff R2: 0.200으로 분석하였다.
결과 및 고찰
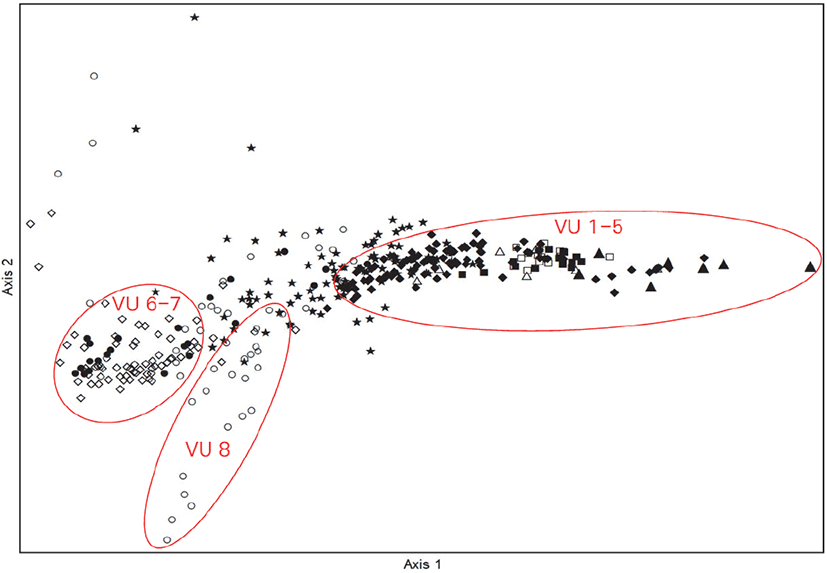
백두대간 지리산권역(육십령-천왕봉 구간) 마루금 내 373개소의 산림식생조사 자료를 바탕으로 Z-M 식물사회학적 방법에 의해 식생유형 분류를 수행한 결과(Table 1), 최상위 수준에서 신갈나무군락군(Quercus mongolica community group)으로 분류되었고, 신갈나무군락군은 구상나무군락(Abies koreana community), 소나무군락(Pinus densiflora community), 비목나무군락(Lindera erythrocarpa community), 신갈나무전형군락(Q. mongolica typical community)으로 세분되었다. 구상나무군락은 가문비나무군(Picea jezoensis group), 둥근이질풀군(Geranium koreanum group), 함박꽃나무군(Rhododendron tschonoskii group), 구상나무전형군(A. koreana typical group)으로 세분되었고, 가문비나무군은 흰참꽃나무소군(R. tschonoskii subgroup)과 가문비나무전형소군(P. jezoensis typical subgroup)으로 각각 세분되었다. 소나무군락은 비목나무군(L. erythrocarpa group)과 신갈나무전형군(Q. mongolica typical group)으로 세분되어 총 9개의 식생단위와 9개의 종군으로 분류되었다. 최상위단위에 신갈나무군락군으로 분류된 것은 우리나라 산림식생이 전체적으로 신갈나무 군락형으로 대표된다는 결과(Yun et al., 2011)와 백두대간 남한지역 산림식생의 구조와 종조성에서 신갈나무가 가장 최상위 계층에 넓게 형성되어 있다는 결과(Hwang, 2016)와 일치하였다.
* The other 418 companion species omitted.
본 식생단위는 신갈나무군락군에서 종군 2의 구상나무, 잣나무 등의 식별종 출현에 의해 구상나무군락으로 구분되었고, 구상나무군락의 하위종군으로 가문비나무군으로 구분되어졌으며 가문비나무군의 하위종군으로 흰참꽃나무소군으로 세분되었다. 본 식생단위에 출현하는 우점도가 3이상인 식물종은 철쭉, 미역줄나무, 실새풀 등 총 14분류군이 출현하였다. 식생단위 1은 가장 높은 암석노출도를 보였고 암석지대에 쉽게 자생하는 바위떡풀, 금마타리, 바위채송화, 돌양지꽃, 참바위취 등의 식물과 함께 하나의 종군을 구성하였다. 흰참꽃나무 역시 백두대간 수령-소사고개 구간에서 삼봉산 주변 해발고 1,200 m 부근의 암석으로 능선부를 형성한 지점에 큰 군락이 분포하고 있다는 기존의 연구 결과처럼(Choo and Kim, 2004), 노암율이 높은 지역에 쉽게 자생하는 것을 알 수 있었다.
본 식생단위는 신갈나무군락군에서 종군 3 이하의 종군에서 식별종들이 출현하지 않음으로 인하여 종군 3의 가문비나무, 두루미꽃 등의 식별종 출현에 의한 가문비나무전형군으로 구분되었다. 본 식생단위에 출현하는 우점도가 3이상인 식물종은 실새풀, 철쭉 등 총 16분류군이 출현하였다. 가문비나무는 우리나라 계방산, 덕유산, 지리산의 아한대림에 분포한다고 알려져 있으며(Miyawaki, 1988; Nikolove and Helmisaari, 1992), 구상나무, 잣나무, 사스레나무, 시닥나무, 마가목 등 아고산대 지역에서 주로 자생하는 종들과 더불어 상대우점치가 높게 나타난다는 기존의 연구와 일치하였다(An et al., 2010; Ko et al., 2014).
본 식생단위는 신갈나무군락군에서 종군 2의 구상나무군락의 하위종군으로 둥근이질풀, 기린초 등의 식별종에 의해 둥근이질풀군으로 세분되었다. 본 식생단위에 출현하는 우점도가 3이상인 식물종은 철쭉, 미역줄나무 등 총 10분류군이 출현하였다. 식생단위 3은 교목층과 아교목층이 존재하지 않고 관목층과 초본층으로만 이루어진 지리산 노고단과 제석봉 지역 위주의 왜림 또는 초원지대로서 소백산 비로봉 초원 지역과 유사한 입지환경과 종조성을 보여(Song and Yun, 2019) 경관생태학적인 장기적 모니터링이 필요할 것으로 판단되었다.
본 식생단위는 신갈나무군락군에서 종군 2의 구상나무군락의 하위종군으로 흰참꽃나무, 바위떡풀 등의 식별종에 의해 흰참꽃나무군으로 세분되었다. 본 식생단위에 출현하는 우점도가 3이상인 식물종은 신갈나무, 실새풀 등 총 11분류군이 출현하였다.
본 식생단위는 신갈나무군락군에서 종군 2 이하의 종군에서 식별종들이 출현하지 않음으로 인하여 종군 2의 구상나무, 잣나무 등의 식별종 출현에 의한 구상나무전형군락으로 구분되었다. 본 식생단위에 출현하는 우점도가 3이상인 식물종은 신갈나무, 철쭉 등 총 39분류군이 출현하였다. 구상나무는 한국의 특산식물로 IUCN 적색목록 평가에서 위기종(EN)에 등재되는 등 국내·외 많은 학계의 관심을 받고 있는 종으로 기후변화에 의해 서식처가 급속도로 줄고 있는 종 가운데 하나이다(Kim and Hyun, 2000; Kim et al., 2001; Yoon and Kong, 2011). 지리산 구상나무림에 관한 식물사회학적 기존 연구에서 구상나무림 아래 층위의 종조성 비교에서 대부분 유사한 종조성을 보였고(Kim et al., 2018a; Kim et al., 2018b), 덕유산 구상나무림의 경우, 신갈나무, 조릿대, 주목 등과 주로 함께 나타나는 등의 종조성 차이를 보였다(Kim et al., 2011).
본 식생단위는 신갈나무군락군에서 종군 6의 소나무, 주름조개풀 등의 식별종에 의해 소나무군락으로 구분되었고, 소나무군락의 하위종군으로 비목나무군으로 세분되었다. 본 식생단위에 출현하는 우점도가 3이상인 식물종은 신갈나무, 가는잎그늘사초 등 총 25분류군이 출현하였다.
본 식생단위는 신갈나무군락군에서 종군 6 이하의 종군에서 식별종들이 출현하지 않음으로 인하여 종군 6의 소나무, 주름조개풀 등의 식별종 출현에 의한 소나무전형군락으로 구분되었다. 본 식생단위에 출현하는 우점도가 3이상인 식물종은 신갈나무, 철쭉 등 총 17분류군이 출현하였다.
본 식생단위는 신갈나무군락군에서 종군 7의 비목나무, 산뽕나무 등의 식별종에 의해 비목나무군락으로 구분되었다. 본 식생단위에 출현하는 우점도가 3이상인 식물종은 신갈나무, 잣나무 등 총 38분류군이 출현하였다. 중부지방 비목나무 자생림의 식물군집구조를 분석한 기존 연구에서 비목나무와 함께 생육하는 종들로 소나무, 졸참나무, 느티나무, 서어나무, 까치박달, 밤나무 등과 자생한다고 밝혔으며(Lee et al., 1994), 비목나무를 비롯하여 대부분 계곡부에 주로 자생하는 친수성 식물로 식생단위 8에서 우점하여 나타나는 비목나무, 산뽕나무, 밤나무, 층층나무, 졸참나무, 쪽동백나무 등 또한 대부분 친수성 식물이 함께 나타나는 것을 알 수 있었다.
본 식생단위는 종군 1의 신갈나무군락군 이하의 종군에서 식별종들이 출현하지 않음으로 인하여 종군 1의 신갈나무, 철쭉 등의 식별종 출현에 의하여 신갈나무전형군락으로 구분되었다. 본 식생단위에 출현하는 우점도가 3이상인 식물종은 신갈나무, 철쭉 등 총 50분류군이 출현하였다.
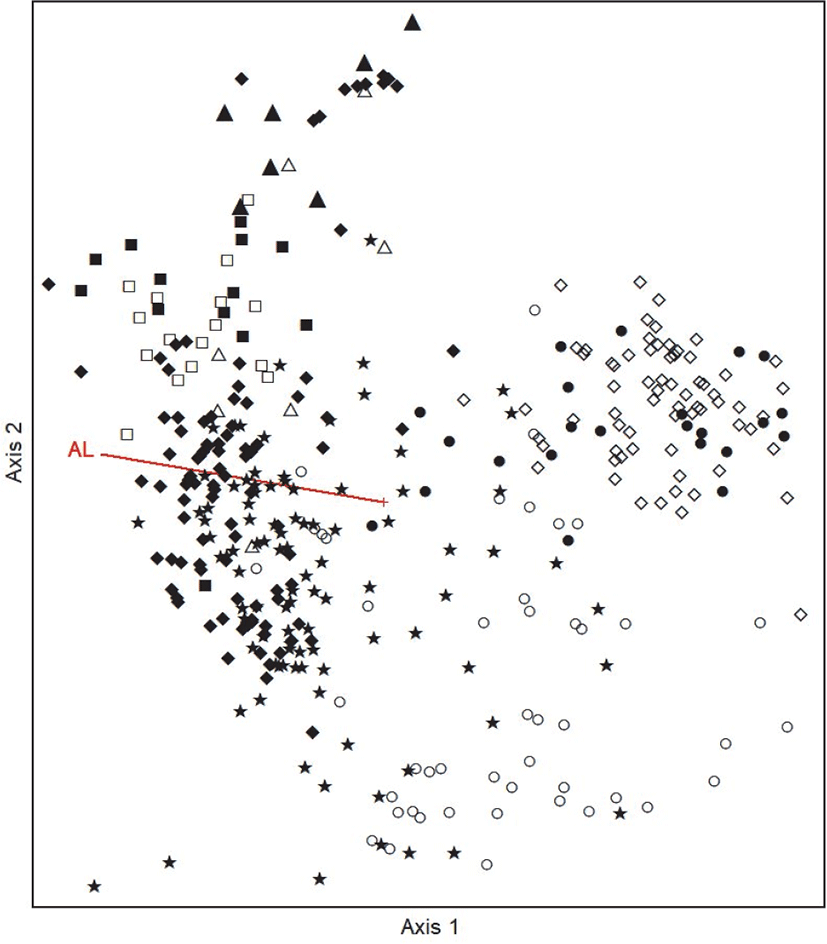
TWINSPAN에 의한 군락부류를 보완하고 군락의 분포를 직관적으로 알아보기 위해 Ordination 분석방법 중 DCA 기법을 적용(Choi and Kang, 2006)하여 1축과 2축을 기준으로 전체 373개소 조사구를 배치한 후 군락의 분포특성을 분석하였다(Figure 2). Ordination은 각 조사구간의 상이성을 바탕으로 조사구를 배치하여 요약하는 것으로(Orloci, 1978), 표본이 좌표의 위치에 근거하여 내재되어 있는 인자를 찾는 기법이며 이를 통해 각 조사구간의 분포거리에 따라 상이성 및 유사성을 확인할 수 있다(Ludwig and Reynolds, 1988; Kang et al., 2019). DCA 분석 결과 제1축과 2축의 eigenvalue가 각각 0.750, 0.645로 3개 축 전체의 합 1.917의 72.8%에 해당하여 total variance에 대한 집중률이 높은 편으로 나타났다. 조사구별 종구성에 의한 서열분석 결과, 식생단위 1∼5(구상나무군락)이 제2축을 기준으로 밀집하여 나타났고 식생단위 6∼7(소나무군락)이 양축을 기준으로 밀집하여 유형화 되는 것으로 나타났다. 식생단위 8(비목나무군락)은 제1축을 기준으로 밀집하여 나타나 위의 식생유형분류의 결과와 유사성을 확인할 수 있었다. 그 외 식생단위 간 구분이 명확히 구분되어 나타나지 않은 이유는 총 9개의 식생단위가 모두 신갈나무군락군의 최상위 계열에서 식생단위 및 종군이 구분되어져 전반적으로 서로 비슷한 종조성 및 입지환경요인을 가지기 때문인 것으로 판단된다.
식물사회학적 식생유형분류를 통해 구분된 9개의 식생단위를 기준으로 Curtis and McIntosh(1951)의 방법으로 식생단위별 중요치를 산출한 결과는 다음과 같다(Table 2).
평균상대우점치는 가문비나무(32.0)으로 가장 높게 나타났으며 사스래나무(16.1), 구상나무(12.4), 잣나무(11.0)의 순으로 나타났다. 교목층 중요치는 가문비나무(49.6)가 가장 높게 나타났으며 사스래나무(11.5), 구상나무(19.7), 잣나무(14.7)의 순으로 나타났다. 아교목층 중요치는 사스래나무(27.9)가 가장 높게 나타났고 가문비나무(19.9), 시닥나무(16.8), 마가목(13.3)의 순으로 나타났다. 관목층 중요치는 미역줄나무(13.1)가 가장 높게 나타났으며 진달래(12.1), 철쭉(10.1), 사스래나무(7.2)의 순으로 나타났다. 초본층 중요치는 실새풀(22.6)이 가장 높게 나타났고 산앵도나무(5.1), 퍼진고사리(4.8), 미역줄나무(4.2)의 순으로 각각 나타났다. 교목층에서 가문비나무가 크게 우점하고 있으며 당분간 가문비나무, 구상나무, 잣나무 등의 상록침엽수의 우세가 지속될 것으로 판단된다.
평균상대우점치는 가문비나무(23.6)으로 가장 높게 나타났으며 구상나무(19.5), 사스래나무(14.1), 잣나무(7.9) 의 순으로 나타났다. 교목층 중요치는 가문비나무(44.8)가 가장 높게 나타났으며 구상나무(31.6), 잣나무(14.4), 사스래나무(5.4)의 순으로 나타났다. 아교목층 중요치는 사스래나무(32.0)가 가장 높게 나타났고 시닥나무(14.4), 마가목(12.9), 구상나무(9.8)의 순으로 나타났다. 관목층 중요치는 철쭉(24.8)이 가장 높게 나타났으며 미역줄나무(13.8), 시닥나무(10.3), 진달래(9.0)의 순으로 나타났다. 초본층 중요치는 실새풀(19.4)이 가장 높게 나타났고 퍼진고사리(6.5), 가는잎그늘사초(4.5), 미역줄나무(4.2)의 순으로 각각 나타났다. 교목층에서 가문비나무, 구상나무, 잣나무 등 상록침엽수가 우세하지만 아교목층에서 사스래나무의 중요치가 높게 나타나 추후 천이과정을 고려하였을 때 지위경쟁을 보일 것으로 예측된다.
평균상대우점치는 호랑버들(31.1)으로 가장 높게 나타났으며 미역줄나무(16.4), 철쭉(11.8), 진달래(5.4)의 순으로 나타났다. 식생단위 3의 경우, 교목층과 아교목층에서 종조성이 나타나지 않았다. 관목층 중요치는 호랑버들(33.3)이 가장 높게 나타났으며 미역줄나무(17.3), 철쭉(12.6), 진달래(5.7)의 순으로 나타났다. 초본층 중요치는 새(15.2)이 가장 높게 나타났고 포아풀(8.0), 백운산원추리(7.8), 실새풀(6.4)의 순으로 각각 나타났다. 바람이 강하게 불어오는 노고단과 제석봉 인근 교목층까지 자리잡지 못한 왜림 및 초원지대인 식생단위 3은 소백산 비로봉 인근 초원지대처럼 시간 경과에 따른 초본층의 세력변화에 관한 모니터링이 필요할 것으로 판단되었다.
평균상대우점치는 구상나무(20.1)으로 가장 높게 나타났으며 잣나무(17.0), 신갈나무(15.1), 당단풍나무(7.0)의 순으로 나타났다. 교목층 중요치는 구상나무(31.4)가 가장 높게 나타났으며 잣나무(29.3), 신갈나무(19.8), 사스래나무(3.9)의 순으로 나타났다. 아교목층 중요치는 당단풍나무(19.6)가 가장 높게 나타났고 신갈나무(16.3), 구상나무(10.3), 쇠물푸레나무(9.4)의 순으로 나타났다. 관목층 중요치는 철쭉(15.0)이 가장 높게 나타났으며 미역줄나무(11.9), 진달래(11.9), 시닥나무(7.9)의 순으로 나타났다. 초본층 중요치는 실새풀(14.8)이 가장 높게 나타났고 새(5.3), 가는잎그늘사초(4.8), 퍼진고사리(3.3)의 순으로 각각 나타났다. 교목층에서 구상나무와 잣나무의 지위경쟁이 이루어지고 있는 것으로 보이지만 아교목층에서 상관을 이룰 수 있는 종들 가운데 신갈나무의 우점도가 가장 높아 추후 신갈나무림으로의 천이가 진행될 것으로 판단된다.
평균상대우점치는 신갈나무(27.2)으로 가장 높게 나타났으며 구상나무(15.0), 당단풍나무(10.3), 사스래나무(6.2)의 순으로 나타났다. 교목층 중요치는 신갈나무(47.3)가 가장 높게 나타났으며 구상나무(21.1), 사스래나무(8.0), 잣나무(7.7)의 순으로 나타났다. 아교목층 중요치는 당단풍나무(28.9)가 가장 높게 나타났고 구상나무(11.2), 신갈나무(10.5), 사스래나무(6.1)의 순으로 나타났다. 관목층 중요치는 철쭉(27.0)이 가장 높게 나타났으며 노린재나무(9.0), 미역줄나무(8.9), 진달래(5.8)의 순으로 나타났다. 초본층 중요치는 실새풀(14.1)이 가장 높게 나타났고 조릿대(7.8), 단풍취(4.8), 가는잎그늘사초(4.8)의 순으로 각각 나타났다. 교목층에서 신갈나무의 우점도가 구상나무에 비하여 높게 나타나고 있지만 아교목층에서는 두 수종의 중요치 차이가 거의 나타나지 않고 있으며 관목층에서는 구상나무의 중요치가 신갈나무보다 상대적으로 다소 높게 나타나고 있어 시간이 경과함에 따라 두 수종간 지위경쟁이 이뤄질 것으로 판단된다.
평균상대우점치는 소나무(35.5)로 가장 높게 나타났으며 밤나무(10.1), 신갈나무(6.8), 비목나무(5.1)의 순으로 나타났다. 교목층 중요치는 소나무(69.1)가 가장 높게 나타났으며 밤나무(7.5), 신갈나무(4.3), 산벚나무(3.3)의 순으로 나타났다. 아교목층 중요치는 밤나무(17.4)가 가장 높게 나타났고 신갈나무(12.1), 비목나무(10.9), 층층나무(10.1)의 순으로 나타났다. 관목층 중요치는 진달래(15.4)가 가장 높게 나타났으며 산초나무(11.5), 비목나무(9.0), 생강나무(8.2)의 순으로 나타났다. 초본층 중요치는 주름조개풀(21.3)이 가장 높게 나타났고 가는잎그늘사초(9.1), 진달래(3.6), 고마리(3.1)의 순으로 각각 나타났다. 교목층에서 소나무의 중요치가 현저히 높게 나타나고 아교목층에서 밤나무, 신갈나무, 비목나무, 층층나무, 서어나무 등이 소나무에 비하여 다소 높게 나타나 향후 천이과정상 참나무류가 우세할 것으로 보여지지만 아교목층내에서도 다수의 종의 지위경쟁이 불가피하여 당분간은 소나무의 우세가 지속될 것으로 판단된다.
평균상대우점치는 소나무(37.5)로 가장 높게 나타났으며 신갈나무(13.4), 굴참나무(8.0), 쇠물푸레나무(3.6)의 순으로 나타났다. 교목층 중요치는 소나무(67.0)가 가장 높게 나타났으며 신갈나무(13.3), 굴참나무(7.4), 리기다소나무(5.2)의 순으로 나타났다. 아교목층 중요치는 신갈나무(18.1)가 가장 높게 나타났고 소나무(13.3), 굴참나무(12.0), 쇠물푸레나무(9.2)의 순으로 나타났다. 관목층 중요치는 진달래(16.5)가 가장 높게 나타났으며 철쭉(8.4), 개옻나무(7.6), 싸리(7.4)의 순으로 나타났다. 초본층 중요치는 주름조개풀(18.2)이 가장 높게 나타났고 그늘사초(4.1), 담쟁이덩굴(3.6), 싸리(3.1)의 순으로 각각 나타났다. 교목층에서 소나무가 크게 우점하고 있고 아교목층에서도 신갈나무, 소나무, 굴참나무의 지위경쟁이 예상되는 바 당분간 소나무의 우세가 지속될 것으로 판단된다.
평균상대우점치는 층층나무(13.4)로 가장 높게 나타났으며 신갈나무(9.2), 일본잎갈나무(7.0), 졸참나무(5.7)의 순으로 나타났다. 교목층 중요치는 층층나무(21.5)가 가장 높게 나타났으며 일본잎갈나무(13.9), 신갈나무(10.1), 굴참나무(8.9)의 순으로 나타났다. 아교목층 중요치는 신갈나무(11.9)가 가장 높게 나타났고 산뽕나무(9.6), 당단풍나무(9.2), 층층나무(7.9)의 순으로 나타났다. 관목층 중요치는 비목나무(9.0)가 가장 높게 나타났으며 생강나무(7.7), 조릿대(6.2), 쪽동백나무(3.6)의 순으로 나타났다. 초본층 중요치는 조릿대(18.7)이 가장 높게 나타났고 주름조개풀(4.6), 뱀고사리(4.4), 오미자(3.6)의 순으로 각각 나타났다. 교목층에서 층층나무가 가장 높은 우점도를 보이지만 아교목층에서는 신갈나무가 가장 높게 나타나 추후 지위경쟁이 예상된다.
평균상대우점치는 신갈나무(31.3)로 가장 높게 나타났으며 당단풍나무(7.2), 물푸레나무(5.4), 굴참나무(4.4)의 순으로 나타났다. 교목층 중요치는 신갈나무(48.2)가 가장 높게 나타났으며 굴참나무(8.5), 물푸레나무(5.0), 노각나무(3.5)의 순으로 나타났다. 아교목층 중요치는 신갈나무(21.9)가 가장 높게 나타났고 당단풍나무(19.7), 쇠물푸레나무(9.5), 까치박달(7.8)의 순으로 나타났다. 관목층 중요치는 철쭉(18.4)이 가장 높게 나타났으며 미역줄나무(10.4), 노린재나무(10.3), 조록싸리(10.3)의 순으로 나타났다. 초본층 중요치는 조릿대(19.5)이 가장 높게 나타났고 실새풀(7.8), 가는잎그늘사초(4.6), 단풍취(3.3)의 순으로 각각 나타났다. 교목층과 아교목층에서 신갈나무의 중요치가 크게 높아 신갈나무림으로의 천이과정이 지속될 것으로 판단된다.
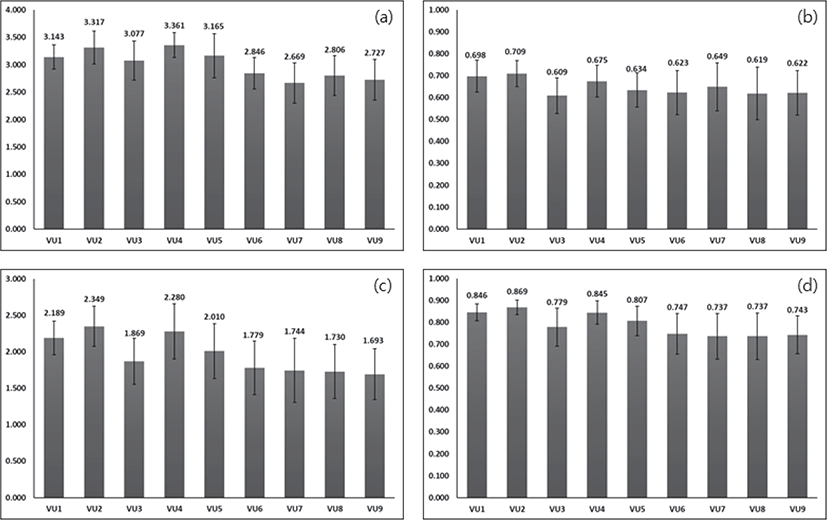
백두대간 지리산권역(육십령-천왕봉 구간) 마루금의 산림식생을 식물사회학적 방법으로 분석하여 구분된 9개의 식생단위를 기준으로 종다양도를 분석한 결과는 다음과 같다(Figure 3). 평균해발고도가 상대적으로 높은 구상나무군락(식생단위 1∼5)의 종다양도가 다른 식생단위에 비하여 종다양도가 높게 나타났다. 해발고도 증가에 따른 종다양성의 변화는 선형, 단봉형, 시그모이드(sigmoid)형과 같이 다양한 양상과 지역성이 보고되고 있다(Wang et al., 2007). 저지대에 비해 고지대 능선부는 바람이 많고 숲 틈이 상대적으로 많아 바닥 광량이 충분하며, 습윤한랭한 조건에 의해 토양 염류가 풍부하며, 이것은 식물종 사이의 자원 경쟁 강도는 낮추어 풍부도는 증가시킬 수 있다. 개체목 또는 개체군이 수관을 이루는 동안 교란에 의해 고사되거나 쓰러져 수관층이 열리고 광온성식물(eurythermal plant)에 의해 다시 채워지는데(Brokaw and Busing, 2000), 지리산권역 내 분포하는 구상나무군락과 가문비나무군락은 그 생육지가 아고산대에 한정되어 형질교환이 가능한 개체군의 크기가 매우 작기 때문에 진화가 어렵고 그 결과 멸종될 가능성이 매우 높은 식물 중 하나이다(Soule and Simberloff, 1986; Simberloff 1988). 또한 기후변화 및 여러 원인에 의해 고사율이 높아(Kim, 1994; Koo et al., 2001; Kim et al., 2016), 많은 숲 틈이 발생한 것을 관찰할 수 있었다. 또한 종다양성은 수관층유형, 경관 그리고 교란의 종류에 의해 결정되는데(Ellum et al., 2010; Duguid et al., 2013), 기존 우점종에 대한 다양성보다 교란 발생 후 이입되는 종에 의한 종다양성 증가가 나타나(Kumar and Ram, 2005; Bolton and D’Amato, 2011), 많은 교란이 발생하고 있는 고해발지역 상록침엽수림이 높은 종다양도를 갖는 것으로 판단되었다.
환경인자(해발고도, 사면경사도, 지형, 암석노출도, 토양노출도)와 식생단위와의 상관관계를 살펴보면(Figure 4), 각 축의 설명값(eigen value)은 1축(64.1%), 2축(19.1%)으로 백두대간 마루금 위주의 조사였기 때문에 식생단위 1, 2, 4, 5와 해발고도가 제1축상 배열과 양의 상관관계를 가지는 것으로 나타났다.